
鹅掌楸不同种源苗期生长特征及其变异规律分析
2018-02-28 02:50:19潘文婷厉月桥茹广欣夏良放
江西农业大学学报 2018年1期
夏 莘,潘文婷,厉月桥,茹广欣,刘 儒,夏良放*
(1.国家林业局泡桐研究开发中心,河南 郑州 450003;2.中国林业科学研究院 亚热带林业实验中心,江西 分宜 336600;3.河南农业大学,河南 郑州 450000)
鹅掌楸属(Liriodendron)隶属木兰科(Magnoliaceae),因叶子形如马褂,又名马褂木。鹅掌楸为落叶乔木,其生长迅速,树干通直,是很好的用材树种,并且由于其叶形美观、花期较长、抗病虫、抗逆性较强,因此,也是非常好的园林绿化树种[1-3]。鹅掌楸属植物在新生代有二十余种[4],但经过第四冰期后,大部分灭绝,现在全世界只有中国鹅掌楸(Liriodendronchinense)和北美鹅掌楸(Liriodendrontulipifera)两种[5-7]。且由于地理的变迁、人为因素及其本身的生物学特性、天然群体种子萌发率低等影响,导致其处于濒危状态[8-12]。如何为基地建设提供优良的鹅掌楸种源是林木育种学家最关心的问题。
目前,国内外在鹅掌楸人工林培育方面的研究,主要集中在杂交、扦插以及林分生长发育规律特征等方面[13-19]。董纯等[20]和李斌等[21]曾对鹅掌楸种源试验林进行了部分研究,但分析的性状都比较少,本文对中国鹅掌楸15个种源和北美鹅掌楸5个种源进行不同种源幼苗物候期、生长节律、10种生物量性状等的比较、分析,探索鹅掌楸地理种源苗期主要经济性状的地理变异规律,以期为适应本地栽培种源的选择提供理论依据。
