
异源表达TaEDR1抑制拟南芥edr1白粉病抗性
2018-03-15 11:09武广珩傅仙玉邓家耀刘金仙
武夷学院学报 2018年12期
武广珩,傅仙玉,邓家耀,刘金仙*
(1.福建省生态产业绿色技术重点实验室,福建 武夷山 354300;2.武夷学院 生态与资源工程学院,福建 武夷山 354300;3.武夷学院 茶与食品学院,福建 武夷山 354300)
白粉病是由白粉菌科(Erysiphacesae)真菌引起的植物病害。当植物被侵染时,会产生大量由菌丝体、分生孢子梗和分生孢子构成的肉眼可见的白色粉状物,并由此而得名。白粉病在全世界分布广泛,危害双子叶植物,如葡萄、草莓、黄瓜等。同时它也侵染诸多的禾本科植物,如小麦、大麦、燕麦和多种牧草。其中小麦白粉病是由气传性专化寄生真菌布氏白粉菌(Blumeria graminis)引起的真菌性病害,为世界各主要麦区的主要病害之一,且发病范围日益扩大,危害程度不断加重,可引起13.4%~34%的产量损失[1]。白粉菌是高等植物的专性活体寄生菌,通常生长在植物的表面,菌丝体表生或半内生,以吸器进入寄主细胞吸取养分,从菌丝上长出直立的分生孢子梗,在梗端形成单生或成串的分生孢子。
拟南芥白粉病抗性相关基因EDR1基因编码一个Raf样MAPKKK蛋白激酶负调控水杨酸(Salicylic Acid,SA)诱导的防卫反应,edr1突变体表现出对白粉菌的抗性,以及病原菌诱导后比野生型积累更多的PR1转录本[2-3]。EDR1通过与MAPK级联蛋白激酶MKK4和MKK5相互作用以及影响二者的蛋白表达水平来精确控制植物的免疫防卫反应[4]。KEG基因编码一个包含环指结构E3泛素连接酶结构域和激酶域的蛋白,该基因的一个特异错义突变能够抑制edr1的相关表型[5]。KEG与EDR1相互作用并招募EDR1在反式高尔基体和早胞内体(TGN/EE)上定位[6]。KEG调控内膜系统蛋白的运输,同时在白粉菌侵入的表皮细胞中发生特异性的降解[7]。EDR1还与另一个正调控植物免疫和细胞死亡的E3泛素连接酶ATL1在TGN/EE中相互作用;当在拟南芥和烟草中过量表达ATL1导致植物出现细胞死亡的表型,并且这种细胞死亡能够被EDR1抑制;而与此对应的是,下调ATL1的表达能够有效的抑制edr1介导的白粉病的抗病表型,以上这些暗示ATL1可能是EDR1潜在的底物[8]。近期研究发现,edr1的抑制子基因LLG1,编码一个糖基磷脂酰肌醇锚定蛋白,与flg22识别受体FLS2互作,调控FLS2蛋白的积累及flg22诱导的FLS2降解,影响BIK1的磷酸化和活性氧的产生等,从而调控FLS2激活的免疫反应[9]。
EDR1基因在在水稻和小麦的抗病中也表现出重要的作用。通过CRISPR/Cas系统的基因定点编辑技术在普通小麦中创制的Taedr1基因编辑突变体同样表现出对白粉菌的抗性,且无明显的发育缺陷,验证了EDR1基因在小麦育种的应用前景[10]。水稻OsEDR1的T-DNA插入突变体和RNAi转基因植株对水稻白叶枯病菌黄单胞杆菌表现出抗性,在黄单胞杆菌和水稻的互作中,OsEDR1的表达促进了乙烯的合成但抑制了水杨酸(SA)和茉莉酸(JA)介导的抗性信号通路[11]。郭传宇等研究还发现AtEDR1也参与拟南芥对稻瘟菌的非寄主抗性[12]。
