
饲喂频率对生长猪饲粮养分消化率的影响
2018-03-07 06:52曹山川陈冠希谢乙宁刘静波
中国畜牧杂志 2018年2期
刘 静,曹山川,陈冠希,谢乙宁,刘静波
(西南科技大学生命科学与工程学院,四川绵阳 621010)
随着养猪生产工业化进程的加快,饲养方式也由传统的限制饲喂频率模式(每日饲喂2~3次)逐步过渡到高摄食频率的自由采食模式(每日采食15次左右)[1],饲喂频率的变化是否影响了饲粮养分的消化率以及可消化养分在体内的代谢过程目前报道较少。Chastanet等[2]研究指出,当生长猪日采食量控制在维持需要的3倍时,自由采食组与每日饲喂1~2次比较,其总能(GE)和干物质(DM)的表观全肠道消化率(ATTD)显著降低,自由采食组的日采食量显著高于限制饲喂频率组,而采食量的提高可能是造成养分消化率降低的主要因素,而非饲喂频率。Ruckebusch等[3]研究表明,与自由采食组相比,每天饲喂1~2次对养分消化率无负面影响,但生长猪小肠和胃的能动性显著降低。Friend等[4]研究表明,在限制饲喂条件下,每天饲喂1次或5次对生长育肥猪粗蛋白(CP)和DM的全肠道消化率无显著影响。早期研究中高饲喂频率模式基本是每日饲喂2~5次,与生长猪自由采食条件下的采食频率(每日15次)相差较大[1-3]。Le Naou等[5]和Liu等[6]研究指出,当饲喂频率从每天2次增加到12次时,生长猪的糖脂代谢发生显著变化,主要原因可能在于增加饲喂频率改变了饲粮的养分消化率。因此,本次试验在接近自由采食水平的条件下,比较生长猪每天饲喂2次或12次时的养分消化率差异。
1 材料与方法
1.1 试验设计和饲养管理 试验选用24头DLY三元杂交去势公猪,平均初始体重为(40.7±1.3)kg,并在回肠末端安装简单T型瘘管。根据试验动物的初始体重按完全随机区组设计分为2个处理组,每个处理组12个重复,每个重复1头猪。各处理组的饲粮相同,高饲喂频率组每天饲喂12次(M12),限制饲喂频率组每天饲喂2次(M2)。根据NRC(2012)猪营养需要配制本次试验需要的玉米-豆粕型饲粮,试验饲粮的组成和营养成分见表1。
在饲粮中添加0.5%的三氧化二铬作为外源指示剂,计算GE、粗灰分(Ash)、DM、粗脂肪(EE)、碳水化合物(CHO)、CP、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)等的表观回肠消化率(AID)和ATTD。试验期间生长猪的平均日采食量根据初始体重的5%计算得出,每次饲喂量由日采食量除以每日饲喂频率计算得出。M2组在每天08:00和18:00进行饲喂,未采食完的饲粮在10:00和20:00取出烘干后称重记录剩余量。M12组每隔2 h饲喂1次,每天08:00开始饲喂,次日07:00取出未采食完的饲粮烘干后称重记录剩余量。试验期间将生长猪单独饲养在代谢笼内,代谢笼配有不锈钢料槽和低压饮水器,维持(20±2)℃的室温和每日12 h的光照。根据试验开始和结束时的动物体重计算平均日增重(ADG),根据试验期间每头猪的饲粮消耗量计算平均日采食量(ADFI),而耗料增重比(F/G)为ADFI和ADG的比值。