1 材料与方法
1.1 试验地概况
试验地设于亚林中心(江西分宜)年珠林场,北纬27°34′41″,东经114°33′47″,海拔200 m,当年气象资料表明:年均温度16.8 ℃,最高气温39.9 ℃,最低气温-5.3 ℃,年均降水量1 910 mm,相对温度80%,无霜期256 d,属亚热带季风气候。
1.2 材料来源
由项目组提供收集保存的种源20个,其中:15个中国鹅掌楸种源来自全国7个省地覆盖全分布区,分别是: 四川叙永(XY)、四川酉阳(YY)、湖北鄂州(EZ)、安徽黄山(HS)、贵州陌南(MN)、贵州黎平(LP)、湖南浏阳(LY)、湖南桑植(SZ)、湖南绥宁(SN)、浙江富阳(FY)、浙江松阳(SY)、江西庐山(LS)、江西武夷山(WYS)、云南勐腊(YN)、大别山舒城(DBS)。5个北美鹅掌楸种源,分别为:美国密苏里(MSL)、路易斯安纳(LYS)、北卡罗来纳(BK)、南卡罗来纳(NK)、佐治亚(ZZY)。
1.3 育苗设计
采用随机区组,重复4次,共80个小区,每小区6~10 m2,每平方米苗不超过35株。用裸根育苗方法,苗地前作为水稻土,苗木出土后,采用相同的田间常规管理措施。
1.4 调查方法
分别对各种源的物侯期、生长节律(每隔15天观测1次)、病虫害、抗寒性进行调查观测,年终在重复中同抽20株测定苗高和地径。用标准株调查苗木的根系和生物量及节间长,共抽3个重复,每重复取5株,清洗根系后称鲜质量,调查侧根和主根数,分别地上和地下部分烘干后称量。
1.5 统计分析
根据调查数据,通过SAS软件和SPSS软件进行有关统计分析,然后依据分析结果对种源表现作出初步评定。
