
黄淮麦区小麦主栽品种粒重与籽粒灌浆特性的关系
2018-03-01 08:45苗永杰赵德辉田宇兵闫俊良夏先春张何中虎中国农业科学院作物科学研究所国家小麦改良中心北京0008中国农业科学院棉花研究所河南安阳455000国际玉米小麦改良中心CIMMYT中国办事处北京0008
作物学报 2018年2期
苗永杰 阎 俊 赵德辉 田宇兵 闫俊良 夏先春张 勇, 何中虎,中国农业科学院作物科学研究所 / 国家小麦改良中心, 北京 0008; 中国农业科学院棉花研究所, 河南安阳 455000; 国际玉米小麦改良中心(CIMMYT)中国办事处, 北京 0008
黄淮麦区是我国最重要的小麦产区, 播种面积和总产分别约占全国的 55%和 60%, 对保障粮食安全至关重要[1]。过去60余年, 黄淮麦区小麦产量年遗传增益约为 0.48%~1.05%, 其构成因素中千粒重年遗传增益较大, 为0.35%~0.51%[2]。虽然近15年产量遗传增益放缓, 但粒重仍持续提高[3], 表明粒重改良是该麦区产量显著提高的关键因素。粒重主要受基因型控制, 并受环境显著影响, 粒重在产量构成因素中的遗传力最高[4-5]。已定位了大量粒重相关QTL, 位于2A、4D、5B、6B、7B和7D染色体上的 QTL效应较大, 单个位点可解释 4.8%~28.0%的表型变异[6-8]。其中, TaCwi[9]、TaGW2[10]、TaSus2[11]、TaCKX6[12]、TaSAPl[13]、TaGS1a[14]、TaGS-D1[15]、TaGASR7-A1[16]、TaGS5-3A[17]、6-SFT-A2[18]等多个相关基因已被克隆, 可解释 4.8%~14.6%的表型变异。
粒重由籽粒灌浆速率和持续时间决定[19-23]。灌浆速率主要受基因型控制, 灌浆持续时间主要由特定地区的气候和耕作栽培制度决定[23-26]。有关灌浆速率和持续时间对粒重的贡献尚无定论, 多数研究认为, 在灌浆期偏短的地区, 灌浆速率对粒重的贡献大于持续时间[26-30]。因此, 育种工作的重点是提高灌浆速率。
灌浆速率是决定我国北部冬麦区小麦品种粒重最重要的参数[29]; 黄淮麦区水地和旱地品种籽粒灌浆特性存在显著差异, 旱地品种的灌浆速率高于水地品种, 但其籽粒灌浆时间较短[30]。这些研究多集中于描述具体品种的灌浆特性, 对不同粒重类型品种间的籽粒灌浆特性可能因为工作量大而缺乏系统研究, 且很少涉及黄淮麦区水地主栽品种的粒重与籽粒灌浆特性之间的关系。本研究通过分析品种间籽粒灌浆特性的差异, 明确粒重与籽粒灌浆特征参数的关系, 揭示灌浆速率和持续时间对粒重的相对重要性, 旨在为黄淮麦区小麦品种的粒重改良提供理论依据。
1 材料与方法
1.1 品种及田间设计
选用黄淮麦区 14份水地主栽品种和苗头品系(表 1), 于 2012—2015年度种植在河南安阳。良星99和周麦 18分别是黄淮北片和南片冬麦区国家水地组区域试验的对照品种。济麦22和矮抗58是黄淮麦区近10年累计推广面积最大的两个主栽品种。郑麦366、周麦16、良星66、周麦27、存麦1号、中麦 895和中麦 875均为当前生产上的主推品种,中麦871和中麦140是本课题组新育成的苗头品系。荔垦4号是中麦895、中麦875和中麦871的父本。
采用随机区组设计, 2次重复。小区面积6 m2, 6行区, 4 m行长, 行距20 cm。试验地前茬玉米, 收获后秸秆还田, 播种前底施复合肥(N、P2O5、K2O 比例17∶1∶17) 750 kg hm–1和尿素75 kg hm–1, 深翻。10月10日前后播种, 每公顷基本苗210万株。分别于越冬期、返青期、孕穗期和灌浆期灌溉 4次, 并结合越冬水和孕穗水分别追施尿素112.5 kg hm–1和150 kg hm–1。返青期化学除草一次, 抽穗扬花期“一喷三防”, 其他管理措施同当地大田生产。
