
洪泽湖河蚬增殖放流效果评估
2018-03-12 05:52:28刘燕山张彤晴唐晟凯李大命刘小维王莲莲
水产养殖 2018年1期
刘燕山 ,张彤晴 ,唐晟凯 ,李大命 ,刘小维 ,王莲莲
(1.江苏省淡水水产研究所,江苏 南京 210017;2.江苏省内陆水域渔业资源重点实验室,江苏 南京 210017;3.中国科学院南京地理与湖泊研究所,江苏 南京 210008)
河蚬(Corbicula fluminea)栖息于淡水及咸淡水的江河、湖泊及入海口的沙或沙泥质底质中,在我国广有分布[1]。河蚬具有非常高的营养价值,其软体部分粗蛋白含量达63.3%,有氨基酸17种,富含钙、钠、钾、镁、铁、锌、铜、锰等矿物元素,是一种营养价值高的淡水经济贝类[2]。洪泽湖河蚬作为其农产品地理标志,是当地最具特色的水产品之一,其外壳干净、多呈棕黄色、纹沟粗疏、体厚饱满,鲜活河蚬气味清新,煮熟后鲜味浓厚,略带甜味,感观特征明显[3]。洪泽湖河蚬品质优良,深受国内外人们的喜爱,每年有近万吨的河蚬出口日本和韩国[4],据统计,2008年以来,日韩河蚬市场的80%以上来自洪泽湖[3]。然而,近年来洪泽湖自然水域河蚬的资源量呈现下降的趋势,在洪泽湖底栖动物的调查中发现,1991年河蚬的生物量约108.6 g/m2[5],2005年为120.8 g/m2[6],至 2010 年仅剩 19.5 g/m2[7]。