为了进一步研究EDR1蛋白的抗白粉菌机制,明确EDR1蛋白是否具有跨物种的功能保守性,本研究选择研究较为清楚的小麦EDR1蛋白作为对象,构建了TaEDR1基因的过量表达载体并转化了拟南芥edr1突变体,对转基因拟南芥的抗病表型进行了相关分析,为解析EDR1蛋白功能,以及为EDR1同源蛋白在其他经济作物中的利用提供理论依据。
1 材料与方法
1.1 材料
野生型拟南芥(Arabidopsis thaliana)Columbia-0生态型 (Col-0)、pad4-1和edr1突变体,大肠杆菌DH5α,农杆菌GV3101由本实验室保存。植物表达载体pBI-1.4t,白粉菌菌株Golovinomyces cichoracearum UCSC1和小麦品种KN199由福建农林大学唐定中教授馈赠。
1.2 方法
1.2.1 拟南芥的种植
拟南芥种子经表面消毒和4℃春化处理三天,然后平铺于在含有1%蔗糖的1/2 MS固体培养基上,种子萌发7 d后移植。对于观察表型的植物和用于瞬时表达的烟草,需放置在9h光照、光照强度7 000~8 000 lx、相对湿度50%~60%和温度21~24℃的温室;对于收集种子的植物,我们将其放置在16 h光照、光照强度10 000~12 000 lx、相对湿度50%~60%和温度21~24℃的温室。
1.2.2 载体构建
为了获得35S-TaEDR1-pBI1.4t载体,我们使用Promega RNA提取与反转录试剂盒获得小麦KN199幼苗(出芽5 d)的cDNA。以cDNA为模板,设计特异引物 OE-F(CGCTCGAGATGAAGATCCCGTTTGTGACCAAGT,划线部分为Xho I酶切位点)和OE-R(CCGTCGACATGTGCGGTTCACCGATATAAAGAT, 划线部分为Sal I酶切位点)使用KOD plus(东洋纺)将TaEDR1完整CDS序列扩增出来。扩增程序:96℃预变性 5 min,95℃变性 10 s,68℃延伸3 min,68℃补充延伸5 min,30个循环。利用琼脂糖凝胶DNA回收试剂盒 (中科瑞泰)回收目的片段,插入克隆载体pEASY-Blunt Simple中(北京全式金生物公司),将测序验证后的序列通过Xho I和Sal I(大连宝生物公司)酶切后连入由CaMV35S驱动的植物表达载体pBI1.4t中,进行菌落PCR及双酶切检测,将验证正确的重组质粒转化农杆菌GV3101感受态细胞,进行菌落PCR鉴定阳性克隆。PCR引物合成及产物测序由上海生工生物公司完成。
1.2.3 遗传转化与纯合体筛选
将上述含重组质粒的农杆菌活化后,继代培养至OD600=2.0,常温3 200 g离心收集菌体,用等体积含0.03%SilwetL-77的5%蔗糖溶液重悬,使用2 mL吸管将重悬的菌液滴在拟南芥突变体edr1花上。暗培养24 h后放置于生长室,在长光照条件培养至种子成熟,得到T0代种子。将T0代种子在相应抗性(卡那霉素,50 μg/mL)的不加蔗糖的MS平板上筛选,5~8 d后挑选转化子,移苗到土中继续培养。T1代转化子用pBI1.4t载体筛选标记基因新霉素磷酸转移酶Ⅱ(Neomycin phosphotransferaseⅡ,NPTⅡ) 基因片段进行扩增,设计引物 NPTⅡ-F:5′-GGCTATTCGGCTATGACTGGGC-3′和 NPT Ⅱ -R:5′-GGCGATACCGTAAAGCACGAGG-3′PCR 验证,长度 660 bp。 