2 结果与分析
2.1 不同种源幼苗各性状特征及比较分析
2.1.1 不同种源幼苗各性状特征 经测定(表1),鹅掌楸各种源的苗高、地径、根系、鲜质量、生物量和节间长均有所差别。不同种源苗高变异幅度为40.33~83.64 cm,其中YN苗高最高,DBS苗高最低,二者间相差2.07倍,地径变异幅度为1.12~1.40 cm,其中FY地径最大,XY地径最小,前者为后者的1.25倍,方差分析表明:不同种源鹅掌楸幼苗在苗高和地径性状间存在明显差异(P<0.01);幼苗主根数量变幅在1.01~1.47根,主根数较多的种源有FY、BK、MSL和SN,侧根数量变幅在28.20~37.53根,侧根数量较多的种源有SN、SY、MN和HS。
在幼苗生物量方面,除叶量外,各种源全株鲜质量为33.60~70.97 g,生物量为10.73~25.62 g,其中BK、LYS和FY的生物量最高,而DBS、YY和XY的生物量最低。同时也可以看出,不同种源生物量在地上与地下的分配比例也有所差异,幼苗地上生物量与地下生物量分配比变动幅度为0.68~1.54,以SN和HS地上生物量与地下生物量比值最高,分别为1.54和1.48,总体来看北美鹅掌楸的地下生物量明显高于地上生物量,除BK外的4个种源,地上生物量仅为地下生物量的80%左右。
幼苗节间长与苗高呈正相关,各种源平均节间长变幅在1.96~3.49 cm,节间较长的种源有SN、YN、EX和LS。
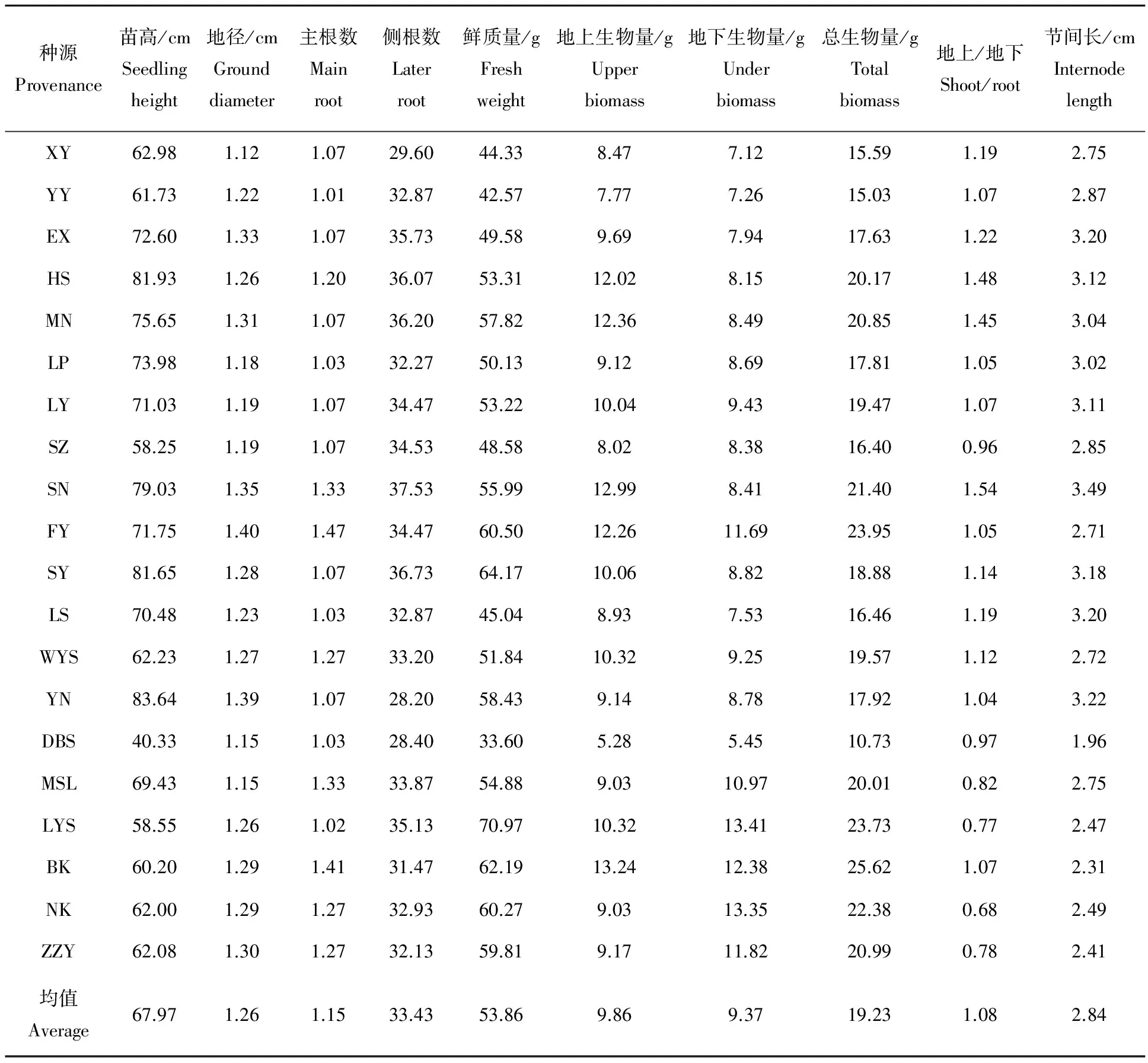
表1 鹅掌楸不同种源幼苗各性状特征