增殖放流是恢复衰退渔业资源的有效方法之一,同时它也有利于优化渔业资源结构、改善水域生态环境。因此,为减缓洪泽湖自然水域河蚬的资源量下降的趋势,本研究以剪草沟、大新滩和马浪岗的自然水域作为研究试点,进行河蚬的增殖放流实验,并在放流后的第二年对其形态学指标、规格和生物量等进行监测,以评估洪泽湖自然水域河蚬的增殖放流效果。
1 材料方法
1.1 样品采集和处理
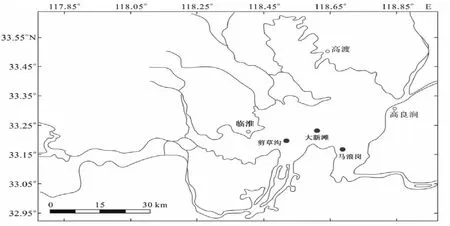
图1 洪泽湖河蚬增殖放流站位点
2014年11月在洪泽湖的剪草沟(33.21541°N,118.50311°E)、大新滩(33.25116°N,118.5982472°E)和马浪岗(33.17079°N,118.69689°E)进行河蚬的增殖放流(图1),三个站点没有或很少有河蚬,各点的水域面积、河蚬的放流量、放流生物量和放流规格如表1所示,并于2015年11月、2016年3月、7月和9月对三个站点的增殖放流效果进行评估。采集河蚬工具为划耙,耙宽0.7 m,耙间距6 mm,采样时向前行进20 m左右,每个点采集三个样方。将河蚬样品冲洗干净,带回实验室测量。采样时记录站点水体温度。
1.2 样品测量和数据分析
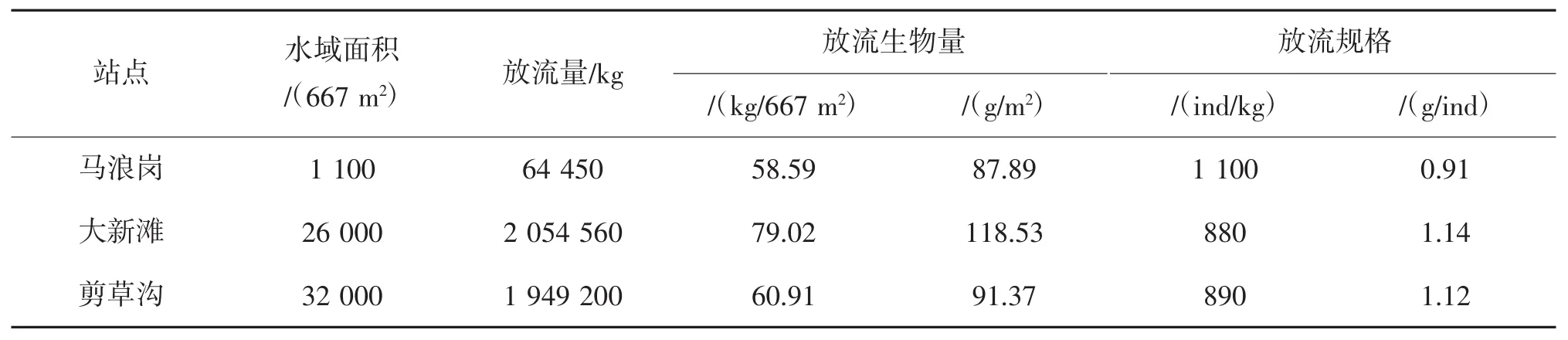
表1 河蚬的放流量和放流规格
将采集的河蚬样品在实验室进行称重,用于计算生物量,生物量(g/m2)=样方重(g)/(0.7×20)。从每个样方随机选取50个河蚬,用游标卡尺测量其壳长、壳宽和壳高(精确到0.10 mm),然后将河蚬用抹布擦干后,放在电子天平上秤个体体质量(湿重,精确到 0.10 g)。
