
中东大西洋中部海域鲐鱼渔场的时空变化初步研究
2018-03-10 02:56李显森逄志伟朱建成应一平
水产科学 2018年1期
李显森,逄志伟,朱建成,应一平,孙 珊
( 中国水产科学研究院 黄海水产研究所,农业部海洋渔业可持续发展重点实验室,山东 青岛 266071 )
鲐鱼(Pneumatophorusjaponicus)属于大洋暖水性中上层鱼类,主要分布于印度洋、太平洋和大西洋东部的温、热带海域,西非的温带和亚热带海域也是其重要的分布海域[1-2]。中国近海已经进行了诸多有关鲐鱼的研究,刘勇等[3]对东海北部和黄海南部鲐鱼生长特性进行了研究,官文江等[4]对东海南部海洋净初级生产力与鲐鱼资源量变动关系进行了研究,崔科等[5]对东黄海鲐鱼资源丰度与海表温度的关系进行了研究,李纲等[6-7]对夏季东海渔场鲐鱼产量与海洋环境因子的关系、东海鲐鱼资源和渔场时空分布特征进行了研究,并指出该海域渔场分布与该海域海表温度、海表高度和叶绿素a 含量关系密切,高峰等[8]基于提升回归树开发了东、黄海鲐鱼渔场预报模型。
位于非洲大陆西岸、大西洋东岸的联合国粮食与农业组织34渔区,其中的3.11和1.32小区东临毛里塔尼亚共和国,其地处热带,受加那利寒流和几内亚暖水及海洋信风等的影响下而涌升流显著,渔业资源,尤其是中上层鱼类资源相当丰富,盛产鲐鱼,是世界著名的渔场[2,9-10]。在对该渔场研究方面,国外学者对相关海域中上层鱼类的组成、多样性和生物学等进行了相关研究[11-12]。我国学者主要对该海域鱼类资源开发现状及前景、渔场及环境特征进行了研究[2,10,13-14],对佛得角海岸中心渔场方面的研究较少涉及,而对太平洋海域中心渔场方面的研究开展较多,已经涉及东南太平洋智利外海竹荚鱼(Trachurusmurphyi)[15-17]、北太平洋鱿鱼(Ommastrephesbartrami)[18]等。对比发现,有关中东大西洋鲐鱼研究相对较少,朱清澄等[1]对西非近海鲐鱼生物学特性的进行了研究。然而,对中东大西洋鲐鱼中心渔场及其影响因子方面的研究鲜有出现。
广义可加模型作为一种非线性关系模型,因能更深入地探讨渔场与影响因素间的关系以及确定各因素的重要程度,而在研究渔场方面得到了广泛应用[10,19-24]。本研究根据2012年1—8月中国大型拖网渔船在佛得角海岸作业的渔业数据以及遥感获取的海洋环境数据,运用广义可加模型对该海域鲐鱼中心渔场的时空变动及其影响因素进行分析,以期揭示鲐鱼中心渔场的季节变化规律,为合理利用与开发中东大西洋鲐鱼资源提供基础资料。
1 材料与方法
1.1 数据来源
研究数据来自于2012年1—8月上海开创远洋渔业有限公司、中国水产有限公司的大型拖网渔船在中东大西洋联合国粮食与农业组织34渔区3.11和1.32小区海域(北纬16°~22°,西经16°~19°)作业的鲐鱼产量数据。原始数据包括每次作业的日期、经纬度和网次产量等。另外,海表温度和叶绿素a 含量数据来自NASA水色遥感网站(http://oceans.gsfc.nasa.gov),研究海域为北纬16°~22°,西经16°~19°,研究时间为2012年1—8月。
1.2 数据处理与分析
由于本研究所涉及的大型拖网渔船主机功率基本相同(约5250 kW),均使用1440网型(网口周长1440 m,网总长360 m,网口目数60,网口网目尺寸24 m,网口网线直径12 mm,上纲长230 m)的中层拖网,拖速约5 n mile/h,因此可以认为捕捞能力基本相同。生产统计数据按月进行汇总,渔获产量的分布实际上代表了渔业资源量的空间分布和变化,渔业上常常采用渔场资源重心描述渔场空间位置的变动[15],本研究利用产量重心法研究中心渔场的空间变化。对生产统计资料进行数据标准化处理,按下式计算单位捕捞努力量渔获量(t/h)和月产量重心。