1.2 田间调查指标及方法
开花至成熟期间的天数即为灌浆持续时间。于开花初期从每个小区选取同一天开花, 穗型、长势和大小基本一致, 无病虫害的单茎200个挂牌标记。从花后第6天开始, 每隔6 d取样一次, 直至成熟收获。每次各小区取15穗, 105°C杀青15 min, 65°C烘干24 h, 调查粒数, 并用千分之一天平称重, 计算千粒重。
以花后相对灌浆时间(x)为自变量, 粒重(y)为因变量, 采用Logistic方程y = k/(1+ae–bx)对不同品种的籽粒灌浆进程进行拟合, 绘制籽粒灌浆速率曲线,其中a和b为品种参数, k为理论粒重。
采用一阶和二阶求导, 计算最大灌浆速率(Rmax,mg grain–1d–1)及其到达时间(Tmax, d)、渐增期持续时间(T1, d)和灌浆速率(R1, mg grain–1d–1)及其增重(W1,g)、快增期持续时间(T2, d)和灌浆速率(R2, mg grain–1d–1)及其增重(W2, g)、缓增期持续时间(T3, d)和灌浆速率(R3, mg grain–1d–1)及其增重(W3, g)、平均灌浆持续时间(T, d)和灌浆速率(Ra, mg grain–1d–1)等参数。气压、温度、湿度、降水量、风速和日照等主要气象资料来自中国气象数据网(http://cdc.cma.gov.cn/)河南安阳站“中国地面气候资料日值数据集”。
1.3 统计分析
采用Statistical Analysis System (SAS9.2)软件,调用PROC NLIN程序进行灌浆数据的Logistic方程拟合。调用PROC CLUSTER程序, 按Ward最小平方和法, 依据品种粒重进行聚类分析。调用 PROC GLM 模型, 粒重类型为固定效应, 类内品种、年份内重复及其相关互作为随机效应, 进行方差分析。调用PROC CORR程序进行相关分析。
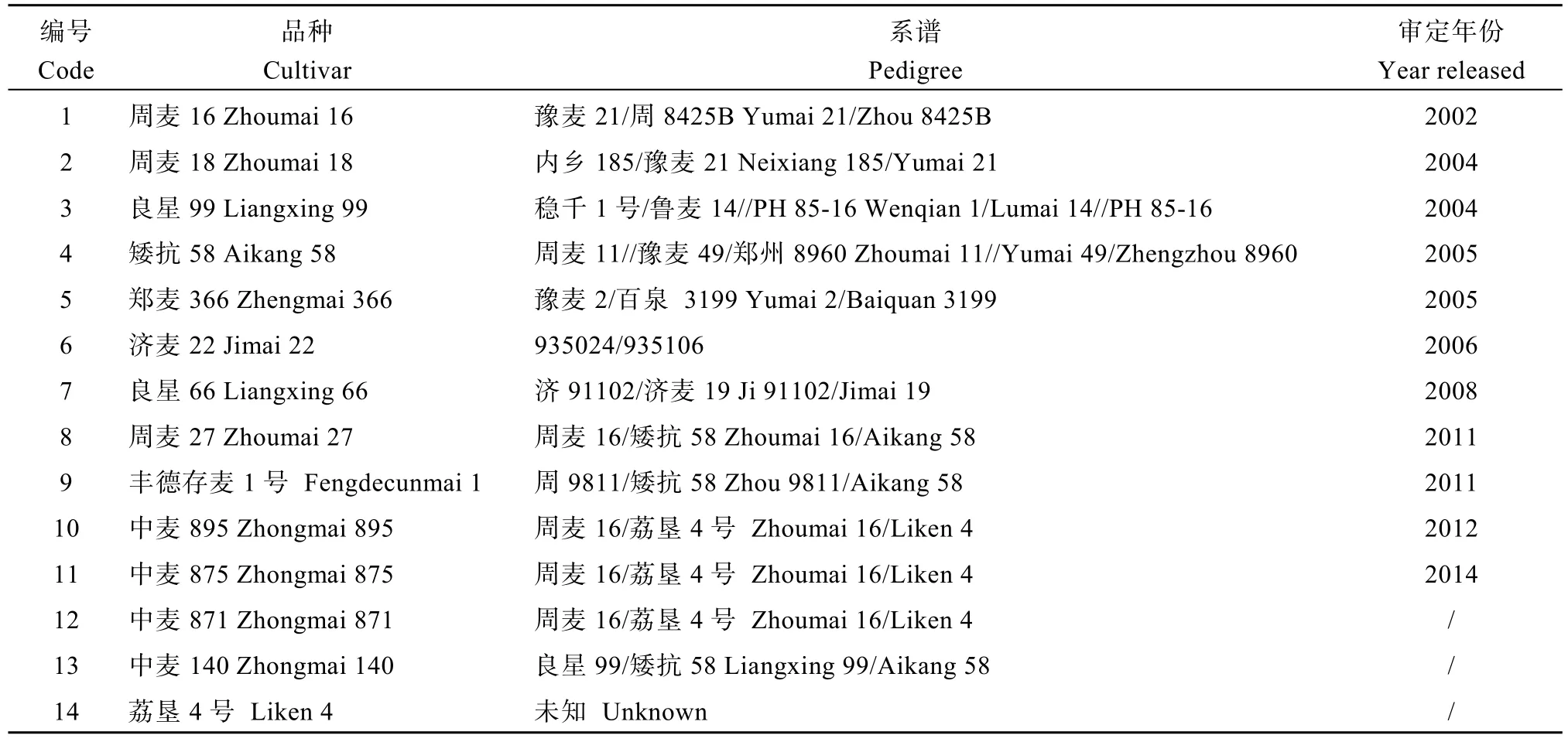
表1 14份参试品种名称、系谱及其审定年份Table 1 Names, pedigrees and released years of the 14 cultivars investigated
2 结果与分析
2.1 籽粒灌浆特征参数基本统计量分析
千粒重及所有灌浆特征参数的变异范围均较大(表2)。参试品种的千粒重介于41.8~53.8 g之间, 平均和最大灌浆速率分别为 1.15~1.50 mg grain–1d–1和1.94~2.63 mg grain–1d–1, 渐增期、快增期和缓增期灌浆速率分别为 0.78~1.02、1.70~2.31和 0.69~1.10 mg grain–1d–1, 相应时期的籽粒增重量分别为9.2~12.1、25.2~33.3 和 9.2~12.1 mg grain–1, 上述参数的品种间变异范围均大于年份。品种间灌浆持续时间、最大灌浆速率到达时间和渐增期持续时间的变异范围分别为 36.3~40.3、18.0~19.8和 11.3~12.4 d,品种间变异范围均小于年份。品种间快增期和缓增期持续时间的变异范围分别为 12.7~15.5 d和11.1~13.9 d, 品种间变异范围与年份相当。
2.2 籽粒灌浆特征参数方差分析
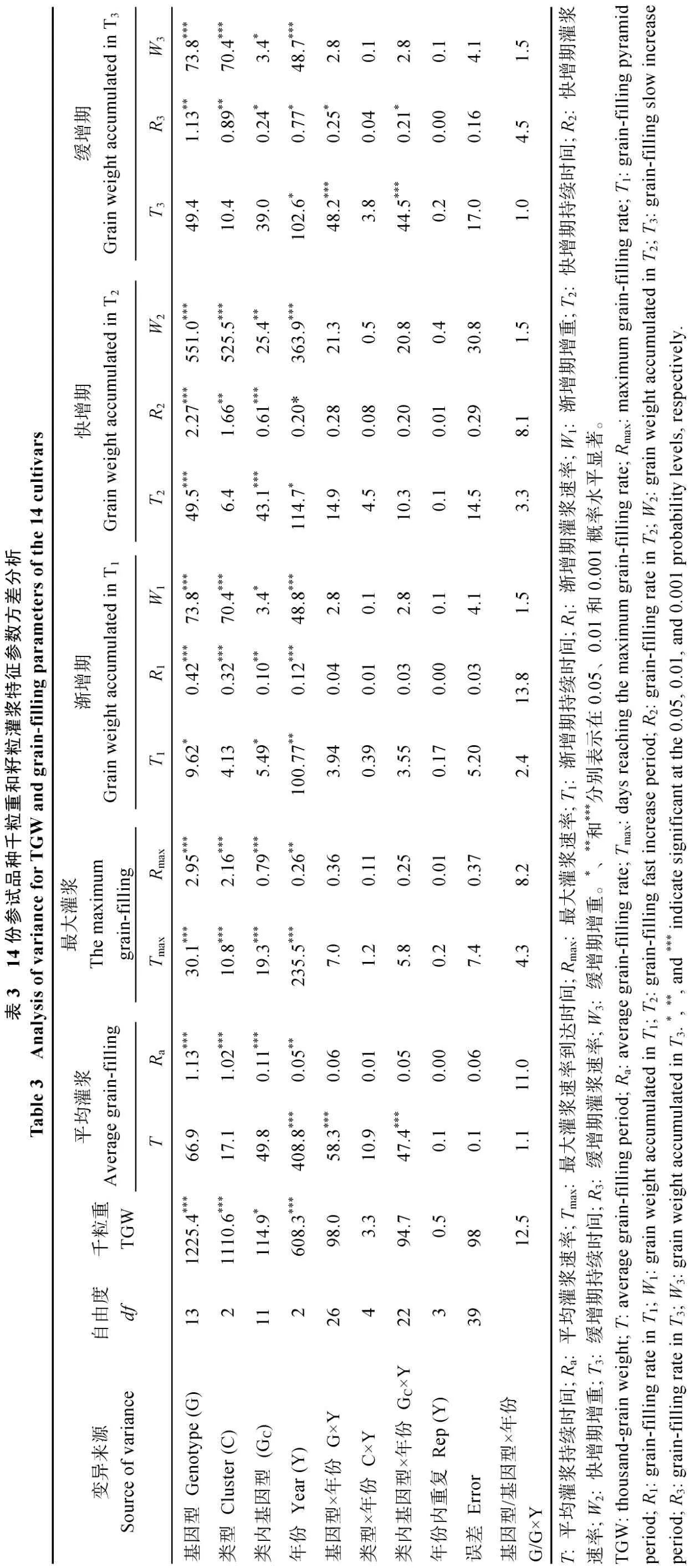
千粒重、所有籽粒灌浆速率及各时期籽粒增重量参数均受基因型和年份效应的显著影响, 灌浆持续时间和缓增期持续时间的年份和基因型×年份效应、渐增期和快增期持续时间的基因型和年份效应显著(表3)。千粒重、所有籽粒灌浆速率及各时期增重量的基因型效应较大, 其次为年份效应, 且基因型效应远大于基因型×年份效应, 表明粒重、灌浆速率和增重量各参数主要受基因型控制。所有灌浆持续时间的年份效应较大, 其次为基因型效应, 且基因型效应均大于基因型×年份效应, 表明灌浆持续时间主要受环境影响。