利用Excel 2010和SPSS 19.0对数据进行统计分析并绘图,单因素方差分析形态学指标随时间或地点的差异性,用两因素方差分析时间和站位对河蚬规格和生物量的影响,数据用平均值±标准差来表示,以P<0.05为差异具有统计学意义水平。
2 结果
2.1 水温
监测期间,水体的温度随时间变化显著(P<0.01),如图2所示,3月份水温最低,为10.98℃,7月最高,为32.30℃。
2.2 形态学指标
放流后第2年,三个点的河蚬形态学指标监测结果如表2所示,可以看出,2016年3月各点的河蚬形态学指标均降低,其余监测时间均为马浪岗河蚬的壳长和壳高大于大新滩和剪草沟。将各形态学指标的所有数据按时间和地点进行单因素方差分析,结果表明,2015年11月河蚬的长、宽、高显著高于2016年 3月、7月和9月(P<0.05),而大新滩、剪草沟的河蚬形态学指标显著低于马浪岗(P<0.05)。
2.3 规格
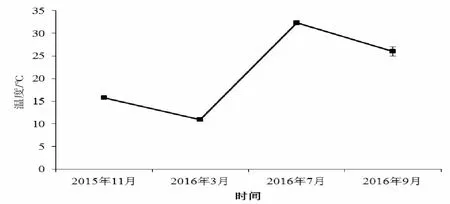
图2 监测期间水体温度变化
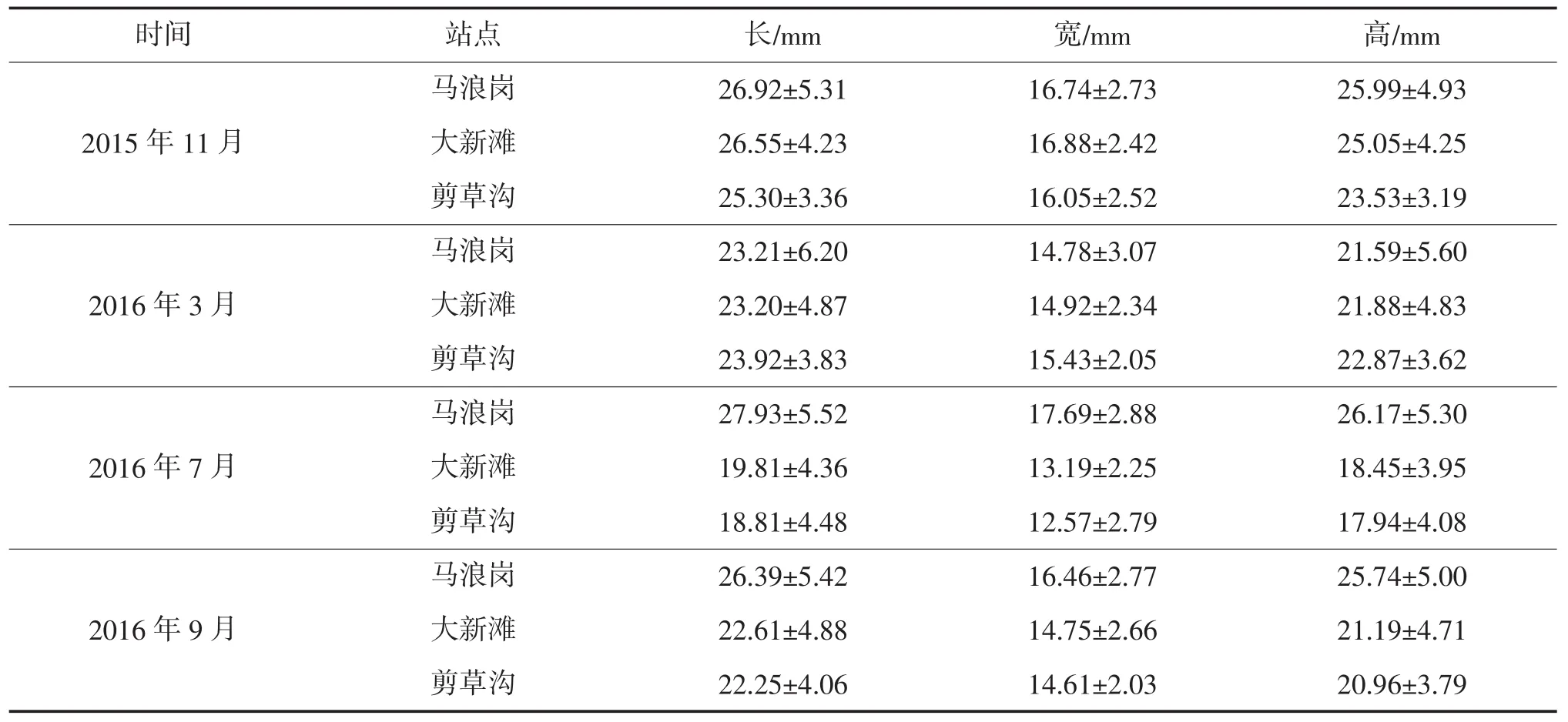
表2 放流后第二年河蚬的形态学指标
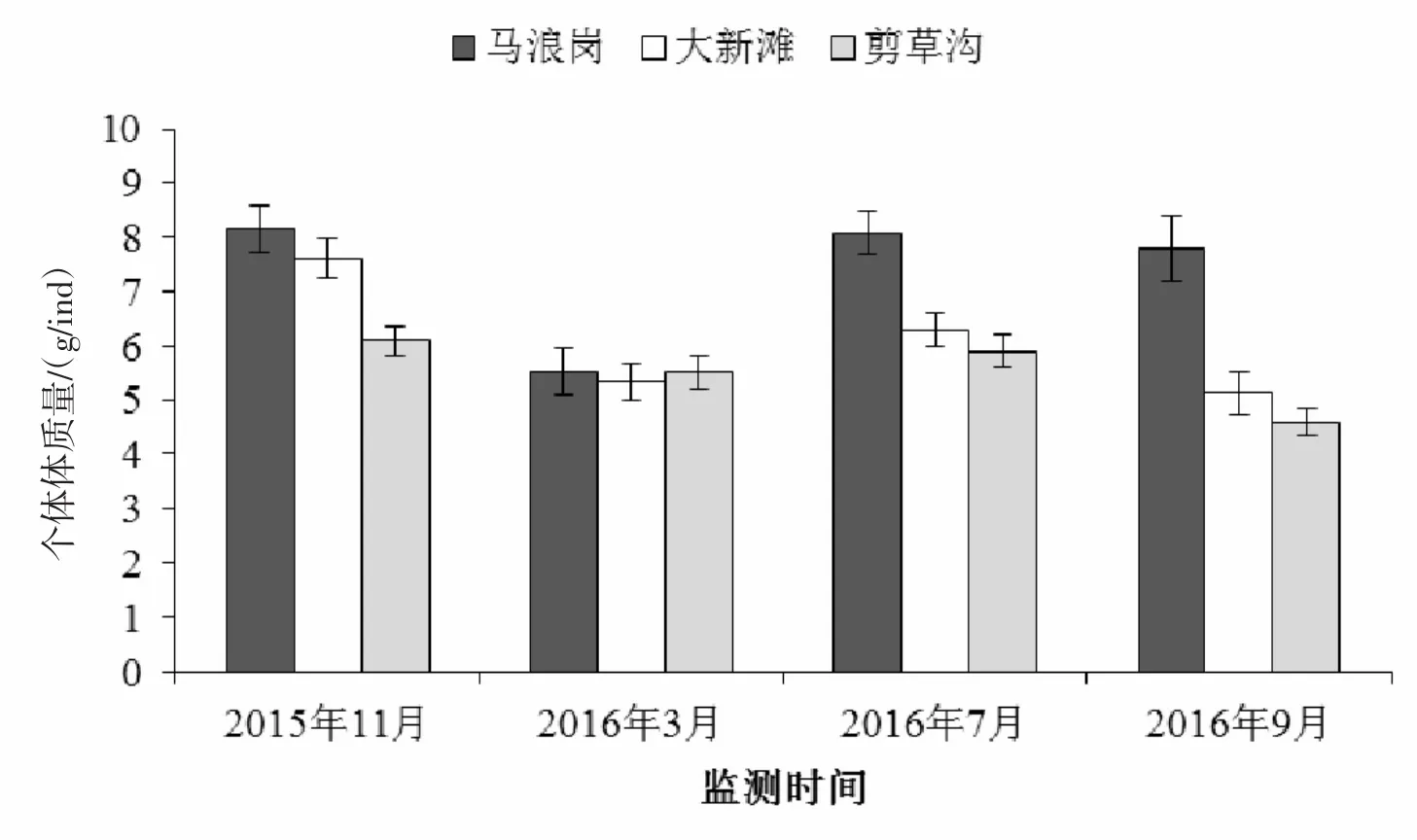
图3 放流后第二年河蚬的规格
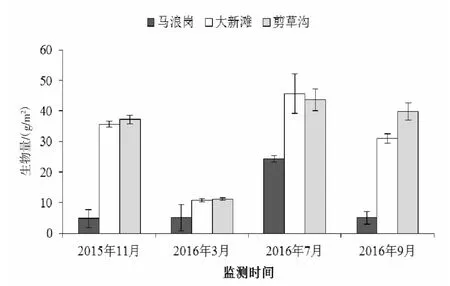
图4 放流后第二年河蚬的生物量
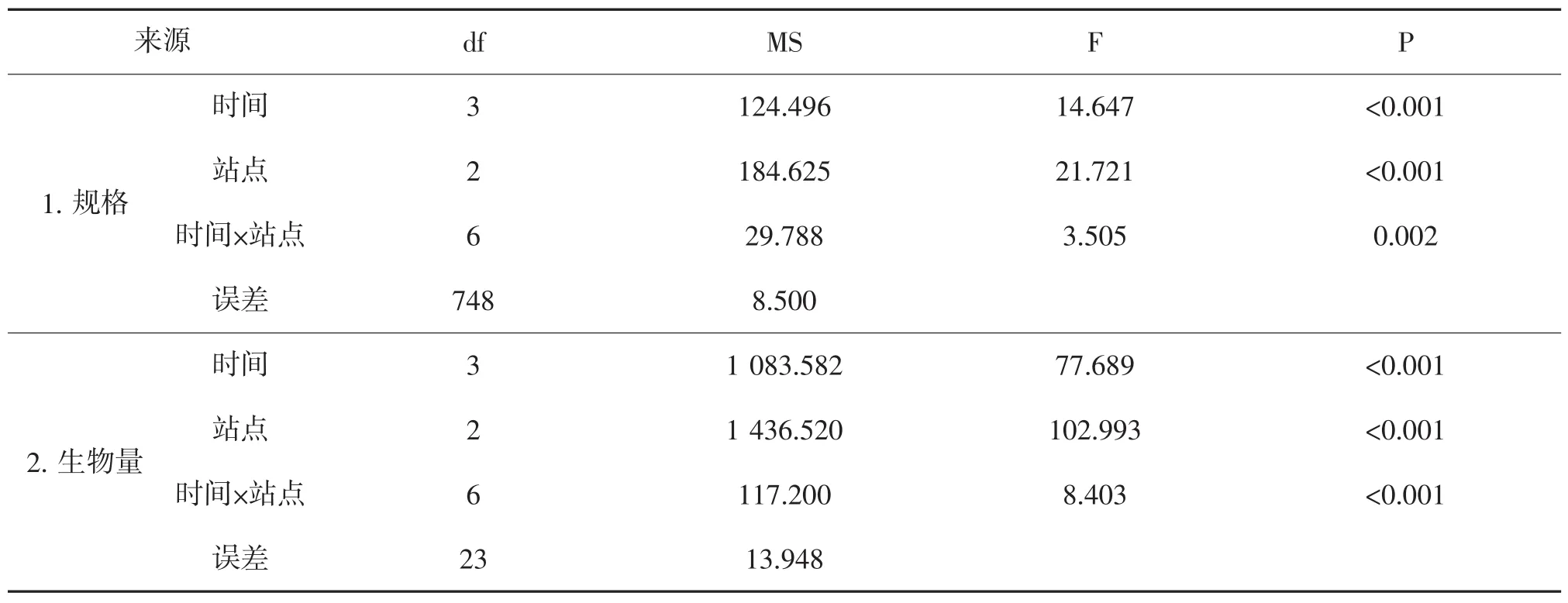