筛选得到的转基因阳性植株,进一步培养收获转基因种子。提取筛选到的阳性植株T3代的叶片RNA,反转录生成 cDNA,利用实时定量 PCR仪 (Eppendorf Realplex2)进行扩增(SYBR Premix Ex Taq Kit,反应程序为:95℃预变性90 s,95℃变性15 s,55℃退火15 s,72℃延伸20 s,40个循环)。以拟南芥ACT2基因为内参,分别设计TaEDR1基因定量引物Q-TaEDR1-F:5′-GCTCATTGGCATTCAAGGGA-3′和 Q-TaEDR1-R:5′-GCATTGTCAGTGGCATTTCT-3′,内参 ACT2 的引物 Q-ACT2-F:5′-AGTGTCTGGATCGGTGGTTC-3′和Q-ACT2-R:5′-CCCCAGCTTTTTAAGCCTTT-3′。 病程相关基因 PR1 引物:Q-PR1-F:5′-GTGGGTTAGCGAGAAGGCTA-3′和 Q-PR1-R:5′-ACTTTGGCACATCCGAGTCT-3′。
1.2.4 表型分析
Typan Blue染色:由于Typan Blue可以染色白粉菌的菌丝和分生孢子以及死亡的植物细胞。4周大小的拟南芥植物叶片接种保存于易感性突变体pad4-1上的白粉菌菌株Golovinomyces cichoracearum UCSC1;接种6 d后,采收叶片,将其置于Trypan Blue染液 (20 mL乙醇,10 mL苯酚,10 mL超纯水,10 mL 83%乳酸和10 mg台盼蓝粉末)中煮沸2~5 min;染色后的叶片用水合三氯乙醛(2.5 g/ml)过夜脱色;用超纯水洗涤三次后用50%甘油保存。依据Wang等[13]采用的方法,显微镜下计数每个独立孢子形成的分生孢子梗的数目,随后统计分析。对于基因型个体的表型记录,则取接种8 d后的叶片进行Typan Blue染色,使用光学显微镜(Leica DM2500)进行观察和拍照。
DAB染色:DAB可以染色白粉菌的侵染植物细胞时产生的过氧化氢(H2O2)。按一下步骤进行:收集接种白粉菌48 h的叶片,将其置于DAB(1 mg/mL,pH=5.8)染液中染色12 h,随后用无水乙醇脱色;用超纯水洗涤三次后用50%甘油保存;使用光学显微镜(Leica DM2500)进行观察和拍照。
1.2.5 数据分析
通过GenBank数据库进行序列检索,利用Clustal omega软件进行序列比对,用MEGA 7软件构建系统发育树。使用Excel 2007对分生孢子梗计数数据进行分析处理。
2 结果与分析
2.1 EDR1同源蛋白保守性
为了分析EDR1蛋白在植物中的保守性,我们对NCBI数据库中EDR1近缘蛋白进行提取,利用Clustal Omega 软件进行比对分析(https://www.ebi.ac.uk/Tools/msa/clustalo/)并通过MEGA7.0软件构建蛋白序列进化树。发现EDR1同源蛋白存在于单子叶和双子叶植物中,并呈现较高的保守性(图1A)。其中,不同物种中EDR1蛋白的激酶结构域的一致性高达89.3%(图1B)。

图1 EDR1同源蛋白序列分析Figure 1 Sequence analysis of EDR1 homologous proteins