1.2 样品收集 整个试验期为9 d,包括5 d适应期,2 d(第6天和第7天)粪便样品收集期,2 d(第8天和第9天)食糜样品收集期,每天收集08:00—18:00的全部样品。在食糜收集期间,样品袋每1 h更换1次,每次收集的粪样和食糜保存在-20℃条件下。试验结束后,将收集的每头猪的粪样和食糜样分别混合均匀待用,粪样在65℃条件下烘干,食糜样品冷冻干燥后粉碎,将处理好的样品保存好待测。

表1 试验日粮组成和营养成分(风干基础)
1.3 指标测定与方法 饲粮、粪样和回肠食糜的DM、EE、CP、Ash、NDF和ADF含量分别按照国标GB/T6435-2014、GB/T 6433-2006、GB/T 6432-94、GB/T 6438-2007、GB/T 20806-2006和 GB/T 20805-2006的方法测定。用全自动热量分析仪(Model 6400;Parr Instrument,Moline,IL)测定饲粮、粪样以及回肠食糜GE含量。按照国标GB/T 18246-2000方法测定氨基酸含量,将处理后的样品用氨基酸自动分析仪测定(L-8800 Hitachi,Tokyo,Japan)。样品处理:样品用过氧甲酸氧化过夜后在7.5 mol/L HCl中110℃条件下水解24 h后测定蛋氨酸和半胱氨酸的含量;样品在LiOH中110℃条件下水解22 h后测定色氨酸的含量;剩余15种氨基酸含量,样品在6 mol/L HCl中110℃条件下水解24 h后测定。
将饲粮、粪样和回肠食糜用浓硝酸和高氯酸消化处理后用比色法分别测定铬含量。饲粮、粪样和回肠食糜中CHO含量按公式计算:CHO = DM-(CP+EE+Ash)。
养分和GE的AID 和ATTD 按公式计算:

其中,Crdiet、Crilealdigesta和Crfeces分别表示饲粮、回肠食糜和粪样中的铬浓度;Ndiet、Nilealdigesta和Nfeces分别表示饲粮、回肠食糜和粪样中养分和GE含量;后肠养分消化率由以下公式计算得出:
后肠养分消化率=ATTD-AID
1.4 统计分析 数据用SAS统计分析软件的MIXED程序统计分析数据。饲喂频率作为固定效应,体重为随机效应。当P<0.05时认为差异显著;当 0.05<P≤0.1时认为有趋势。
2 结果与分析
2.1 饲喂频率对猪生长性能的影响 由表2可知,因各处理组的初始体重无显著差异(P>0.1),且各处理组的ADFI也无显著差异(P>0.1)。另外,饲喂频率对生长猪的ADG、试验结束体重以及F/G均无显著影响(P>0.1)。
2.2 饲喂频率对养分和GE的AID的影响 由表3可知,饲喂频率不影响生长猪的CHO、DM、EE、CP、Ash、NDF、ADF以及GE的AID(P>0.1)。
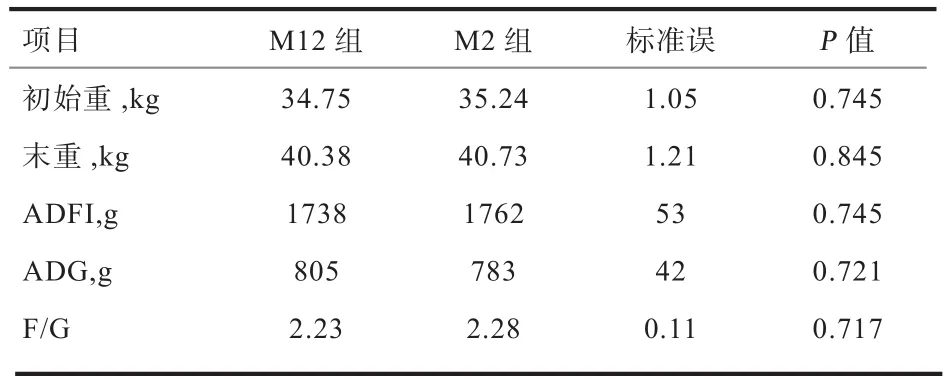
表2 饲喂频率对猪生长性能的影响
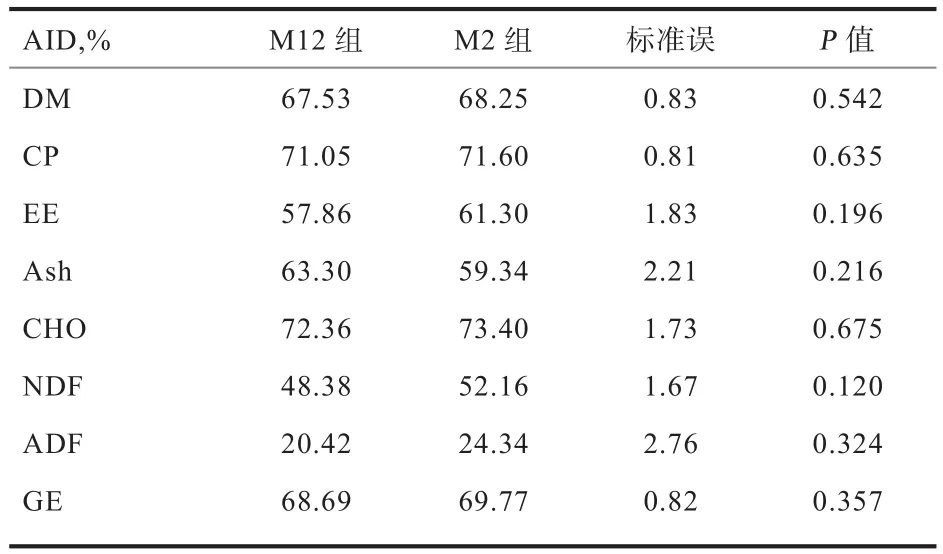
表3 饲喂频率对养分和GE的AID的影响