表3 河蚬规格和生物量时空变化的两因素方差分析
放流后第2年,河蚬的规格在站点间差异具有统计学意义(P<0.01),并随时间变化差异具有统计学意义(P<0.01,表3)。三个样点河蚬的规格如图3所示,马浪岗、大新滩和剪草沟的河蚬规格分别在5.54~8.14、5.13~7.62 和 4.58~6.10 g/ind 之间,各监测时间内规格均为马浪岗大于大新滩和剪草沟,各站点均于2016年3月减小后有所增长,最大值出现在2015年11月。至2016年7月,三个站点的河蚬规格分别达到8.09 g/ind、6.29 g/ind和5.9 g/ind。
2.4 生物量
放流后第2年,河蚬的单位面积生物量在站点间差异随时间变化差异具有统计学意义(P<0.01,表3),其生物量监测结果如图4所示,监测期间,马浪岗河蚬的生物量均显著小于大新滩和剪草沟(P<0.05);马浪岗河蚬的生物量呈先增长后降低的趋势,2016年7月值最大为24.29 g/m2,而大新滩和剪草沟河蚬的生物量则呈现先降低后增长的变化趋势,最低值均出现在2016年3月,分别为10.77 g/m2和11.15 g/m2,最大值均为2016年7月,分别为45.69 g/m2和43.69 g/m2。
3 讨论
增殖放流是缓解洪泽湖河蚬的资源量逐渐降低的最有效的途径之一,因此进行河蚬的增殖放流就显得尤为重要,目前在该湖区主要有自然水域苗种放流[8-10]和网围增殖两种增殖方法,相对而言,自然水域苗种放流法具有对自然环境影响小、操作简单,人工投入少,成本低等优点。