
几种沙生灌木光合作用及水分生理特征比较
2018-03-01 21:51何苗宁虎森罗青红吉小敏雷春英
防护林科技 2018年1期
何苗,宁虎森,罗青红,吉小敏,2,雷春英,2
(1.新疆林科院造林治沙研究所,新疆 乌鲁木齐 830063;2.新疆精河荒漠生态系统国家定位观测研究站,新疆 博州 833300)
全球气候变暖,土地沙化已成为人类社会所面临的最严重的生态环境问题之一。截至2014年,全国沙化土地面积为172.12万km2,新疆沙化土地和具有明显沙化趋势的土地面积达74.71万km2和4.71万km2,分别占全国的43.4%和15.68%[1]。近年来,新疆实施一系列防沙治沙工程项目,取得了一定的成果,但局部地区沙漠化发展扩大和风沙危害严重的态势仍未得到根本遏制。运用植物材料治沙是“生物治理技术”的主要内容,是实现沙漠治理的最有效途径。多年研究与实践也证明,我国西部干旱、半干旱区,真正能发挥作用的植物一般为多年生灌木、半灌木和草本。但沙化土地分布区自然环境条件恶劣,筛选适应沙区环境的植物就成为生物治沙的重要环节。
关于耐旱植物生理生态适应性,前人已做了大量研究工作。尹立河等[2]通过比较毛乌素沙漠4种植物水势特征,得出不同植物叶水势的变化因种类、季节、气候条件的不同而异,但净辐射量是主要影响因子。周瑞莲等[3]通过分析不同生境中沙生植物体内的抗氧化酶活力和渗透调节物质,得出不同科属植物维持叶片水分和氧自由基代谢平衡可能是其适应沙漠环境生存的重要生理调控机理。谭永芹等[4]通过压力-容积法测定了5种干旱区木本植物的水分参数,蒸腾速率,水势和渗透势等指标,并进行了综合评价和抗旱能力排序。赵哈林等[5]通过研究风吹和沙埋处理下小叶锦鸡儿光合作用参数,指出中度胁迫时小叶锦鸡儿可以通过生长与生理调节来适应环境,当重度胁迫时,会导致固沙植物的死亡。
植物的幼苗阶段对外界环境的变化较为敏感[6],因此,本文将在前人研究的基础上,以两年生沙生植物沙地桑、沙木蓼、柠条、红皮沙拐枣为研究对象,通过比较各物种间光合作用特征参数、叶绿素含量和水势的变化,探究4种植物光合生理过程对沙地环境的响应适应机制以及种间光合生理生态特征参数的差异,旨在为新疆荒漠地区提供适宜造林树种,同时也为荒漠化退化生态系统的恢复和保护提供理论依据。
1 试验设计与研究方法
1.1 研究区概况
试验区位于古尔班通古特沙漠西南缘精河县的沙丘道班(83°32′ N、44°57′ E),属典型温带大陆性气候,年均日照时数达2 709.6 h;年均气温7.2 ℃,年均降水量102 mm,年均蒸发量1 423.9 mm,无霜期171 d。试验区土壤为沙土,地表以下0~60 cm土层土壤pH值为7.96,有机质为2.22 g·kg-1,全N、全P、全K分别为0.12 g·kg-1、0.28 g·kg-1、14.58 g·kg-1。
1.2 试验设置
选取沙木蓼(Atraphaxisbracteata)、沙地桑(Morusalba)、柠条(Caraganakorshinskii)和红皮沙拐枣(Calligonumrubicundum)的2年生健康幼苗作为试验材料。试验植物按照0.5 m×1.0 m的株行距栽植,每种植物栽3个小区,随机排列作为重复。小区边缘埋深2 m的塑料膜作为隔水带,每个小区的西头安装供水管道、控水阀门和计量装置以统一灌溉制度,每隔15 d灌水一次,灌溉量为12 000 m3·hm-2,并采用除草等田间管理措施。
1.3 测定指标与方法
1.3.1 光合—光强响应曲线测定方法 选择晴朗日,于9:00—11:00测定。先进行预试验,使用CIRAS-2的CO2控制系统,将CO2浓度控制在(380±10)μmol·mol-1,叶室相对湿度控制在45%~55%,叶温控制在(30±2) ℃,预试验之后,利用LED人工光源控制叶室光合有效辐射,设置PAR为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、100、50、0 μmol·m-2·s-1共14个梯度,每隔2 min记录下响应的光合参数值,进行光响应曲线的拟合。光合-光响应曲线采用直角双曲线修正模型[7]。