式中,CPUE为单位捕捞努力量渔获量(t/h); n为总网数;Ci为第i网的产量(t);h 为拖网时间(h);X、Y分别为产量重心的经、纬度;Xi为第i网中心点的经度,Yi为第i网中心点的纬度。
将获得的海表温度、叶绿素a含量数据和渔获数据按0.1°×0.1°网格重新抽取, 并根据空间位置进行匹配后并统计分析。
利用广义加性模型拟合渔获量与海洋环境因子(海表温度、叶绿素、经度、纬度)的关系[22], 建立以单位捕捞努力量渔获量为非独立变量、以主要的环境因子为解释变量的广义加性模型模型[23], 其表达式如下:
log(CPUE+0.001)=S(SST)+S [log(chl+0.001)]+S(longitude)+S (lattitude)+ ε
式中,S为自然立方样条平滑;SST 为海表温度;chl 为叶绿素a含量,为防止应变量出现零值,将单位捕捞努力量渔获量、chl加上0.001 后进行对数化的处理;longitude 为经度;latitude 为纬度;ε 为误差项。广义可加模型模拟结果中上、下95%的置信区间虚线离实线最接近的区间定义为渔场最适环境范围[23]。
2 结 果
2.1 鲐鱼平均单位捕捞努力量渔获量的月变化
鲐鱼平均单位捕捞努力量渔获量的计算表明,月平均单位捕捞努力量渔获量的范围为0.16~1.94 t/h,平均单位捕捞努力量渔获量最高的月份为1月,平均单位捕捞努力量渔获量最低的月份为4月。月平均单位捕捞努力量渔获量在1—8月基本呈现先减少,后增加的趋势。各月平均单位捕捞努力量渔获量为0.86 t/h,平均单位捕捞努力量渔获量较高的月份为1月和7月,其他月份的平均单位捕捞努力量渔获量较低(图1)。

图1 鲐鱼平均单位捕捞努力量渔获量的月变化
2.2 鲐鱼渔场的时空变化
通过对鲐鱼渔场产量重心的计算表明,该海域中心渔场的分布具有明显的月变化,鲐鱼中心渔场纬度方向的位移明显大于经度方向的位移,鲐鱼中心渔场位置基本呈现先往东南方向推移,且在4月到达最东南端,然后往西北推移趋势,直至7月到达最西北端;后再向东南推移的趋势。由2012年1—8月鲐鱼中心渔场的时空变化情况可以看出,1—3月渔场重心从(西经17.12°,北纬19.88°)向东南方向移动至渔场中心最东端(西经16.27°,北纬17.56°), 3—4月,渔场重心向南移动至最南端(西经16.39°,北纬17.44°);4—7月,作业渔场重心沿西北方向移动,7月份,渔场产量重心移动至海域的最西北端(西经17.35°,北纬20.28°),后渔场重心又往东南方向移动至(西经17.28°,北纬20.25°)(图2)。

图2 鲐鱼中心渔场时空变化
2.3 研究海域及鲐鱼渔场中心的叶绿素a含量、海表温度分布
鲐鱼渔场中心所在海域海表温度、叶绿素a含量的分布统计见图3、表1。不同月份渔场叶绿素a含量为0.23~10.00 mg/m3,渔场中心叶绿素a含量为3~8 mg/m3居多,叶绿素a含量的平均值3月、5月和7月较高,其他月份较低。综合看来,4月、5月和7月渔场中心的叶绿素a含量相对较低;其他月份渔场中心的叶绿素a含量相对较高,各月份渔场中心叶绿素a含量呈现一定的波动性。各月份海表温度平均值和渔场中心海表温度最高的为8月,最低的为8月。不同月份渔场中心的海表温度为17.2~27.6 ℃,除2月和8月外,渔场中心的海表温度相对较为稳定,渔场中心的海表温度为20~21 ℃居多。
2.4 广义可加模型模型分析
将所研究渔场的鲐鱼单位捕捞努力量渔获量与各影响因子(海表温度、叶绿素、经度、纬度)进行广义可加模型模型分析发现(图4),渔场的最适海表温度为19~22 ℃,最适叶绿素a含量为4.481~7.388 mg/m3,经度集中在西经16°30′附近,纬度集中在北纬18°30′位置附近。海洋环境与鲐鱼单位捕捞努力量渔获量的回归方程的显著性检验表明, 海表温度、叶绿素a含量和经度在单位捕捞努力量渔获量上的回归均极显著(P<0.01, 表2)且显著性强弱顺序依次为海表温度、经度和叶绿素a含量,而纬度在单位捕捞努力量渔获量上的回归不显著(P>0.05, 表2),海表温度在单位捕捞努力量渔获量上的回归显著性最强。广义可加模型调整后的决定系数r2为0.446,解释偏差为49.9%。