按千粒重将14个品种聚为3类, 高粒重品种包括周麦 18、中麦 895和中麦 875, 中等粒重品种包括周麦16、良星99、济麦22、存麦1号和中麦140,低粒重品种包括矮抗58、良星66、郑麦366、周麦27、中麦871和荔垦4号。千粒重、最大灌浆速率到达时间、所有籽粒灌浆速率及其增重量参数均受粒重类型及类内基因型效应的显著影响, 渐增期和快增期持续时间的类内基因型效应显著, 灌浆持续时间、缓增期持续时间及其灌浆速率的类内基因型×年份效应显著。粒重的类型效应远大于类内基因型效应, 类型×年份及类内基因型×年份效应不显著。所有灌浆速率参数及各时期籽粒增重量的类型效应均大于类内基因型效应, 灌浆持续时间相关参数的类型效应不显著, 表明不同粒重类型品种间的灌浆速率及各时期籽粒增重量均存在显著差异, 灌浆持续时间则差异不显著。
2.3 粒重类型间籽粒灌浆特征参数分析
所有品种的籽粒饱满度均较好, 表明灌浆正常。Logistic方程决定系数(R2)均在0.99以上, 说明拟合方程可以有效描述籽粒灌浆进程。不同粒重类型品种灌浆进程均呈“S”型变化曲线, 灌浆速率变化趋势呈单峰曲线, 表现为慢—快—慢的特征(图 1),不同粒重类型品种间平均和最大灌浆速率及渐增期、快增期和缓增期灌浆速率及其籽粒增重量均存在显著差异, 表现为高粒重>中等粒重>低粒重, 灌浆持续时间则差异不显著(表4)。渐增期、快增期和缓增期持续时间分别约占整个灌浆期的 31.2%、36.9%和 31.9%, 其籽粒增重量则分别约占粒重的21.1%、57.8%和21.1%。与渐增期和缓增期相比, 快增期高、中、低粒重类型品种间灌浆速率和籽粒增重量差异较大。高粒重和中等粒重类型品种的最大灌浆速率到达时间显著晚于低粒重类型品种, 其他灌浆持续时间相关参数类型间差异均不显著。
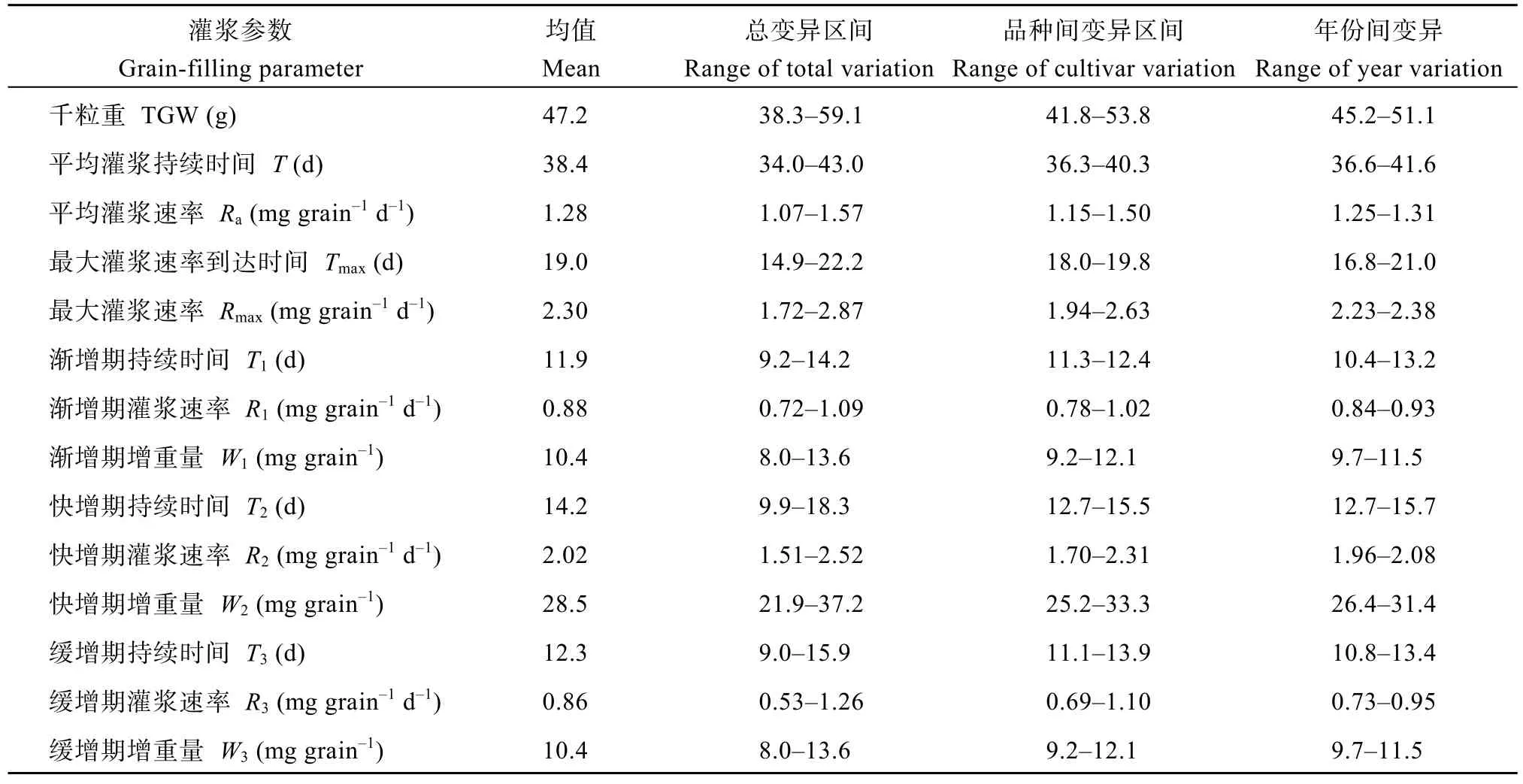
表2 14份参试品种千粒重和籽粒灌浆特征参数均值及其变异范围Table 2 Mean and variation of thousand-grain weight (TGW) and grain-filling parameters of the 14 cultivars investigated
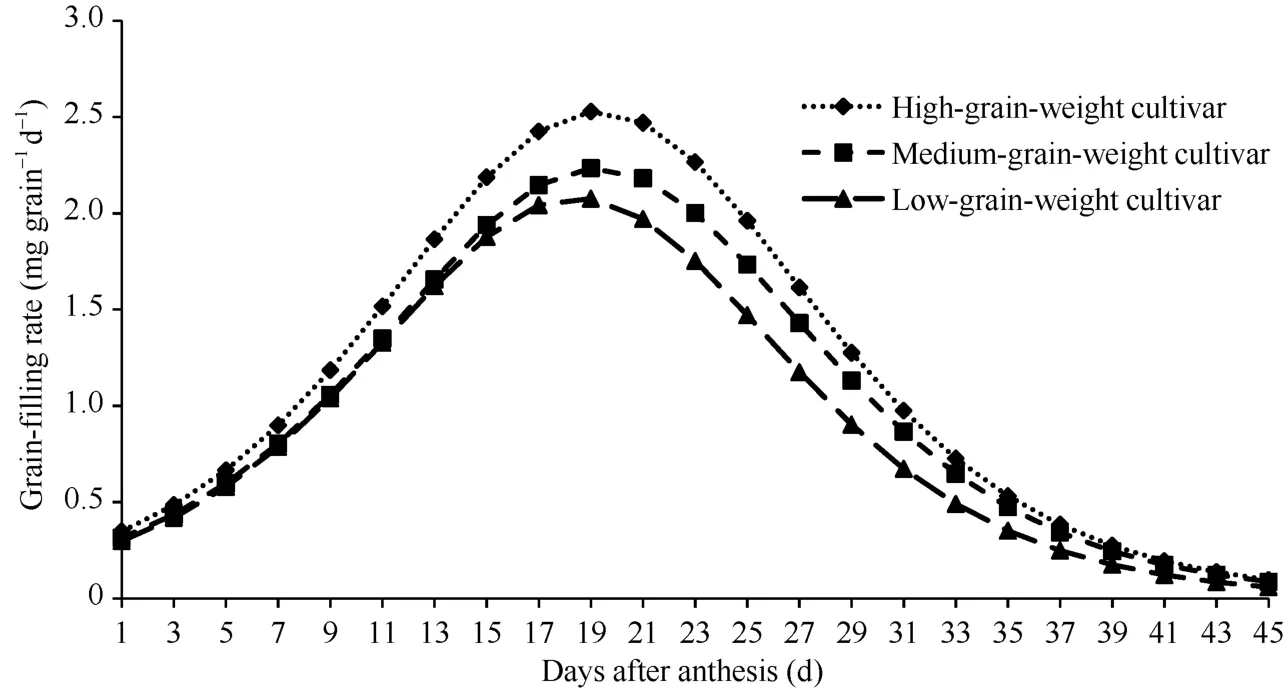
图1 不同粒重类型小麦品种的籽粒灌浆速率曲线Fig. 1 Grain-filling rate curve of different grain-weight wheat cultivars
2.4 千粒重与籽粒灌浆特征参数的相关性
千粒重与各时期籽粒灌浆速率均呈显著正相关(P < 0.001), 其中与快增期灌浆速率的相关程度最密切(r = 0.97), 与平均灌浆速率的相关程度次之(r= 0.90) (图2和表5)。千粒重与各时期籽粒增重量均呈显著正相关(r = 0.97, P < 0.001), 与灌浆持续时间参数相关均不显著(表5)。
?
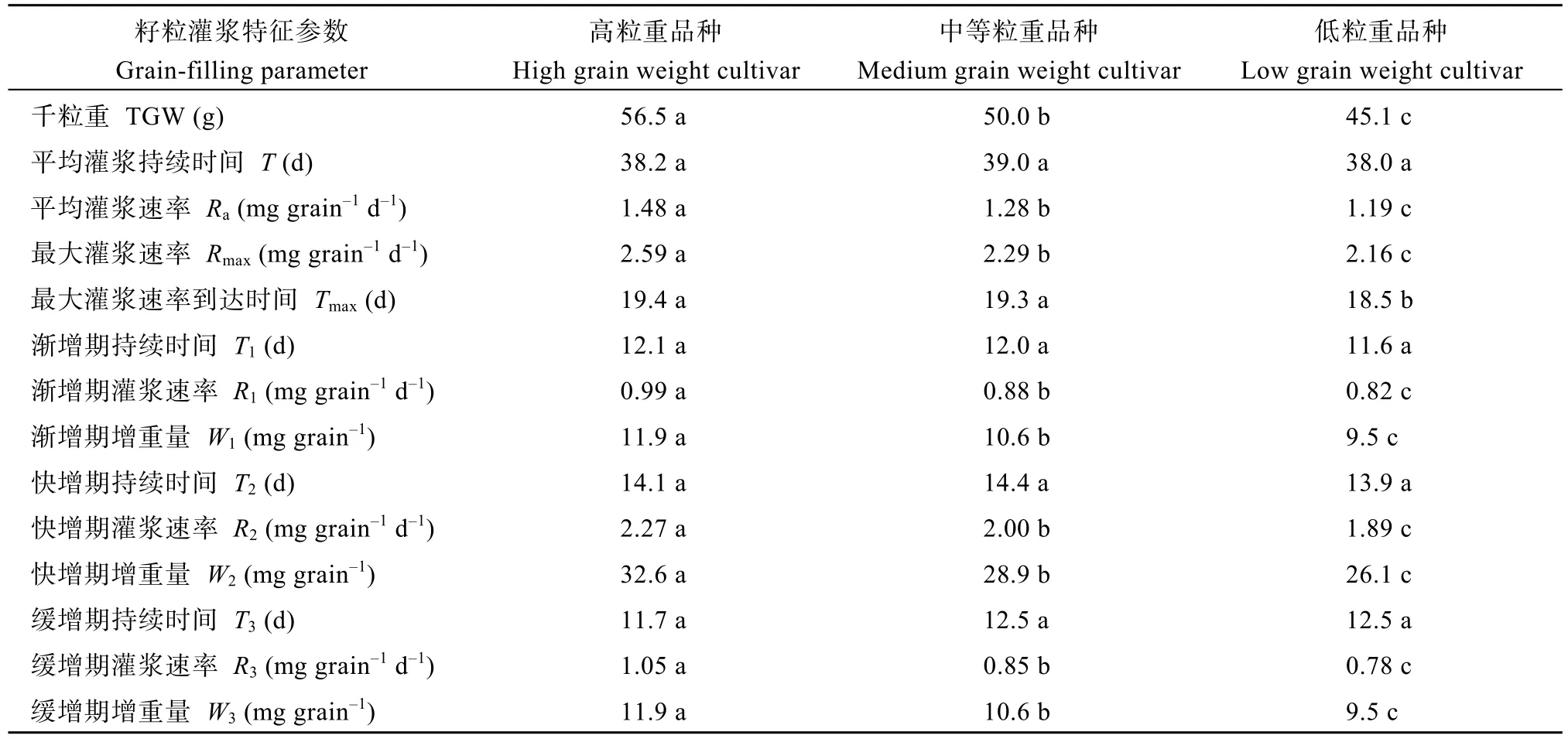
表4 不同粒重类型品种的籽粒灌浆特征参数Table 4 Grain-filling parameters in different grain-weight cultivars
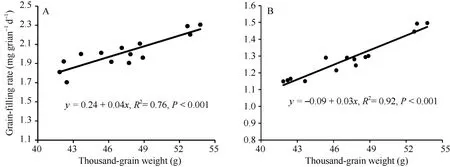
图2 千粒重与灌浆速率的线性回归Fig. 2 Linear regressions between thousand-grain weight and grain-filling rate