2.2 TaEDR1基因表达载体构建
为了在拟南芥中异源表达小麦的TaEDR1基因,播种小麦KN199种子,生长5 d后,收集新鲜叶片提取RNA,经反转录获得cDNA。利用特异性引物OE-F和OE-R扩增长度获得了2896bp的TaEDR1 CDS编码区 (图2A)。将 CDS序列连接到 pEASY-Blunt Simple克隆载体中,测序正确后用Xho I和Sal I双酶切连接到pBI1.4t表达载体。将构建好的重组表达载体35S-TaEDR1-pBI1.4t转化大肠杆菌并进行菌落PCR验证,提取验证正确的菌落质粒用Xho I和Sal I双酶切鉴定,能够切出目的片段,表明载体构建成功(图2B)。

图2 TaEDR1 CDS表达载体的构建Figure 2 TaEDR1 CDS expression vector constructed
2.3 异源表达TaEDR1抑制突变体edr1表型
通过农杆菌GV3101介导的浸花法将重组植物表达载体35S-TaEDR1-pBI1.4t转入拟南芥突变体edr1,获得16个具有T1代转化子,PCR扩增转化子中的NPTII基因(660 bp),发现12个阳性转基因植株(图3A)。此后,4周大小的12个T1代的转基因植株、野生型和edr1接种白粉菌,8 d后发现T1代的转基因植株、野生型各自的叶片上都有大量白粉菌菌丝和分生孢子梗的生长,而edr1突变体仅有少量的菌丝和分生孢子梗,并出现明显的细胞死亡。说明拟南芥中异源表达TaEDR1基因能够抑制突变体edr1的抗病表型(图3B)。
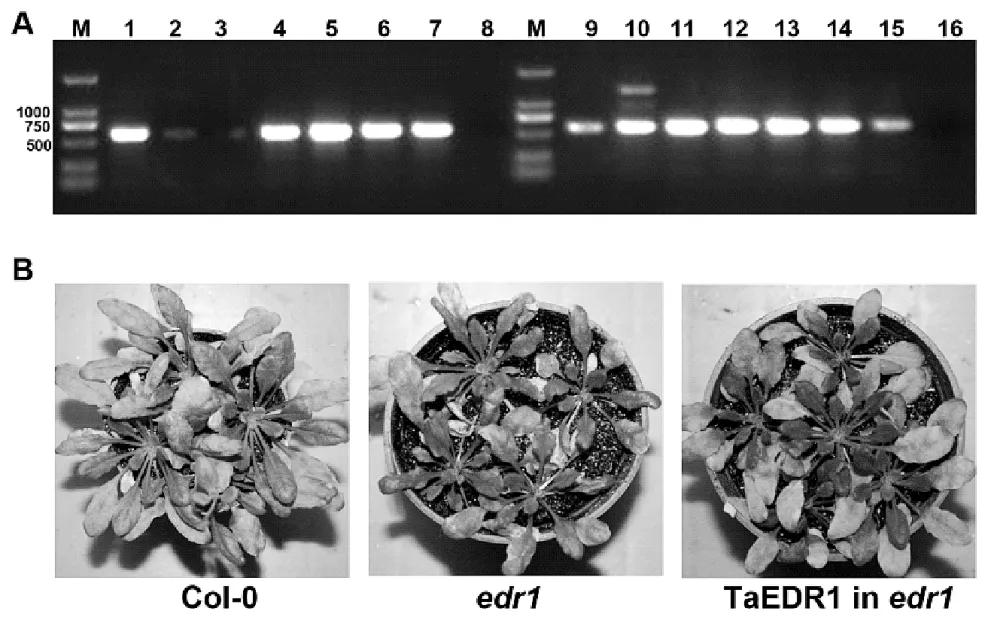
图3 异源表达TaEDR1抑制拟南芥edr1突变体白粉菌抗病表型Figure 3 Heterologous expression of TaEDR1 complementary edr1 mutant resistance phenotype
2.4 异源表达TaEDR1抑制突变体edr1中PR基因的表达和过氧化氢的积累
为了进一步了解TaEDR1在拟南芥抗病反应中的功能,我们将35S-TaEDR1-pBI1.4t遗传转化植株收种和分离鉴定,获得T3代的稳定转基因株系。提取这些株系的RNA并合成cDNA,利用实时荧光定量PCR检测TaEDR1在转基因株系中的表达水平,选取表达量最高的OE-5进行后续实验(图4A)。
为了明确TaEDR1在拟南芥中抑制edr1表型的能力,4周大小的野生型、edr1和转基因植株OE5接种白粉菌。通过白粉菌在叶片表面的生长观察和白粉菌单个孢子上分生孢子梗生长的定量计数发现OE5和野生型无明显差异,都明显多于edr1突变体上生长的分生孢子梗数,即表现出更加抗病的表型(图4B、4C和4E)。此外,对4周大小的野生型、edr1和OE5进行白粉菌接种,在48 h后取新鲜叶片进行DAB染色。结果发现突变体edr1在白粉菌接种48 h后,白粉菌孢子入侵点处出现大量过氧化氢积累,而Col-0和OE-5植物仅出现少量积累(图4D)。
为了了解突变体中病程相关基因表达是否发生变化,对4周大小的野生型Col-0、edr1和OE5接种白粉菌,在接种后0、2、4 d收集叶片提取RNA、转录和对抗病相关基因AtPR1进行实时定量PCR分析。结果发现,接菌前后Col-0和OE-5中AtPR1基因的表达都没有明显的差异;突变体edr1中AtPR1的表达接菌后2 d,较Col-0和OE-5已经出现明显的上调,在接菌后4 d,上调幅度已经远大于Col-0和OE-5(图4F)。