2.3 饲喂频率对养分和GE的后肠消化率的影响 由表4可知,M2组的EE、ADF和NDF的后肠消化率显著低于M12组(P<0.05),其中EE的后肠消化率由11.54%降到2.50%,并且NDF降低了9.34个百分点。M2组GE的后肠消化率有低于M12组的趋势(P<0.1),由15.03%降到11.95%。饲喂频率对CHO、DM、Ash和CP的后肠消化率无显著影响(P>0.1)。

表4 饲喂频率对养分和GE后肠消化率的影响
2.4 饲喂频率对养分和GE的ATTD的影响 由表5可知,M2组的GE、DM、Ash、EE和NDF的ATTD显著低于M12组(P<0.05),其中GE、DM和EE的ATTD分别降低1.99%,1.82%和5.59%。与高饲喂频率组相比,限制饲喂频率组ADF和CP的ATTD分别降低了11.2%,3.04%,有降低的趋势(P<0.1)。饲喂频率对CHO的ATTD无显著影响(P>0.1)。

表5 饲喂频率对养分和总能ATTD的影响
2.5 饲喂频率对氨基酸AID的影响 由表6可知,饲喂频率对生长猪的精氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、蛋氨酸、苯丙氨酸、苏氨酸、色氨酸、缬氨酸、丙氨酸、天冬氨酸、半胱氨酸、谷氨酸、甘氨酸、脯氨酸、丝氨酸和酪氨酸的AID无显著影响(P>0.1)。
3 讨 论
本试验将各处理组的日采食量设定在相同水平(日采食量按初始体重的5%计算,该水平接近自由采食的水平),目的在于使M12组(从每天08:00开始进行饲喂,每次间隔2 h,每次饲喂量为日采食量的1/12)和M2组(每天08:00和18:00饲喂,每次饲喂量为日采食量的1/2)的ADFI一致。本试验研究结果表明,当ADFI接近自由采食的条件下,增加饲喂频率显著提高了GE和养分的ATTD,但饲喂频率对GE和养分的AID影响不显著,由此可见,增加饲喂频率能提高养分在大肠中的发酵程度。增加饲喂频率虽然能提高养分的消化率,但是对各试验处理组的动物的生长性能并未造成影响。
Hatori等[7]和Sherman等[8]研究一致表明,饲喂高脂或正常饲粮,增加饲喂频率都可使大鼠的ADG显著提高。但前期关于饲喂频率对猪生长性能影响的研究结果不一致,Allee等[9]研究表明,饲喂频率的降低对生长猪的ADG和饲料报酬有显著改善作用。在相同的ADFI的条件下,饲喂频率从每天2次提高到6次时,对妊娠母猪和后备母猪的ADG和饲料报酬有显著改善作用[10-11]。然而在本试验中,并未观察到饲喂频率对生长猪的ADG和饲料报酬有显著影响,主要原因可能是试验周期太短,以至观察不到试验主效应对生长性能的影响。此外,本试验结果和前期研究结果不一致的主要原因可能是不同试验间选取的试验动物处于不同的生长阶段,而试验动物在不同的生长阶段,其脂肪和蛋白质的沉积能力有显著差异。