1.3.2 植物组织水势测定 用PMS1000型便携式植物水势压力室对4树种的枝条水势进行测定,测定时间选在7:00、14:00和21:00。测定时,用修枝剪快速剪下树冠阳面中部的初生枝条,放入仪器装置,观察第一滴组织液渗出时的压力,此时的压力值即为植物的茎水势值。每种植物每个小区重复3次取值,并将3个小区作为组内重复。
1.3.3 叶片叶绿素含量测定 叶片叶绿素含量的测定与光合参数测定同时进行,每个树种分别选取10片健康叶片,用叶绿素测定仪读取SPAD值。
1.4 数据统计分析
用Excel 2007、SPSS 20.0进行数据整理分析,单因素方差分析和最小显著差数法进行方差分析和差异显著性比较,用Origin7.5软件绘图。
2 结果与分析
2.1 4种植物水势日变化特征

图1 4种植物水势日变化特征
如图1所示,除沙拐枣外,其他3种植物日水势均表现为“V”形变化趋势,即早晚高,中午低。沙拐枣水势日变化呈逐渐降低的趋势,但中午后,降低趋势减缓。7:00时,4种植物水势大小依次为沙地桑>红皮沙拐枣>柠条>沙木蓼;21:00时4种植物水势依次为沙地桑>沙木蓼>红皮沙拐枣>柠条。
2.2 4种植物叶绿素含量比较
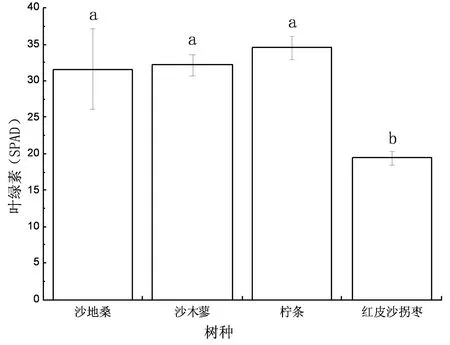
图2 4种植物叶绿素含量
如图2所示,4种植物中柠条叶绿素含量最高,其余按高到低依次为沙木蓼、沙地桑、红皮沙拐枣。沙地桑、沙木蓼、柠条叶绿素含量无显著差异(P>0.05)。红皮沙拐枣与其他3种植物叶绿素含量之间差异显著(P<0.05)。
2.3 4种植物光合作用—光强响应曲线特征
图3显示,4种沙生植物的Pn随着光强的增大表现出相似的变化规律。在光强为0~200 μmol·m-2·s-1时,Pn呈线性增大趋势,待光强上升到一定程度,Pn上升趋势减缓,直到达到光饱和点,而当光强继续增加时,Pn趋于稳定甚至小幅下降,即当光强超过饱和光强时,出现了光抑制现象。

图3 4种植物净光合速率、蒸腾速率、气孔导度、水分利用效率-光合有效辐射应曲线
4种植物叶片蒸腾速率(Tr)对光强的响应曲线以一元二次方程拟合较为理想(表1),R2值均大于0.9。由图3可以看出,4种植物在光强为0时,Tr的大小顺序为沙地桑>柠条>沙木蓼>红皮沙拐枣,当光强逐渐升高至2 000 μmol·m-2·s-1时,沙地桑、沙木蓼、柠条、红皮沙拐枣的蒸腾速率值分别为6.57 mmol·H2O·m-2·s-1、6.27 mmol·H2Om-2·s-1、6.27 mmol·H2Om-2·s-1、4.33 mmol·H2O·m-2·s-1。整体上看,沙地桑蒸腾作用最强,柠条,沙木蓼次之,红皮沙拐枣蒸腾作用最弱。
4种植物叶片的气孔导度(Gs)对光强的响应曲线也遵循一元二次方程变化,R2值均大于0.9(表1)。随着光强的增加,植物气孔逐渐打开(图3),但总体上来看,Gs随光强变化较小。四种植物中,柠条气孔导度最高,沙木蓼和沙地桑次之,红皮沙拐枣气孔导度最低。
如图3所示,水分利用效率-光强响应曲线与净光合速率-光强响应曲线变化规律相似,即在低光强时曲线呈线性增加,且4种植物水分利用效率差异不明显,随着光强的增加,曲线上升趋势逐渐减缓,同时各植物种间差异逐渐显现,当光强高于1 200 μmol·m-2·s-1时,水分利用效率大小依次为红皮沙拐枣>柠条>沙地桑>沙木蓼。

表1 4种植物光合作用参数对光强响应曲线回归方程
通过直角双曲线修正模型计算Pn-PAR曲线特征参数,结果如表2所示,4种植物的表观量子效率大小顺序为沙地桑>柠条>沙木蓼>红皮沙拐枣,且沙地桑表观量子效率显著高于其他3种植物(P<0.05)。最大净光合速率(Pnmax)大小依次为柠条>沙地桑>红皮沙拐枣>沙木蓼,除沙地桑与红皮沙拐枣外,各植物种间差异均达到显著水平(P<0.05)。光饱和点大小依次为红皮沙拐枣>沙地桑>柠条>沙木蓼,除沙地桑与柠条外,各植物种间差异均显著(P<0.05)。光补偿点大小依次为柠条>红皮沙拐枣>沙木蓼>沙地桑,柠条与红皮沙拐枣差异不显著(P>0.05),但显著高于沙木蓼和沙地桑(P<0.05)。暗呼吸速率大小依次为柠条>红皮沙拐枣>沙木蓼>沙地桑,其中柠条暗呼吸速率显著高于其他3种植物(P<0.05)。