3 讨论
黄淮冬麦区小麦灌浆期经常遇到高温、干旱和干热风的影响, 使籽粒不饱满, 粒重下降, 造成产量损失。大量研究表明, 粒重主要取决于灌浆速率,而灌浆持续时间对粒重的贡献相对较小[6,8,12,20-21,27]。本试验结果与此一致, 黄淮麦区小麦品种的千粒重与各时期籽粒灌浆速率均呈显著正相关, 其中与快增期灌浆速率相关最密切, 且千粒重与各时期增重量均呈显著正相关, 与灌浆持续时间各参数相关不显著。渐增期、快增期和缓增期的持续时间分别约占灌浆期 31.2%、36.9%和 31.9%, 籽粒增重量则分别约占粒重的 21.1%、57.8%和 21.1%, 进一步表明快增期籽粒增重量对粒重影响较大。由此可见, 灌浆速率, 特别是快增期灌浆速率, 对粒重的形成具有重要贡献。
粒重与平均灌浆速率呈显著正相关(r = 0.90, P< 0.001)。由于灌浆速率测定繁琐, 利用分离群体对该性状进行基因定位难度很大。鉴于灌浆速率对粒重的重要性, 可以根据粒重与灌浆持续时间计算平均灌浆速率, 仅需记录开花期、成熟期和粒重, 对相关性状进行基因定位, 以进一步解析粒重的遗传机制。王瑞霞等[31]利用平均灌浆速率将粒重相关QTL定位于 1B、2A和 3B染色体, 可分别解释 5.8%~20.8%的表型变异。因此, 进一步明确 TaCwi[9]、TaGW2[10]等已克隆的粒重基因与籽粒灌浆速率的关系, 将有助于这些基因在育种中的有效利用, 进一步改良黄淮麦区小麦品种的籽粒灌浆特性。
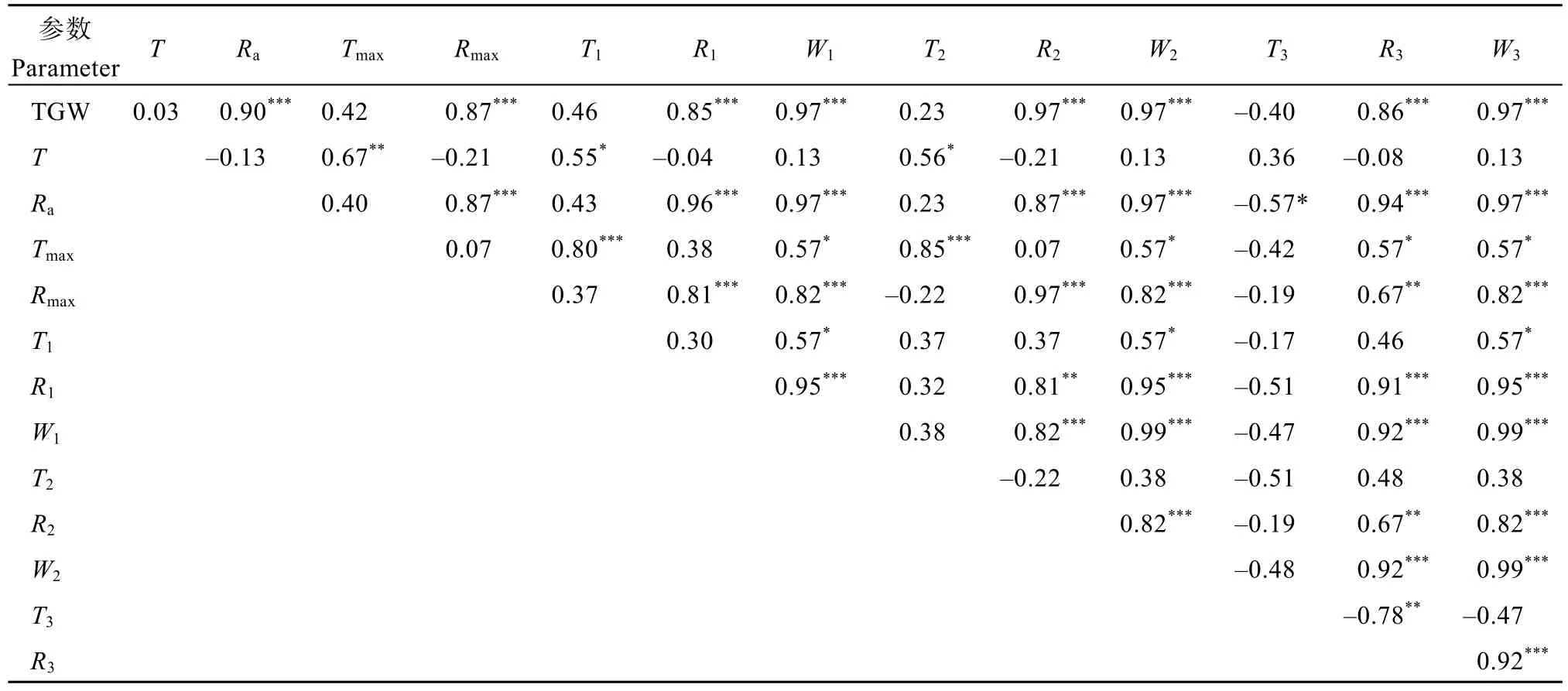
表5 千粒重与籽粒灌浆特征参数的相关系数Table 5 Correlation coefficients between thousand-grain weight and grain-filling parameters
本研究还表明, 来自同一组合的 3个品种中麦895、中麦875和中麦871在粒重和籽粒灌浆速率方面存在显著差异, 中麦 895和中麦 875的粒重和各时期灌浆速率均显著高于其姊妹系中麦 871及其双亲周麦16和荔垦4号。周麦16籽粒较大, 但灌浆速率偏慢, 常年饱满度一般, 粒重并不高, 约 46 g;荔垦 4号成熟落黄好, 籽粒饱满。我们当时组配周麦16和荔垦4号组合是期望通过提高周麦16的灌浆速率来改良其粒重, 以进一步提高产量。根据我们多年多点观察结果, 中麦 895和中麦 875的籽粒饱满度好, 灌浆速率快, 粒重高且稳定, 这说明在进行粒重改良时, 杂交组合亲本选配和后代选择至关重要, 后代选择应主要考察品种的田间落黄特性及其收获后籽粒的大小和饱满度。我们正在对中麦895的快速灌浆和高粒重特性进行研究, 期望发掘相关基因和分子标记, 以辅助改良粒重。
4 结论
黄淮麦区小麦粒重和灌浆速率各参数主要受基因型控制, 灌浆持续时间主要受环境影响。不同粒重类型品种间灌浆速率存在显著差异, 表现为高粒重>中等粒重>低粒重。灌浆速率, 特别是快增期灌浆速率的差异是导致品种间粒重高低的主要因素。
[1]茹振钢, 冯素伟, 李淦. 黄淮麦区小麦品种的高产潜力与实现途径. 中国农业科学, 2015, 48: 3388–3393 Ru Z G, Feng S W, Li G. High yield potential and effective ways of wheat in Yellow and Huai River Valley Facultative Winter Wheat Region. Sci Agric Sin, 2015, 48: 3388–3393 (in Chinese with English abstract)