本研究于2015年11月在马浪岗、大新滩和剪草沟三个自然水域站点分别进行河蚬苗种的增殖放流,至2016年7月三个站点的河蚬规格均大幅增长,分别达到8.09 g/ind、6.29 g/ind和5.9 g/ind,而单位面积生物量分别为 24.29 g/m2、45.69 g/m2和 43.69 g/m2。三个区域河蚬的规格和生物量差异较大,这是由于河蚬的增殖放流效果与其存活和生长密不可分。
影响河蚬存活和生长的因素很多,归纳起来,可以分为非生物因素、生物因素和人为因素,其中非生物因素如水温、溶氧、水流、底质等[11-13],生物因素如天敌[14-15]、饵料生物[16]、竞争者[17-18]等,以及人为因素如污水排放、过度捕捞、采砂、水工设施的大量建造等[19-20],而人为因素是其中的主要因素。本研究发现,三个站点河蚬的形态学指标和规格均为2015年11月最大,由于目前洪泽湖河蚬实行全年全湖禁捕,只有在特殊情况下经授权方可捕捞,因此这可能是受非法捕捞影响的结果,而在洪泽湖捕捞河蚬的常见工具为划耙,齿间距一般在8 mm左右,因此规格较大的河蚬被捕捞而小个体河蚬留存,是导致河蚬规格偏小的原因。研究还发现,河蚬的生物量均为马浪岗小于大新滩和剪草沟,而形态学指标和规格均为马浪岗大于大新滩和剪草沟,这一方面是由于在马浪岗附近有大量的采砂船,非法采砂情况时有发生,致使实验站点水体的浑浊度增大,水体透明度降低,藻类等饵料生物大幅度减少,进而带来的是以藻类等为食的生物种群密度降低,故该区域河蚬生物量较少;另一方面随着以藻类等为食的生物种群密度的降低,该区域河蚬的种内和种间竞争减小,个体的生存空间充裕,故该区域的河蚬规格较大。