2.1.2 不同种源幼苗各性状间的相关关系 从表2可以看出,鹅掌楸幼苗诸多性状间存在显著或极显著正相关关系,如苗高与节间长存在极显著正相关关系,与侧根数和地上生物量呈显著正相关关系。而地径与鲜质量和地上生物量呈极显著正相关关系,而与总生物量呈显著正相关关系。说明鹅掌楸幼苗的许多主要性状间具有密切的相关关系。可以用容易测定的指标估计不易测定的指标,为鹅掌楸苗期选择提供基础。
从图1可以看出,鹅掌楸幼苗生物量与苗高、地径的最佳拟合曲线均为二次曲线关系,随着苗高和侧根数增加呈现先增加后下降的趋势,而随着地径和主根数的增加而增加的趋势,但相关显著性不大。
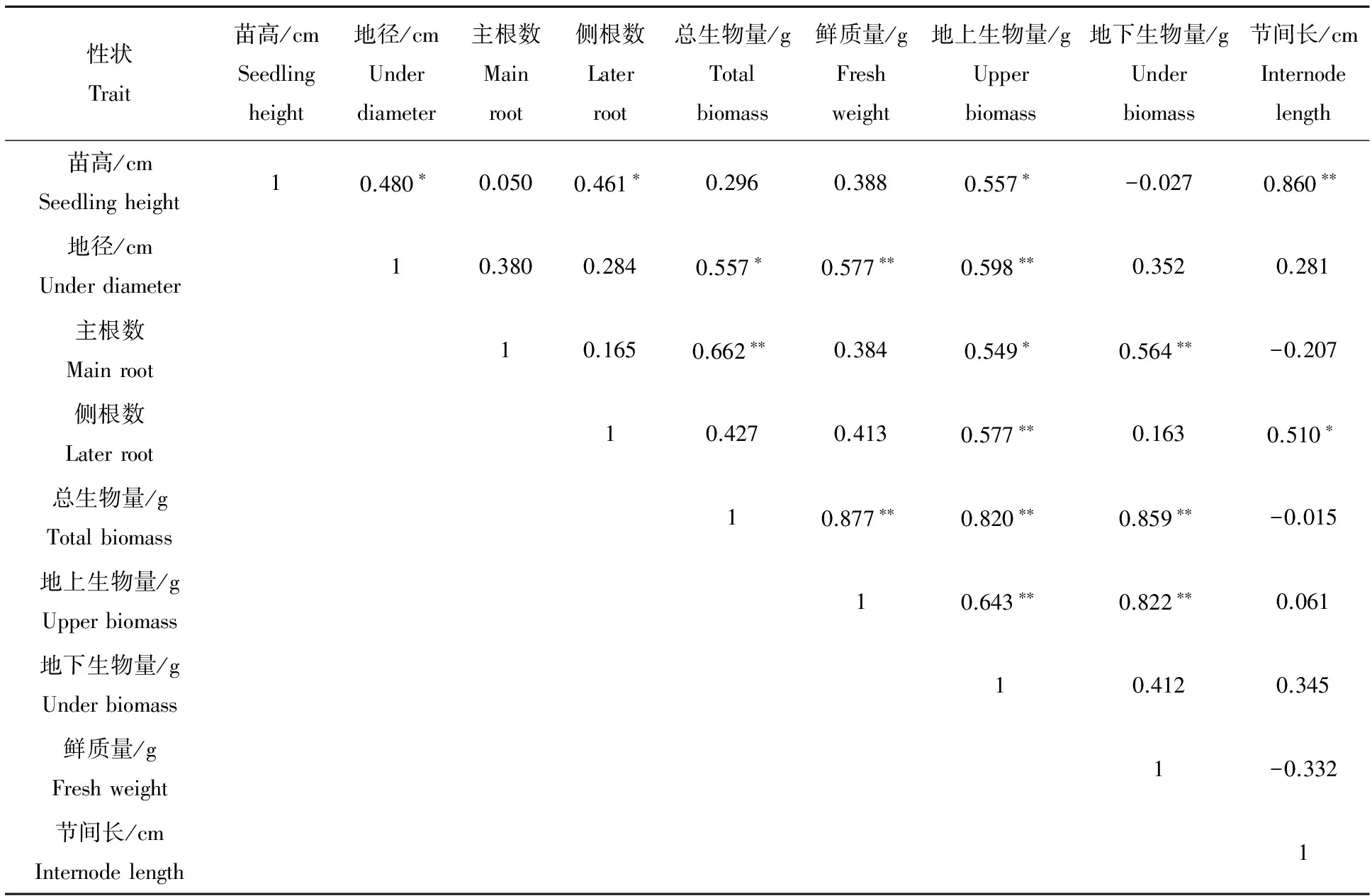
表2 鹅掌楸苗期主要性状间的相关关系
**表示差异极显著(P<0.01);*表示差异显著(P<0.05),下同。
**significant at 0.01 level;*significant at 0.05 level,the same below.

图1 鹅掌楸幼苗生物量与苗高、地径和根系的关系Fig.1 The relationship among seedlings biomass,height,diameter and root numbers of Liriodendron
2.1.3 中国鹅掌楸与北美鹅掌楸幼苗生长性状与生物量性状特征差异 中国鹅掌楸与北美鹅掌楸幼苗各性状指标,采用t检验法进行方差分析结果表明(表3),中国鹅掌楸苗高显著高于北美鹅掌楸,节间长与地上生物量/地下生物量均极显著高于北美鹅掌楸,而地下生物量与总生物量极显著低于北美鹅掌楸,据观测北美鹅掌楸尖削度明显大于中国鹅掌楸,而其它性状差异不显著。说明中国鹅掌楸与北美鹅掌楸幼苗个体发育构型上发生了显著性的变化。