[2]Zhou Y, He Z H, Sui X X, Xia X C, Zhang X K, Zhang G S. Genetic improvement of grain yield and associated traits in the Northern China Winter Wheat Region from 1960 to 2000. Crop Sci, 2007, 47: 245–253
[3]Gao F M, Ma D Y, Yin G H, Rashieed A, Dong Y, Xiao Y G, Xia X C, Wu X X, He Z H. Genetic progress in grain yield and physiological traits in Chinese wheat cultivars of southern Yellow and Huai Valley Winter Wheat Zone since 1950. Crop Sci, 2017,57: 760–773
[4]肖世和, 何中虎. 小麦产量潜力和品质的改良. 见: 庄巧生.中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003.pp 497–542 Xiao S H, He Z H. Wheat yield and end use quality improvement in China. In: Zhuang Q S, ed. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003. pp 497–542 (in Chinese)
[5]Wang Y Q, Hao C Y, Zheng J, Ge H M, Zhou Y, Ma Z Q, Zhang X Y. A haplotype block associated with thousand kernel weight on chromosome 5DS in common wheat (Triticum aestivum L.). J Integr Plant Biol, 2015, 57: 662–672
[6]Huang X Q, Coster H, M. Ganal W, Röder M S. Advanced backcross QTL analysis for the identification of quantitative trait loci alleles from wild relatives of wheat (Triticum aestivum L.). Theor Appl Genet, 2003, 106: 1379–1389
[7]Elouafi I, Nachit M M. A genetic linkage map of the Durum×Triticum dicoccoides backcross population based on SSRs and AFLP markers, and QTL analysis for milling traits. Theor Appl Genet, 2004, 108: 401–413
[8]Quarrie S A, Steed A, Calestani C, Semikhodskii A, Lebreton C,Chinoy C, Steele N, Pljevljakusić D, Waterman E. A high-density genetic map of hexaploid wheat (Triticum aestivum L.) from the cross Chinese Spring × SQ1 and its use to compare QTLs for grain yield across a range of environments. Theor Appl Genet,2005, 110: 865–880