河蚬的资源量往往随季节变化显著,而水温是影响河蚬资源量的又一个重要因素[21]。本研究结果显示三个站点河蚬生物量的最大值均出现在2016年7月,除马浪岗外最小值均为2016年3月,这与最高温度出现在2016年7月,而最低温度为2016年3月有着相同的变化规律,对三个站点的河蚬生物量和水温进行Pearson相关性分析,结果表明两者呈显著的正相关(r=0.584,P=0.046<0.05)。由于河蚬的最适生长温度为9~32℃[10],3月到7月之间均为河蚬生长的最佳季节,此时水体中的藻类随着水温的升高也大量增长,河蚬的饵料食物来源增多,河蚬生物量也增大,至11月水温逐渐降低,水体中的藻类也逐渐减少,河蚬生物量也降低,这与蔡炜[12]进行的太湖河蚬时空分布的研究结果一致。
综上所述,洪泽湖三个站点河蚬的规格和生物量尽管受时间和地点的影响显著,归结起来,其本质是受水温、饵料生物的变动,以及非法捕捞、非法采砂等人为因素的影响,而人为因素是影响自然水域增殖放流效果的主要因素。因此,在进行河蚬增殖的过程中,我们应从人为可控因素入手,呼吁相关部门加大管控力度,禁止非法捕捞、非法采砂、排放污水等行为。此外,河蚬的最适增殖放流密度对其增殖效果的影响并未在本文中涉及,有待进一步探究,而河蚬增殖的另一种方法网围增殖法将大大增加人为可控因素,有待进一步研究。
[1]庄平,宋超,章龙珍.长江口河蚬营养成分的分析与评价[J].营养学报,2009,31(3):304-306.