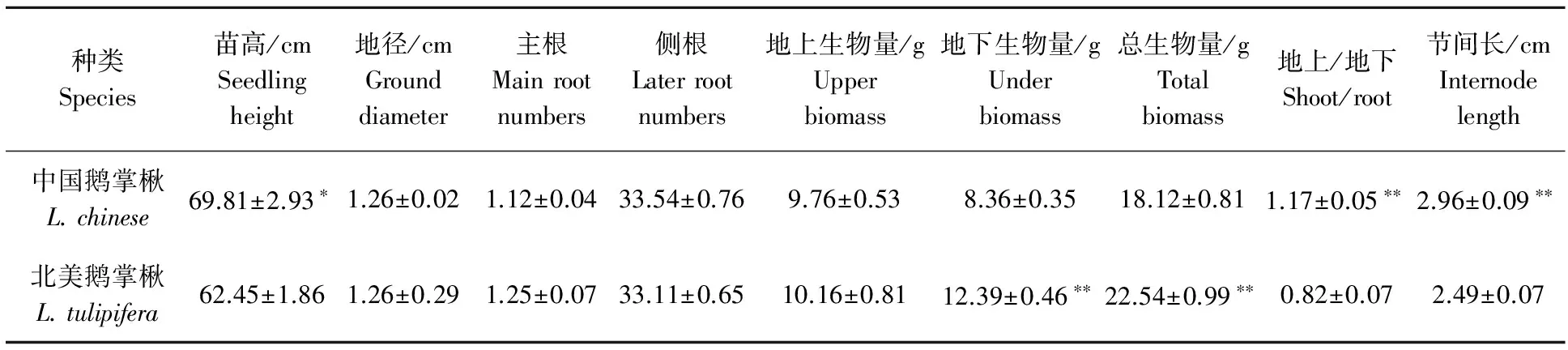
表3 中国鹅掌楸与北美鹅掌楸幼苗各性状特征

图2 不同种源鹅掌楸苗高、地径、生物量聚类分析Fig.2 UPGMA cluster based on seedling height,ground diameter and biomass characters of Liriodendron
2.1.4 不同种源幼苗苗高、地径和生物量的聚类分析 以幼苗苗高、地径和生物量特征为指标,对15个中国鹅掌楸种源进行聚类分析(图2)。图2以欧氏距离10为阈值,参试所有种源可以分为4个类群:第1类群为极慢生、低生物量类群,此类群只有DBS,该种源幼苗苗高、生物量平均值在各种源中均处于最低水平,地径处于较低水平;第2类群为慢生、低生物量积累类群,此类群为XY、YY、SZ和WYS这4个种源,该类种源幼苗苗高、地径生长相对较慢,生物量积累处于中等水平;第3类群为苗期中生、中生物量类群,此类群为EX、LP、LY、LS和FY这5个地理种源;第4个类群为苗期速生、高生物量类群,此类群为HS、SY、YN、MN和SN 这5个地理种源。
2.2 苗木物候期与生长节律
2.2.1 物候期 从表4可以看出,鹅掌楸物候期大部分种源表现基本一致,少数种源有所差异。出苗期以EX、HS、LS和WYS稍早一些,出苗时间在4月15日左右;在真叶期和木质化期中,以XY和YY、SN、DBS和北美种源晚一些;顶芽的形成多数种源在9月下旬到10月上旬,LS顶芽形成最早,9月中旬即可形成顶芽,YN最晚,顶芽形成期在10月中旬,且全年整株木质化程度较慢,苗杆与其它种源不同,表现为非常嫩绿;各种源叶变色期基本保持一致,日期为10月上、中旬。