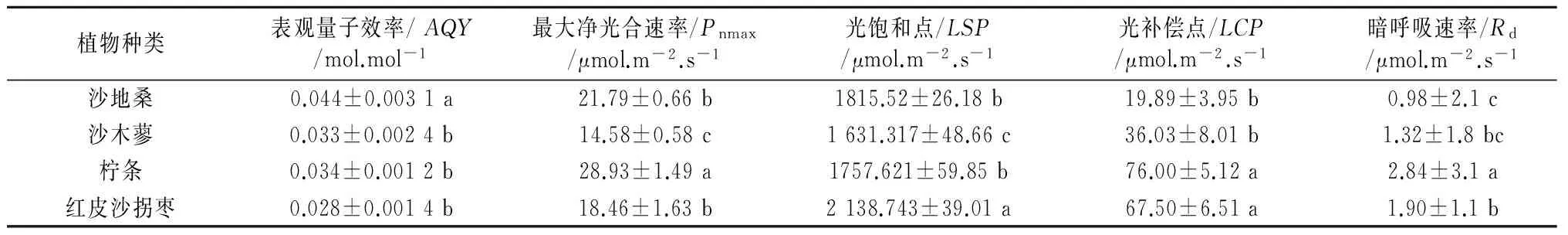
表2 4种植物光合-光强响应参数差异
3 讨论与结论
本研究中4种植物水势的日变化趋势与前人的研究结果相一致[8,9],基本表现出近“V”形变化,这一现象可能是因为,清晨水势高,植物体内并未出现生理缺水,新陈代谢及物质循环没有受到影响,随着气温、光强等环境因子的变化,植物体内更多的水分用来进行光合作用和蒸腾作用而根吸收的水分来不及补给[10],导致清晨至正午水势下降,正午过后,蒸汽压亏缺解除使得水势有所回升。正常水分条件下,柠条一天中的水势在4种植物种最低,说明它具有较强的吸水能力,相对来讲它对于干旱的耐受力也更强[11]。4种植物在相同的生长环境下出现不同的体内水分特征,这可能与植物的遗传特性和长期对生境适应的对策有关[12]。叶绿素是光合作用的重要参与物质,它在光合作用过程中负责对光能进行吸收和转化[13],植物叶片叶绿素含量的多少是影响植物光合作用能力的重要因素,4种植物中,柠条叶片的叶绿素含量最高,说明它在同等条件下能够更好地利用光能。
最大净光合速率可以反映植物在特定环境下的最大光合能力,本研究中4种植物Pnmax在14.58~28.93 μmol·m-2·s-1之间,其中柠条的Pnmax最大,红皮沙拐枣的Pnmax最小,说明同等环境条件下,柠条能够更多地吸收CO2积累有机物。表观量子效率可以表征植物在低光强对光能的利用效率[14],通常情况下植物的表观量子效率都在0.03~0.07之间[15],4种植物中,沙地桑的表观量子效率最大,说明沙地桑较其他3种植物可以更好地利用弱光。LCP和LSP分别是衡量植物利用强弱光的重要指标[16,17],通过计算LCP和LSP,可以得出植物对光强地适应范围,本研究中,4种植物光强适应范围不同,沙地桑具有较低的LCP和较高LSP,沙木蓼LCP和LSP均较低,柠条具有较高的LCP和较低的LSP,红皮沙拐枣LCP和LSP均较高。由此可见,沙地桑能够更好适应低光强环境,沙木蓼和柠条的光强适应范围相对较窄,红皮沙拐枣对弱光的利用稍弱,但其对强光的适应性更强。Rd是植物在无光条件下的呼吸速率,在一定程度上可以反映植物叶片的生理活性,本研究中沙地桑Rd最低,在无光的条件下沙地桑消耗较少的光合产物以积累更多的物质,柠条具有较高的Rd,同时拥有较高的最大净光合速率,说明柠条叶片具有较强生理活性,光合积累物质和呼吸消耗物质的能力都很强。
植物对于外界自然环境的反应,是其对于自然环境长期适应的结果[18]。研究植物在不同光强下的光合作用过程,可以了解其对于外界环境的适应机制[19]。光响应曲线是对植物光合作用的一种描述,可以很好地反应光合作用的效率[20,21]。4种植物Gs、Tr、水分利用效率随光强的变化表现为一元二次函数变化,且R2值基本大于0.9,拟合程度都很高。当光强超过800 μmol·m-2·s-1后,在4种植物中,柠条Pn最高,这可能是因为在不同光强下,柠条的Gs较大,有利于吸收CO2,而其Tr随着光强的增加上升程度适中,这使得柠条在高光强的环境中仍然具有较高的水分利用效率,而水分利用效率高通常也被认为是抗旱性好的一种表现[22]。红皮沙拐枣在不同光强下的Gs和Tr均最低,其水分利用效率在低光强下低于其他植物,但在高光强条件下,迅速上升,红皮沙拐枣LSP较高,表明在强光下仍可很好地进行光合作用,其叶片的形态使得其Tr较低,这是该植物在长期强光高温环境下进化出的水分利用策略,这一结论与前人研究结果一致[23]。沙木蓼和沙地桑Gs随光强的变化较为相似,试验后模拟计算发现,这两种植物在强光下会出现光饱和现象,但Tr仍随着光强的增加上升,因而达到光饱和点后,水分损失会随光强增加逐渐增加,因而水分利用效率下降,这两种植物的水分利用方式表现为较低光强下充分利用水分进行物质积累,在高光强下水分利用效率下降,若在干旱区出现这种情况,植物必将因水分损失过多死亡,但在长期进化过程中,干旱区植物在正午高温高光强环境下会“午休”,避免损失过多的水分,这也与前人研究得出的沙木蓼和沙地桑在正午会出现“午休”现象的结果相符[24]。环境条件一定时,植物的最大净光合速率显示了其光合作用的能力[25],光合-光响应曲线显示,4种植物中,柠条在强光下的净光合速率最大,同时柠条具有较高水分利用效率,它是判断植物对于环境适应性的重要指标[26],同时,前人研究结果认为,高Pn及高水分利用效率是植物抗旱性良好的表现[27],这也说明柠条在4种植物中抗旱性最好。