图3 海表温度、叶绿素a含量与鲐鱼渔场中心的分布叠加●渔场中心

指标1月2月3月4月5月6月7月8月叶绿素a含量mg/m3变化范围0.23~10.000.75~100.78~100.55~100.87~100.65~100.65~100.26~10平均值2.843.904.522.453.435.375.371.68渔场中心值5~66~78~93~42~37~83~49~10海表温度℃变化范围19.94~21.5216.95~17.8419.20~20.2119.76⁃21.1220.33~27.1216.78~24.3319.73~27.8023.11~32.64平均值20.8817.5419.6420.3420.8420.4722.8627.25渔场中心值20.4~20.617.2~17.319.5~19.620.0~20.120.5~20.620.6~20.821.6~21.827.4~27.6
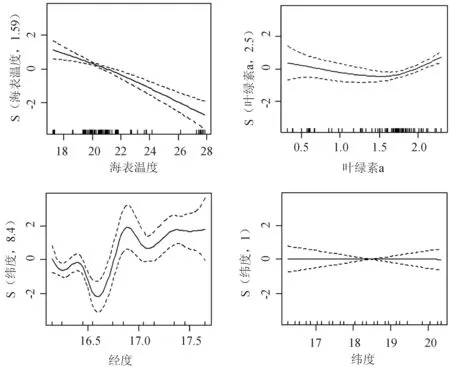
图4 鲐鱼渔获量与环境因子的广义可加模型模拟结果

环境因子有效自由度F检验P海表温度1.58626.3703.14×10-10∗∗∗叶绿素a2.5045.9347.43×10-4∗∗∗经度8.3994.9281.09×10-5∗∗∗纬度1.0000.0020.96
注:***表示差异极显著(P<0.01).
3 讨 论
中东大西洋联合国粮食与农业组织34渔区地理位置独特,南连热带赤道海洋,北接寒冷带海域。南部有向北运动的几内亚湾表面温暖洋流,北部有南移并上升的北大西洋加那利寒冷带深层洋流,冷暖洋流正好在此水域交汇, 形成显著的涌升流,形成了世界著名的渔场[25-26]。已有研究表明,洋流、水团与鲐鱼渔场关系密切,杨红等[27]认为东海夏季沿岸水团与外海水团分布特征及势力的强弱对鲐鱼渔场分布特征有重要影响,苗振清等[28]认为台湾暖流水舌锋进退对东海鲐鱼中心渔场变动具有重要影响且中心渔场通常随台湾暖流水舌锋的进退而移动。李纲等[7]通过分析鲐鱼作业海域的海表高度分析得到, 鲐鱼中心渔场通常位于海表高度极大值和极小值交汇的海域,即出现在冷水团和暖水团交汇区。联合国粮食与农业组织34渔区3.11和1.32小区鲐鱼中心渔场的分布具有明显的月变化,中心渔场基本呈现先往南推移,且在4月份到达最南端,后由南往北推移趋势,且中心渔场在纬度方向的位移明显大于经度方向的位移,这可能主要是由几内亚湾暖流、加那利寒流不同月份的强弱变化造成的。4月份之前,在偏北风影响下,北大西洋加那利寒冷带深层洋流较为强盛并南下[9,25],因此,此时鲐鱼渔场逐渐由北往南推移; 4月份之后,在向北移动的东北信风带等的驱动下,北大西洋加那利寒冷带深层洋流减弱,几内亚湾表面温暖洋流强盛,此时大陆架上盛行向北的海流[9,25],因此此时鲐鱼渔场逐渐由南往北推移。研究表明夏季东海海表温度、叶绿素a含量与鲐鱼产量关系密切,崔科等[5]研究认为东黄海鲐鱼资源4—5月产卵期的海表温度与当年南部作业渔场的单位捕捞努力量渔获量存在着显著的负相关关系,这些都与本研究结果海表温度和叶绿素a含量在鲐鱼单位捕捞努力量渔获量上的回归均极显著(P<0.01)相一致。