[9]Ma D Y, Yan J, He Z H, Wu L, Xia X C. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers. Mol Breed,2010, 29: 43–52
[10]Su Z Q, Hao C Y, Wang L F, Dong Y C, Zhang X Y. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat (Triticum aestivum L.).Theor Appl Genet, 2011, 122: 211–223
[11]Jiang Q Y, Hou J, Hao C Y, Wang L F, Ge H M, Dong Y S.Zhang X Y. The wheat (T. aestivum) sucrose synthase 2 gene(TaSus2) active in endosperm development is associated with yield traits. Funct Integr Genomics, 2011, 11: 49-61
[12]Zhang L, Zhao Y L, Gao L F, Zhao G Y, Zhou R H, Zhang B S,Jia J Z. TaCKX6-D1, the ortholog of rice OsCKXl, is associated with grain weight in hexaploid wheat. New Phytol, 2012, 195:574–584
[13]Chang J Z, Zhang J N, Mao X G, Li A, Jia J Z, Jing R L. Polymorphism of TaSAPl-Al and its association with agronomic traits in wheat. Planta, 2013, 237: 1495–1508
[14]Guo Y, Sun J, Zhang G, Wang Y, Kong F, Zhao Y, Li S. Haplotype, molecular marker and phenotype effects associated with mineral nutrient and grain size traits of TaGS1a in wheat. Field Crops Res, 2013, 154: 119–125
[15]Zhang Y J, Liu J D, Xia X C, He Z H. TaGS-D1, an ortholog of rice OsGS3, is associated with grain weight and grain length in common wheat. Mol Breed, 2014, 34: 1097–1107
[16]Dong L, Wang F, Liu T, Dong Z, Li A, Jing R, Mao L, Li Y, Liu X, Zhang K, Wang D. Natural variation of TaGASR7-A1 affects grain length in common wheat under multiple cultivation conditions. Mol Breed, 2014, 34: 937–947