[1] 屠志方,李梦先,孙涛.第五次全国荒漠化和沙化监测结果及分析[J].林业资源管理,2016(1):1-5
[2] 尹立河,黄金廷,王晓勇,等.毛乌素沙地4种植物叶水势变化及其影响因素分析[J].植物资源与环境学报,2016,25(1):17-23
[3] 周瑞莲,侯玉平,左进城,等.不同沙地共有种沙生植物对环境的生理适应机理[J].生态学报,2015,35(2):340-349
[4] 谭永芹,柏新富,朱建军,等.干旱区五种木本植物枝叶水分状况与其抗旱性能[J].生态学报,2011,31(22):6815-6823
[5] 赵哈林,何玉惠,岳广阳,等.风吹、沙埋对沙地植物幼苗生长和光合蒸腾特性的影响[J].生态学杂志,2010,29(3):413-419
[6] Siam A M J, Radoglou K M, Noitsakis B, et al. Differences in ecophysiological responses to summer drought between seedlings of three deciduous oak species[J]. Forest Ecology & Management, 2009, 258(1):35-42
[7] 叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报,2010,34(6):727-740
[9] Singh K P, Malik R S, Malik D S. Diurnal variations in leaf water potential and stomatal conductance of pigeon pea 〔Cajanuscajan(L.) Millsp.〕 cultivars as affected by irrigation levels[J]. Biologia Plantarum,1983,25(1):1-4
[10] Nobel P S. Patricia Brewer Biophysical Plant Physiology and Ecology[C]. Freeman W H Company,1983.96,507
[11] 李向义,张希明,何兴元,等.绿洲前沿地区多枝柽柳水分关系的特征及灌溉的影响[J].植物生态学报,2004,28(5):644-650
[12] 付爱红,陈亚宁,李卫红,等.干旱、盐胁迫下的植物水势研究与进展[J].中国沙漠,2005,25(5):744-749
[13] 霍红,张勇,陈年来,等.干旱胁迫下五种荒漠灌木苗期的生理响应和抗旱评价[J].干旱区资源与环境,2011,25(1):185-189
[14] Richardson A D, Berlyn G P. Spectral reflectance and photosynthetic properties ofBetulapapyrifera(Betulaceae) leaves along an elevational gradient on Mt. Mansfield, Vermont, USA.[J]. American Journal of Botany, 2002, 89(1):88-94
[15] 丁俊祥,邹杰,唐立松,等.克里雅河流域荒漠-绿洲交错带3种不同生活型植物的光合特性[J].生态学报,2015,35(3):733-741
[16] 张淑勇,周泽福,夏江宝,等.不同土壤水分条件下小叶扶芳藤叶片光合作用对光的响应[J].西北植物学报,2007,27(12):2514-2521
[17] 伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响[J].林业科学,2007,43(5):30-35