对于渔场的研究,了解潜在的渔场位置与环境变量的相互关系是研究的重要内容,通过同步分析海表温度与叶绿素a含量等物理化学因子能使我们更好地了解生物和海洋环境之间的关系[29]。本研究中,海表温度、叶绿素a含量和经度在鲐鱼单位捕捞努力量渔获量上的回归均极显著,且其明显高于该海域中上层鱼类单位捕捞努力量渔获量上的回归显著性水平[10],这可能主要与多鱼种对环境因子等的适应范围较单鱼种较广有关。本研究应用广义可加模型得出研究海域鲐鱼渔场的最适海表温度为19~22 ℃,最适叶绿素a含量为4.481~7.388 mg/m3,此海表温度和叶绿素a含量范围可以作为预报该海域潜在渔场的指标之一。已有研究指出,仅从叶绿素a含量和海表温度影像协同预报潜在渔场位置的方法由于无法全面反映中尺度涡、海洋锋、水团等因素而显得仍有不足,因此在研究中要注意中尺度涡和海洋锋等现象对渔场的影响[6]。另外,崔科等[5]研究认为东黄海鲐鱼资源每年3—4月Elnio3.4A指标与次年南部作业渔场单位捕捞努力量渔获量之间存在着显著的正相关关系,已有研究表明鲐鱼的资源变动受厄尔尼诺、南方涛动等大气候事件的影响明显[30-31]。本研究由于受所研究时间尺度等的影响,未能对重大的气候现象对鲐鱼中心渔场的影响进行深入研究,这也是今后中东大西洋中部该海域渔场研究的重要方向。因此,在研究渔场时,不仅需要研究各种物理化学因素,对于洋流以及各种大气候事件的影响也不容忽视。
综上所述,佛得角海岸联合国粮食与农业组织34渔区3.11小区鲐鱼中心渔场的时空变化与几内亚湾暖流、加那利寒流等洋流不同月份的强弱变化关系密切。因此,在研究该海域鲐鱼中心渔场的时空变化时,一定要结合海表温度、叶绿素a含量等环境指标,加强加那利寒冷带深层洋流、几内亚湾表面温暖洋流等洋流的研究,注意这些洋流的强弱变化对该海域鲐鱼中心渔场的影响。
4 结 论
笔者对中东大西洋中部海域联合国粮食与农业组织34渔区3.11和1.32小区2012年1—8月鲐鱼的渔获量统计资料与主要环境因子关系进行的研究, 发现其中心渔场的分布与海表温度、叶绿素a含量和经度等有一定联系, 并通过广义可加模型模型得出渔场最适海表温度为19~22 ℃和最适叶绿素a含量为4.481~7.388 mg/m3(此海表温度和叶绿素a含量可作为预报鲐鱼潜在渔场的指标之一), 以及集中的时空范围, 经度集中在西经16°30′附近,纬度集中在北纬18°30′位置附近。研究可为国内外相关渔业生产单位在西非海域的渔业生产提供渔场变化规律和渔情信息的参考。
[1] 朱清澄, 张衍栋, 陈丙见, 等. 西非近海鲐鱼生物学特性的初步研究[J].海洋湖沼通报, 2014(2):31-36.
[2] 张禹, 张勋, 周爱忠, 等. 毛里塔尼亚海域小型中上层鱼类资源开发现状及前景分析[J]. 现代渔业信息, 2011, 26(6):3-5.
[3] 刘勇, 严利平, 程家骅.东海北部和黄海南部鲐鱼生长特性及合理利用[J]. 中国水产科学, 2006,13(5):814-822.
[4] 官文江, 陈新军, 高峰, 等. 东海南部海洋净初级生产力与鲐鱼资源量变动关系的研究[J]. 海洋学报, 2013, 35(5):121-127.
[5] 崔科, 陈新军. 东黄海鲐鱼资源丰度与表温关系[J]. 南方水产, 2007,3(4):20-25.
[6] 李纲, 陈新军.东海鲐鱼资源和渔场时空分布特征的研究[J]. 中国海洋大学学报:自然科学版, 2007, 37(6):921-926.
[7] 李纲, 陈新军.夏季东海渔场鲐鱼产量与海洋环境因子的关系[J]. 海洋学研究,2009, 27(1):1-8.
[8] 高峰, 陈新军, 官文江, 等. 基于提升回归树的东、黄海鲐鱼渔场预报[J]. 海洋学报, 2015, 37(10):39-48.
[9] 韩保平, 方海, 阮雯. 毛里塔尼亚海洋渔业概况[J]. 现代渔业信息, 2011,26(4):20-23.
[10] 逄志伟, 李显森, 朱建成, 等. 中东大西洋中部海域中上层鱼类中心渔场的时空变化[J]. 生态学杂志, 2016, 35(11):3072-3079.