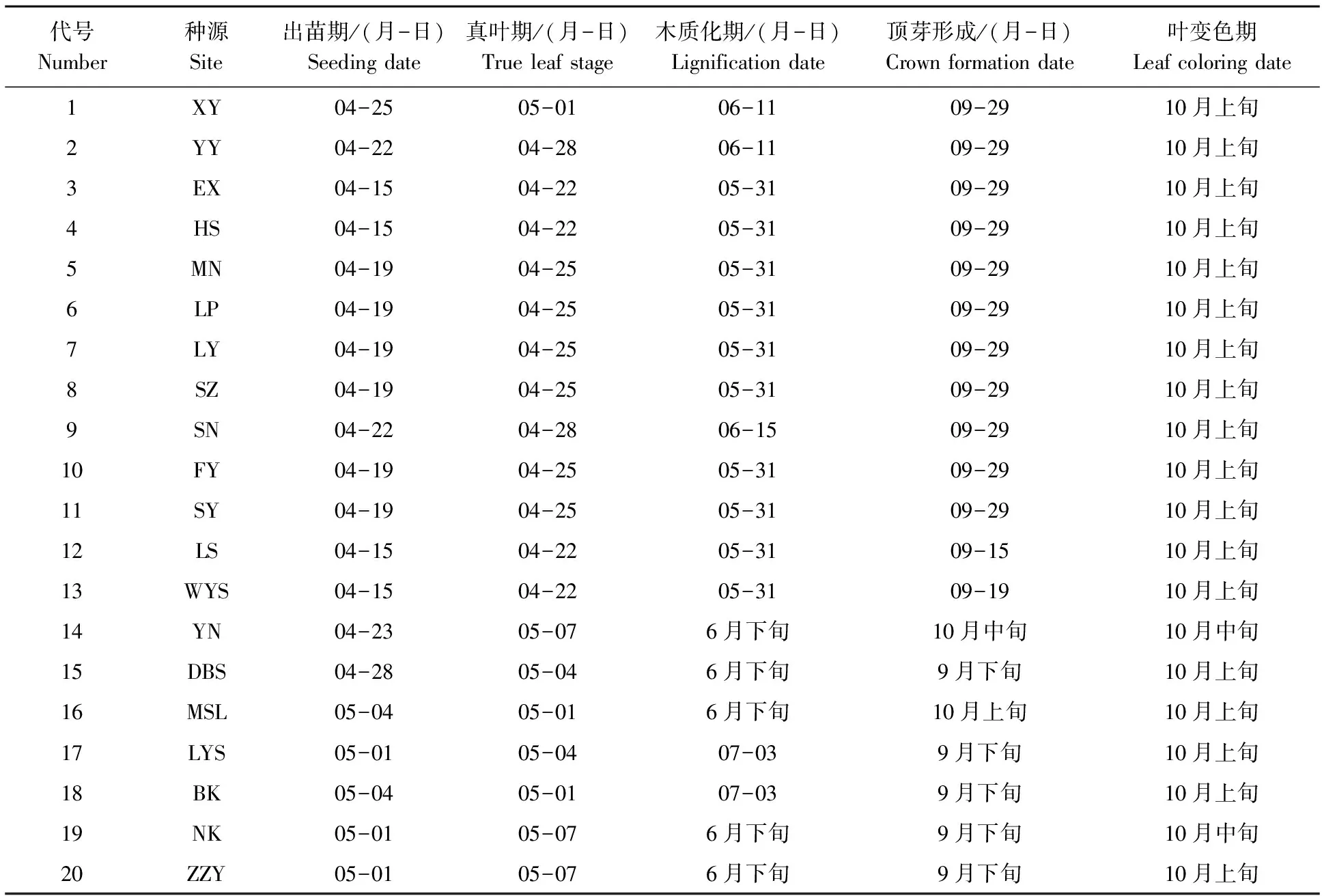
表4 鹅掌楸种源物候期观测
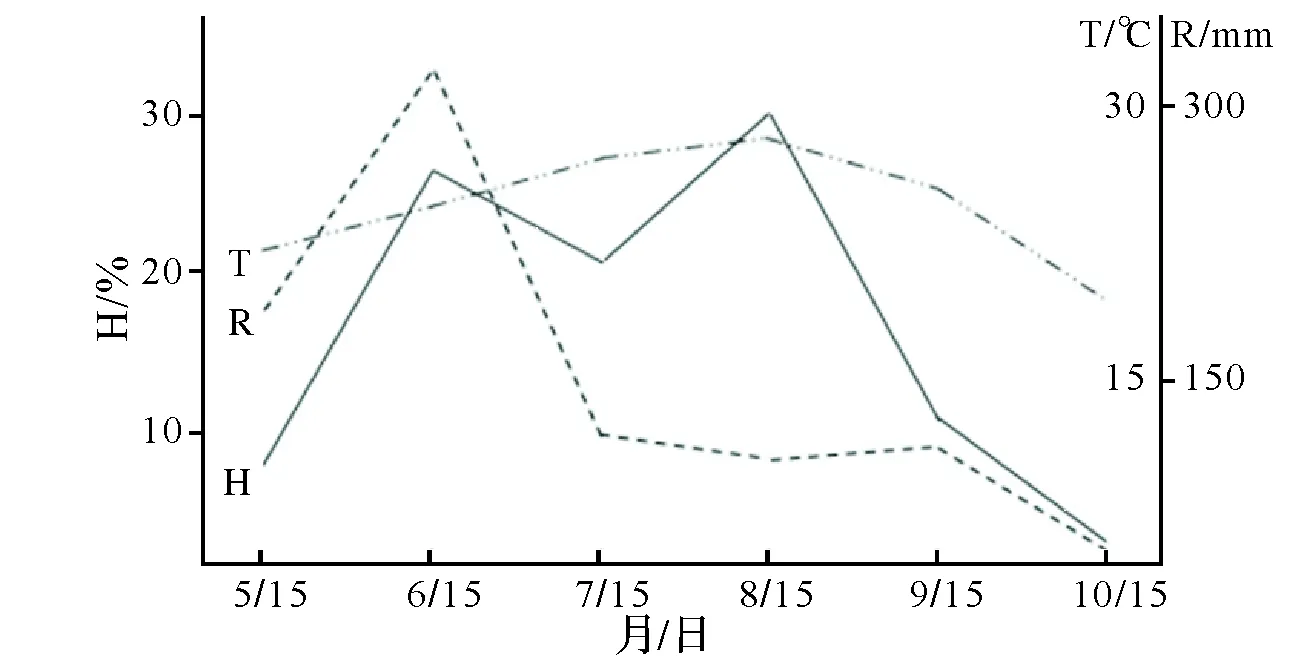
图3 鹅掌楸幼苗平均相对高生长与气温、降水量关系图Fig.3 Correlation analysis between average relative high growth of seedlings of Liriodendron and annual mean temperature and annual precipitation
经调查发现,北美鹅掌楸LYS、BK和NK在5—6月期间出现不同程度的烂梢现象,其中以LYS较重一些,引起的原因可能与这段期间降水量过多,相对湿度偏大有关。个别种源出现根腐病,如DBS,但程度较轻。在7月中旬到下旬期间,大部分种源幼苗出现整株死亡现象,其中较重一些的种源有EX、XY、SZ和LP,平均每重复10~12株。其它种源发生较轻,平均每重复1~4株,分析原因与这段期间持续高温干旱有关。幼苗越冬冻害情况,经调查发现,只有YN和XY两种源受到冻害,其中YN最为严重,幼苗冻害率达90%,冻害后的幼苗顶嫩梢出现枯死,一般为5~15 cm,高的达30 cm,四川叙永种源冻害较轻,危害率为5%。