[18] Zhao Z G, Du G Z, Zhou X H, et al. Variations with altitude in reproductive traits and resource allocation of three Tibetan species of Ranunculaceae.[J]. Australian Journal of Botany, 2006, 54(7):691-700
[19] Wang H, Wang F, Wang G, et al. The responses of photosynthetic capacity, chlorophyll fluorescence and chlorophyll content of nectarine (Prunuspersica, var. Nectarina, Maxim) to greenhouse and field grown conditions[J]. Scientia Horticulturae, 2007, 112(1):66-72
[20] Ye Z P. A new model for relationship between irradiance and the rate of photosynthesis inOryzasativa[J]. Photosynthetica, 2007, 45(4):637-640
[21] 郎莹,张光灿,张征坤,等.不同土壤水分下山杏光合作用光响应过程及其模拟[J].生态学报,2011,31(16):1517-1524
[22] Medrano H, Flexas J, Galmes J. Variability in water use efficiency at the leaf level among Mediterranean plants with different growth forms[J]. Plant and Soil, 2009, 317, 17-29
[23] 朱军涛,李向义,张希明,等.昆仑山北坡不同海拔塔里木沙拐枣的光合生理生态特性[J].生态学报,2011,31(3):611-619
[24] 罗青红,吉小敏,宁虎森,等.灌溉条件下3种沙生灌木光合特性及其与生态因子的关系[J].西北植物学报,2014,34(11):2296-2302
[25] Tartachnyk I I, Blanke M M. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remobilization of apple leaves[J]. New Phytologist, 2004, 164(3):441-450
[26] Sobrado M A. Relation of water transport to leaf gas exchange properties in three mangrove species[J]. Trees, 2000, 14(5):258-262
[27] Chierenbeck K A, Marshall J D. Seasonal and diurnal patterns of photosynthetic gas exchange for Lonicera sempervirens and L. japonica (Caprifoliaceae).[J]. American Journal of Botany, 1993, 80(11):1292-1299
猜你喜欢
干旱地区农业研究(2021年6期)2021-12-08
现代农村科技(2021年5期)2021-12-07
中国煤炭工业(2020年1期)2020-12-28
农村实用技术(2020年9期)2020-11-19
当代陕西(2019年7期)2019-04-25
红蜻蜓·低年级(2017年6期)2017-10-30
植物资源与环境学报(2016年1期)2016-08-12
现代农村科技(2016年6期)2016-03-28
节水灌溉(2016年3期)2016-03-23
中国蔬菜(2015年9期)2015-12-21