[11] Ter-Hofstede R, Dickey-Collas M T, Mantingh I, et al. The link between migration, the reproductive cycle and condition ofSardinellaauritaoff Mauritania, north-west Africa[J]. Journal of Fish Biology, 2007, 71(5):1293-1302.
[12] Wienerroither R, Uiblein F, Moreno F B T. Composition, distribution, and diversity of pelagic fishes around the Canary Islands, Eastern Central Atlantic[J]. Marine Biology Research, 2009, 5(4):328-344.
[13] 张禹, 张勋, 周爱忠, 等. 西北非海域小型中上层渔业资源现状与发展趋势[J]. 现代渔业信息, 2009, 24(12):16-19.
[14] 冯春雷, 郁岳峰, 黄洪亮, 等. 毛里塔尼亚海域渔场的环境特征[J]. 海洋渔业, 2008,30(3):219-226.
[15] 化成君, 张衡, 樊伟. 东南太平洋智利竹筴鱼资源和渔场的时空变化[J]. 生态学报, 2011, 31(19):5676-5681.
[16] 牛明香, 李显森, 徐玉成. 智利外海竹筴鱼中心渔场时空变动的初步研究[J]. 海洋科学, 2009, 33( 11):105-110.
[17] 邹晓荣. 东南太平洋智利竹筴鱼资源、渔场和捕捞技术的研究[D]. 上海: 上海海洋大学, 2003.
[18] 沈新强, 王云龙, 袁骐, 等. 北太平洋鱿鱼渔场叶绿素 a 分布特点及其与渔场的关系[J]. 海洋学报, 2005, 26(6):118-123.
[19] HastieT J,Tibshrani R J.Generalized Additive Models [M]. London: Chapman and Hall, 1990.
[20] Maunder M N, Punt A E. Standardizing catch and effort data: a review of recent approaches[J]. Fisheries Research, 2004, 70(2/3):141-159.
[21] Stenseth N C, Mysteruda D A, Ottersen G, et al. Ecological effects of climate fluctuations[J]. Science, 2002(297):1292-1296.
[22] 樊伟, 崔雪森, 沈新强. 西北太平洋巴特柔鱼渔场与环境因子关系研究[J]. 高技术通讯, 2004(10):84-89.
[23] 朱国平, 朱小艳, 徐怡瑛, 等. 基于GAM 模型分析夏秋季南奥克尼群岛南极磷虾渔场时空分布及与环境因子之间的关系[J]. 极地研究, 2012, 24(3):266-273.
[24] 孙珊,李显森,赵文武,等.基于广义可加模型分析时空和环境因子对2013年南极磷虾资源分布的影响[J]. 水产科学,2015,34(10):609-615.
[25] 吴廷辉. 大西洋[M]. 上海: 商务印书馆, 1982.
[26] 张进宝. 毛里塔尼亚的渔业[J]. 世界农业, 2002(1):33-35.
[27] 杨红, 章守宇, 戴小杰, 等. 夏季东海水团变动特征及对鲐鲹渔场的影响[J]. 水产学报, 2001,25(3):209-214.
[28] 苗振清. 东海北部近海夏秋季鲐鲹渔场与海洋水文环境的关系[J]. 浙江水产学院学报,1993,12(1):32-39.
[29] Wang W. On the relationship between the resources of ommastrephes bartrami and marine environment in the northwest Pacific Ocean based on GIS[J]. Geoinformation Science, 2003,5(1):39-44.
[30] Yatsu A, Watanabe T, Ishida M, et al. Environmental effects on recruitment and productivity of Japanese sardineSardinopsmelanostictusand chub mackerelScomberjaponicuswith recommendations for management[J]. Fisheries Oceanography, 2005,14(4):263-278.
[31] Sun C H, Chiang F S, Tsoa E, et al. The effects of El Nio on the mackerel purse-seine fishery harvests in Taiwan: an analysis integrating the barometric readings and sea surface temperature[J]. Ecological Economics, 2013, 49(8):4724-4729.
猜你喜欢
海洋技术学报(2021年5期)2021-12-30
中国水产(2021年12期)2021-12-06
海洋学报(2021年1期)2021-03-02
大自然探索(2021年12期)2021-02-07
阅读(科学探秘)(2020年8期)2020-11-06
海洋信息技术与应用(2020年2期)2020-07-27
热带海洋学报(2020年3期)2020-05-25
绿色科技(2019年2期)2019-05-21
当代水产(2019年4期)2019-05-16
中国果业信息(2019年1期)2019-01-05