Ruckenbusch等[3]、Hee等[12]以及 de Haer等[13]的研究结果一致表明增加饲喂频率能改善动物养分消化率。增加饲喂频率如何改善养分消化率的机理可能主要涉及以下几方面:增加饲喂频率可以促进肠道蠕动[3];增加饲喂频率可使小肠消化液的分泌增加[3];增加饲喂频率可以促进胰腺的分泌[12-13]。Ruckenbusch等[3]研究表明,限制饲喂频率与自由采食相比,每天只饲喂1次或2次会显著降低生长猪小肠和胃的蠕动能力。但本试验与Mroz等[14]以及Chastanet等[2]的研究结果一致表明,GE和养分的回肠消化率并没有因为限制饲喂频率对生长猪前肠道能动性的负面作用而降低。每天饲喂生长猪2次或12次对CHO、DM、EE、CP、Ash、ADF、NDF以及GE的AID 影响并不显著。虽然Ruckenbusch等[3]的研究结果表明增加饲喂频率可以促进小肠消化液和胰腺的分泌,但是在正常生理范围内,生长猪养分消化率的增加是否与消化液分泌量的增加存在显著相关性还需要更进一步研究验证。另外,因为肠道蠕动活力提高和消化液分泌量增加而导致养分的内源性损失增加以及维持需要的增加,所以消化液分泌量增加对养分消化率改善的积极作用被抵消,最终导致饲喂频率对养分的回肠消化率无显著影响。
Chastanet等[2]研究表明,限制饲喂频率组(每天饲喂1次或2次)的GE和DM在大肠的消化率显著高于自由采食组,限制饲喂频率组的日采食量(每天饲喂1次:1 644 g/d;每天饲喂2次:1 632 g/d)显著低于自由采食组(2 730 /d)是造成上述结果的主要原因。养分在小肠主要依靠酶水解,在大肠主要依靠微生物作用。大肠微生物长期处于平衡状态,自由采食组日采食量过多,从而可能超过了微生物的降解能力,导致自由采食组GE和DM的全肠道消化率显著降低。这与Haydon等[15]的研究结果一致。在本试验条件下,增加饲喂频率能显著改善生长猪的DM、CP、EE、Ash、ADF、NDF和 GE的 ATTD以及 GE、EE 、ADF和 NDF的后肠道消化率。由上述结果可见,因CHO容易被消化吸收,所以一般在前肠段被小肠消化液降解吸收,而ADF和NDF主要在大肠靠大肠微生物的发酵作用消化吸收。本试验第一次明确提出,增加饲喂频率可显著提高CP、ADF和NDF在后肠的发酵作用,从而提高GE和DE的ATTD。肠道微生物菌群的组成以及其在肠黏膜上的定植显著受食糜在肠道的排空速度影响,而影响后肠道养分消化吸收的重要因素之一就是肠道微生物菌群的组成。Ruckenbusch等[3]明确指出,饲喂频率对食糜在肠道的排空速度和肠道蠕动有显著影响。所以,提高养分消化率是否可通过上述过程改变微生物菌群组成还需要进一步研究。饲喂频率对养分消化率的影响机理有很多,前期研究主要涉及消化液的分泌和小肠的蠕动,而对ADF和NDF的发酵过程和后肠道微生物菌群的组成涉及较少。从本试验的研究结果可知,饲喂频率对养分的后肠道消化率有显著影响,但上述过程是否涉及肠道微生物的作用还需进一步研究验证。
4 结 论
饲喂频率对生长猪GE和养分的AID无影响,增加饲喂频率虽然能提高GE和养分的后肠道消化率和ATTD,但对生长猪的生长性能无显著的影响。由此可见,增加饲喂频率可显著提高养分在生长猪后肠道的发酵程度。
[1] Colpoys J D, Johnson A K, Gabler N K. Daily feeding regimen impacts pig growth and behavior[J]. Physiol Behave, 2016,159:27-32.