[2]韩鹏,王勤.河蚬软体部分营养成分分析及评价[J].厦门大学学报,2007,46(1):115-117.
[3]李亚成.洪泽湖河蚬通过农业部农产品地理标志登记专家评审 [EB/OL].http://www.hzh.gov.cn/detail.asp?artid=1903,2015-04-08/2016-03-06.
[4]刘敏,熊邦喜.河蚬的生态习性及其对重金属的富集作用[J].安徽农业科学,2008,36(1):221-224.
[5]袁永浒,王兴元,陈安来,等.洪泽湖螺蚬资源调查报告[J].水产养殖,1994(6):15-16.
[6]严维辉,潘元潮,郝忱,等.洪泽湖底栖生物调查报告[J].水利渔业,2007,27(3):65-66.
[7]张超文,张堂林,朱挺兵,等.洪泽湖大型底栖动物群落结构及其与环境因子的关系 [J].水生态学杂志,2012,33(3):27-33.
[8]李春荣,肖惠明,王荣华.河蚬移殖增殖实验 [J].科学养鱼,1996(5):24.
[9]张友奎,汪家忠.河蚬生态移殖优化增值技术总结[J].内陆水产,2003(2):12-13.
[10]邓勇辉,付辉云,陶志英,等.我国河蚬研究进展与展望[J].江西水产科技,2016(2):33-37.
[11]Fuiman L A,Gage J D,Lamont P A.Shell morphometry of the deep sea protobranch bivalve Ledella pustulosa in the Rockall Trough,north-east Atlantic[J].Journal of the Marine Biological Association of the UK,1999,79(4):661-671.
[12]蔡炜.太湖河蚬的时空分布及对N、P的贡献 [D].南京:南京信息工程大学,2010.
[13]Hornbach D J.Life history traits of a riverine population of the Asian clam Corbicula fluminea[J].American Midland Naturalist,1992:248-257.
[14]黄一鸣,尤玉博,颜金扬,等.河蚬(Corbicula fluminea Müller)的生态与食性的研究[J].福建师范大学学报(自然科学版),1979(2):99-105.
[15]李应森,杨仁民,滕利荣.黄蚬湖泊增养殖技术总结[J].水利渔业,1999,19(5):25-26.
[16]Mouthon J.Life cycle and population dynamics of the Asian clam Corbicula fluminea (Bivalvia:Corbiculidae)in the Saone River at Lyon (France)[J].Hydrobiologia,2001,452(1-3):109-119.
[17]闫云君,李晓宇.汉江流域黑竹冲河部分优势大型底栖动物的周年生产量 [J].水生生物学报,2007,31(3):297-306.
[18]李德亮,张婷,肖调义,等.湖南省大通湖河蚬(Corbicula fluminea)现存量及其时空分布 [J].湖泊科学,2013,25(5):743-748.
[19]毕婷婷,侯刚,张胜宇,等.基于壳长频率分析的洪泽湖河蚬渔获物年龄结构,生长与死亡参数估计[J].水生生物学报,2014,38(4):797-800.
[20]原居林,练青平,王凯伟,等.瓯江干流丽水段渔业资源现状分析与修复研究 [J].水生态学杂志,2010,3(5):122-126.
[21]蔡炜,蔡永久,龚志军,等.太湖河蚬时空格局[J].湖泊科学,2010,22(5):714-722.
猜你喜欢
华人时刊(2022年21期)2022-02-15 03:42:24
水上消防(2020年5期)2020-12-14 07:16:14
华人时刊(2020年13期)2020-09-25 08:21:40
文史春秋(2020年1期)2020-03-16 13:13:36
文史春秋(2019年11期)2020-01-15 00:44:36
智能城市(2018年7期)2018-07-10 08:30:00
中国继续医学教育(2015年2期)2016-01-06 01:36:16
食品工业科技(2014年15期)2014-03-11 18:17:50
振动、测试与诊断(2014年6期)2014-03-01 01:14:50
现代检验医学杂志(2014年1期)2014-02-06 01:29:31