[17]Ma L, Li T, Hao C, Wang Y, Chen X, Zhang X. TaGS5-3A, a grain size gene selected during wheat improvement for larger kernel and yield. Plant Biotech J, 2016, 14: 1269–1280
[18]Yue A Q, Li A, Mao X G, Chang X P, Li R Z, Jing R L. Identification and development of a functional marker from 6-SFT-A2 associated with grain weight in wheat. Mol Breed, 2015, 35: 63
[19]Dngid S D, Brule-Babel A L. Rate and duration of grain filling in five spring wheat (Triticum aestivum L.) genotypes. Can J Plant Sci, 1994, 74: 681–686
[20]Saini H S, Westgate M E. Reproductive development in grain crops during drought. Adv Agron, 1999, 68: 59–96
[21]Zahedi M, Jenner C F. Analysis of effects in wheat of high temperature on grain filling attributes estimated from mathematical models of grain filling. J Agric Sci, 2003, 141: 203–212
[22]Yang J C, Zhang J H. Grain filling of cereals under soil drying.New Phytol, 2006, 169: 223–236
[23]Kamaluddin, Singh R M, Abdin M Z, Khan M A, Alam T, Kham S, Prasad L C, Joshi A K. Inheritance of grain filling duration in spring wheat (Triticum aestivum L. em Thell). J Plant Biol, 2007,50: 504–507
[24]Wong L S L, Baker R J. Selection for time to maturity in spring wheat. Crop Sci, 1986, 26: 1171–1175
[25]Talbert L E, Lanning S P, Murphy R L, Martin J M. Grain fill duration in twelve hard red spring wheat crosses: genetic variation and association with other agronomic traits. Crop Sci, 2001,41: 1390–1395
[26]吴晓丽, 汤永禄, 李朝苏, 吴春, 黄钢, 马蓉. 四川盆地小麦籽粒的灌浆特性. 作物学报, 2014, 40: 337–345 Wu X L, Tang Y L, Li C S, Wu C, Huang G, Ma R. Characteristics of grain filling in wheat growing in Sichuan basin. Acta Agron Sin, 2014, 40: 337–345 (in Chinese with English abstract)