2.2.2 生长节律 各种源幼苗苗高生长节律(H)与降水量(R)、温度(T)紧密相关(图3),大多数种源出现二次生长高峰,第一次在6月份,占全年生长量的26%;第二次出现在8月,占全年生长量的30%。但只有YN一年出现一次生长高峰(8月下旬—9月上旬),且表现生长期最长,在10月上旬还有较大的生长量。
2.2.3 中国鹅掌楸种源幼苗性状的地理变异规律 从表5可以看出:原产地年均气温与中国鹅掌楸苗高、地下生物量呈显著正相关关系,与地径和鲜重呈极显著正相关。表明苗高与地下生物量随着原产地年均气温的增加,呈显著增加的趋势,而地径和鲜重随着年均温的升高呈极显著增加的趋势。而苗木的其它性状与原产地生态因子间的相关关系不显著。说明原产地高气温种源幼苗生长较快。

表5 中国鹅掌楸幼苗性状与地理气候因子间的相关性分析
2.3 鹅掌楸苗期与采种母树主要性状的相关关系
从表6可以看出,鹅掌楸幼苗性状中苗高、地径、鲜重与冠幅呈显著负相关关系(P<0.05),说明鹅掌楸母树冠幅越大,子代幼苗的苗高、地径、鲜质量具有下降的趋势。幼苗侧根数与胸径呈显著负相关关系,说明母树的胸径越大,子代幼苗的侧根数呈现显著减少的趋势。而母树与子代幼苗的其它性状间没有相关性(P≥0.05)。
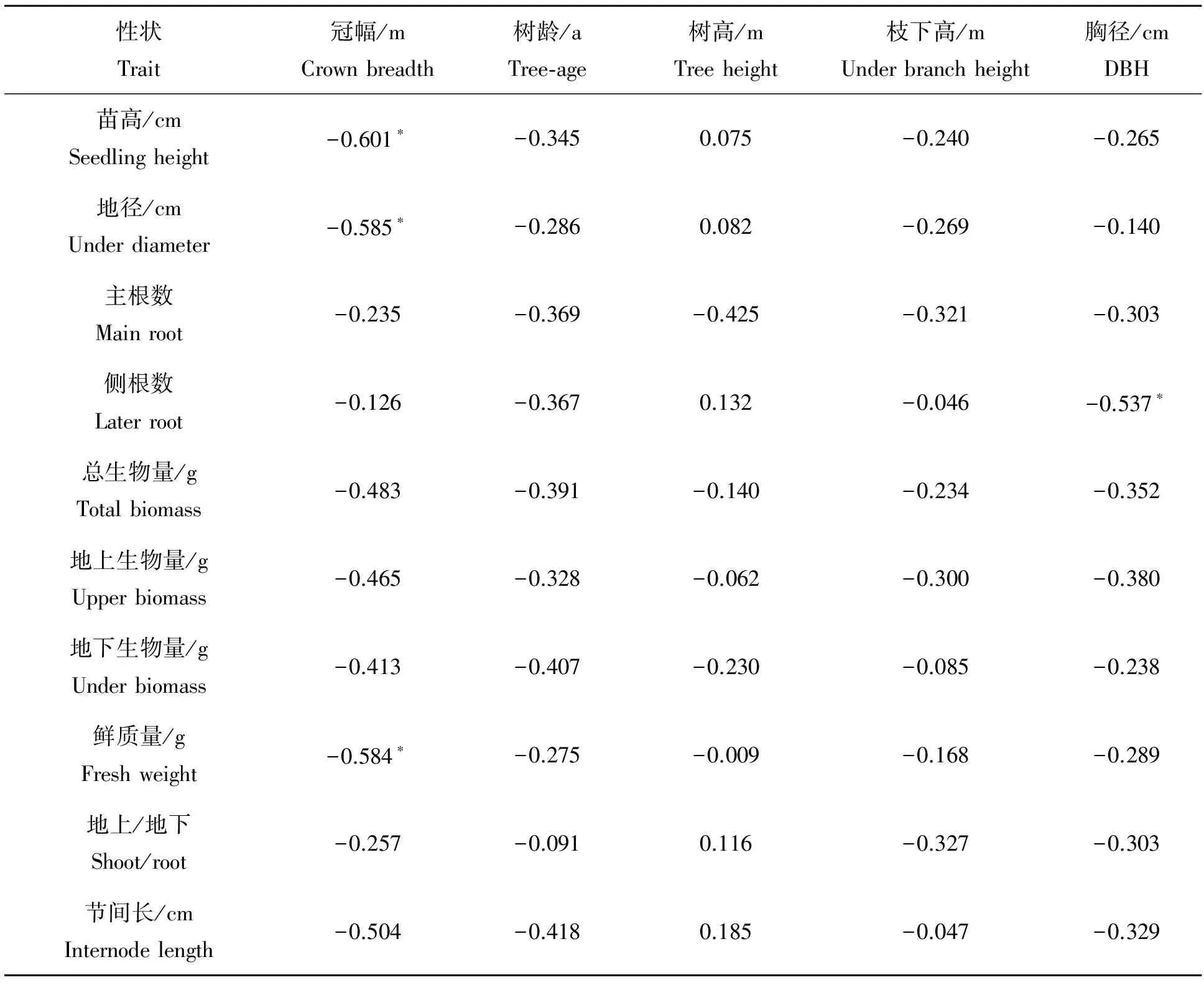
表6 鹅掌楸苗期性状与采种母树主要性状的相关关系
3 结论与讨论
(1)中国鹅掌楸与北美鹅掌楸幼苗生长性状与生物量性状比较表明,中国鹅掌楸苗高显著高于北美鹅掌楸,植株节间长和地上生物量/地下生物量极显著高于北美鹅掌楸。而在地下生物量与总生物量方面中国鹅掌楸种源极显著低于北美鹅掌楸。说明中国鹅掌楸与北美鹅掌楸幼苗个体发育构型上发生了显著性的变化。