[2] Chastanet F, Pahm A A, Pedersen C,et al. Effect of feeding schedule on apparent energy and amino acid digestibility by growing pigs[J]. Anim Feed Sci Technol, 2007, 132: 94-102.
[3] Ruckebusch Y, Bueno L. Effect of feeding on motility of stomach and small-intestine in pig[J]. Br J Nutr, 1976, 35: 397-405.
[4] Friend D W, Cunningham H. Effects of feeding frequency on metabolism, rate and efficiency of gain and on carcass quality of pigs[J]. J Nutr, 1964, 83: 251-256.
[5] Le Naou T, Le Floc’h N, Louveau I,et al. Meal frequency changes the basal and time-course profiles of plasma nutrient concentrations and affects feed efficiency in young growing pigs[J]. J Anim Sci, 2014, 92:2008-2016.
[6] Liu J B, Liu Z H, Chen L,et al. iTRAQ-based proteomic analysis reveals alterations in the liver induced by restricted meal frequency in a pig model[J]. Nutrition, 2016, 32: 871-876.
[7] Hatori M, Vollmers C, Zarrinpar A,et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet[J]. Cell Meta, 2012, 15(6): 848-860.
[8] Sherman H, Frumin I, Gutman R,et al. Long-term restricted feeding alters circadian expression and reduces the level of inflammatory and disease markers[J]. J Cell Mol Med, 2011,15: 2745-2759.
[9] Allee G L, Romsos D R, Leveille G A. Metabolic adaptation induced by meal-eating in the pig[J]. J Nutr, 1972, 102: 1115-1122.
[10] Schneider J D, Tokach M D, Dritz S S,et al. The effects of feeding schedule on body condition, aggressiveness, and reproductive failure in group housed sows[J]. J Anim Sci, 2007,85: 3462-3469.
[11] Schneider J D, Tokach M D, Goodband R D,et al. Effects of restricted feed intake on finishing pigs weighing between 68 and 114 kilograms fed twice or 6 times daily[J]. J Anim Sci,2011, 89: 3326-3333.
[12] Hee J, Sauer W C, Mosenthin R. The effect of frequency of feeding on the pancreatic secretions in the pig[J]. J Anim Physiol Anim Nutr, 1988, 60: 249-259.
[13] de Haer L C M, de Vries A G. Effects of genotype and sex on the feed intake pattern of group housed growing pigs[J]. Livest Prod Sci, 1993, 36: 223-232.
[14] Mroz Z, Jongbloed A W, Kemme P A. Apparent digestibility and retention of nutients bound to phytate complexes as in fl uenced by microbial phytase and feeding regimen in pigs[J].J Anim Sci, 1994, 72: 126-132.
[15] Haydon K D, Knabe D A, Tanksley T D. Effects of level of feed intake on nitrogen, amino acid and energy digestibilities measured at the end of the small intestine and over the total digestive tract of growing pigs[J]. J Anim Sci, 1984, 59: 717-724.
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
中国土壤与肥料(2022年4期)2022-06-14
中国饲料(2021年17期)2021-11-02
中国畜牧杂志(2021年5期)2021-05-17
落叶果树(2021年6期)2021-02-12
兽医导刊(2020年11期)2020-12-28
国际种业前沿动态(2020年16期)2020-10-14
三农资讯半月报(2020年16期)2020-09-14
文苑(2020年12期)2020-04-13