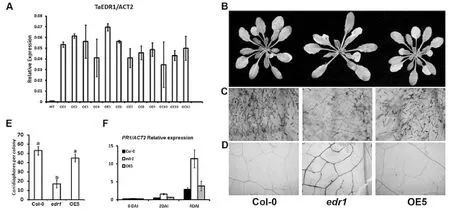
图4 TaEDR1抑制拟南芥edr1突变体中白粉菌诱导的AtPR1基因的大量表达和H2O2的积累Figure 4 TaEDR1 inhibits the expression of AtPR1 gene and accumulation of H2O2in edr1 mutant
A:在12个T3转基因植株中定量分析TaEDR1表达量,以AtACT2作为参照;B:4周大小的野生型、edr1和转基因植株OE5,接种白粉菌后8 d的表型;C:Trypan Blue染色(B)中的叶片;D:对4周大小的野生型、edr1和转基因植株OE5进行白粉菌接种,48 h后取新鲜叶片进行过DAB染色,褐色显示过氧化氢的积累;E:定量分析野生型、edr1和转基因植株OE5叶表面单克隆白粉菌形成分生孢子梗的数目。数据经t检验统计分析,不同字母代表差异显著。该实验重复3次,得到类似的结果(P<0.01);F:4 周大小的野生型、edr1 和转基因植株 OE5 接种白粉菌,实时定量PCR检测接种白粉菌后0、2、4 d的AtPR1的表达情况
3 讨论与结论
植物在生长发育的过程中可能会遭遇各种生物胁迫。以模式植物拟南芥为研究对象,人们通过鉴定和克隆对各种病原菌增强或减弱抗性的突变体,找到了很多基因参与了植物对病原微生物的防卫反应。EDR1基因作为一个抗病基因,多项研究发现其调控了植物(拟南芥、水稻和小麦)对真菌和细菌的抗性[2,10,11]。序列比对发现来自于不同单双子叶植物的EDR1蛋白表现出很高的保守性,其抗性功能都表现出大量的过氧化氢和胼胝质的积累,以及病程相关基因PR1表达的大幅度上调[4,10,12]。由此我们认为EDR1是一个具有广谱抗性的保守蛋白,在不同物种中发挥着类似的功能的基因。由此推测,不同来源的EDR1蛋白很可能能够在不同物种中发挥同样的功能。我们的结果证明了小麦的EDR1基因导入拟南芥中,小麦EDR1蛋白可以在拟南芥中发挥功能,并且激发与AtEDR1类似的信号通路。EDR1蛋白在单双子叶植物中大量存在、功能高度保守,为我们今后在其它经济作物中利用EDR1同源基因提高不同植物抗病性,乃至在分子设计育种中的利用,提供了坚实的理论依据。
此外,复杂农作物物种的植物病理学研究由于缺乏相应的研究体系,发展较为缓慢。利用模式植物拟南芥和相应的突变体,与不同病原菌(假单胞杆菌、卵菌和白粉菌等)的植物病理研究系统的建立,将有利于复杂经济作物的抗病研究。
猜你喜欢
今日农业(2021年19期)2022-01-12
疯狂英语·新策略(2021年9期)2021-11-02
农业科技通讯(2021年1期)2021-03-06
热带作物学报(2019年4期)2019-06-11
东坡赤壁诗词(2019年1期)2019-04-30
中国森林病虫(2018年4期)2018-09-19
新农业(2017年2期)2017-11-06
上海农业学报(2017年3期)2017-04-10
天津医科大学学报(2015年2期)2015-12-22
塔里木大学学报(2015年1期)2015-04-25