[27]Dias A S, Lidon F C. Evaluation of grain filling rate and duration in bread and durum wheat, under heat stress after anthesis. J Agron Crop Sci, 2009, 195: 137–147
[28]Motzo R, Giunta F, Pruneddu G. The response of rate and duration of grain filling to long-term selection for yield in Italian durum wheats. Crop Pasture Sci, 2010, 61: 162–169
[29]曾浙荣, 庞家智, 周桂英, 赵双宁, 曹梅林. 我国北部冬麦区小麦品种籽粒灌浆特性的研究. 作物学报, 1996, 22: 720–728 Zeng Z R, Pang J Z, Zhou G Y, Zhao S N, Cao M L. Grain filling properties of winter wheat varieties in northern part of China. Acta Agron Sin, 1996, 22: 720–728 (in Chinese with English abstract)
[30]吴少辉, 段国辉, 高海涛, 张学品, 温红霞, 余四平, 马飞. 黄淮麦区水、旱生态型小麦籽粒灌浆进程研究. 麦类作物学报,2009, 29: 1015–1021 Wu S H, Duan G H, Gao H T, Zhang X P, Wen H X, Yu S P, Ma F. Research on wheat grain filling process of water and dryland ecological types of wheat in Huang-Huai area. J Triticeae Crops,2009, 29: 1015–1021 (in Chinese with English abstract)
[31]王瑞霞, 张秀英, 伍玲, 王瑞, 海林, 闫长生, 游光霞, 肖世和.不同生态环境条件下小麦籽粒灌浆速率及千粒重 QTL分析.作物学报, 2008, 34: 1750–1756 Wang R X, Zhang X Y, Wu L, Wang R, Hai L, Yan C S, You G X, Xiao S H. QTL mapping for grain filling rate and thousand-grain weight in different ecological environments in wheat.Acta Agron Sin, 2008, 34: 1750–1756 (in Chinese with English abstract)
猜你喜欢
作物学报(2022年3期)2022-01-22
中国农业气象(2021年7期)2021-07-28
麦类作物学报(2021年3期)2021-05-25
劳动保护(2020年11期)2020-12-23
北京农学院学报(2020年1期)2020-03-09
热带农业工程(2019年5期)2019-05-19
中国种业(2019年8期)2019-01-05
中国种业(2019年8期)2019-01-05
种业导刊(2019年5期)2019-01-04
麦类作物学报(2018年10期)2018-12-12