本研究揭示了种源变化的一般生长规律。通过胸高、地径和生物量聚类分析、H×D平均值超过15%以及种源的抗逆性表明,SN、SY、HS和MN为本地表现优良的种源,因此,利用来自这些地区的种子或苗木作为研究地生产育苗的种质材料,将获得良好的苗期表现和较高的气候适宜可靠性,DBS、SZ、XY、YY、YN和北美的5个种源为研究地表现较差种源,在生产中杜绝使用。该结论与李斌等[21]对7年生鹅掌楸种源进行材性分析所选的优良种源不一致(其研究结果为LP和XY);也与董纯等[20]鹅掌楸地理种源试验研究报告结果(其排名前4种源为:HS、XY、LS和MN)有所出入,说明长的快不一定材性好,也有可能生长阶段不一致所导致,建议再对成熟林进行相关分析选择。
(2)各种源幼苗物候期大部分表现基本一致,其中以高海拔的山地种源物候出现得要早一些。一般各种源幼苗高生长一年出现2次高峰期,但只有YN出现一次生长高峰。幼苗前期高生长节律与温度和降水量有关。原产地年均气温与各种源苗高、地下生物量呈显著正相关关系,与地径和鲜质量呈极显著正相关关系,说明来自高气温种源在本地生长表现较快。
诸多研究[21-24]证实许多树种的性状都受到地理、生态等因子的影响,形成了不同的地理变异模式。本研究表明,不同种源鹅掌楸在苗高、地径、物候期和抗逆性方面存在显著性差异,说明鹅掌楸这些苗期性状也具有显著的地理变异。苗木前期高生长节律与温度和降水量有关;原产地年均气温与各种源苗高、地下生物量呈显著正相关关系,与地径和鲜质量呈极显著正相关关系。说明,鹅掌楸苗期表型性状、生长节律和气候适宜性的地理变异可能更多的与原产地温度、降水等因子影响密切相关。
来自北美鹅掌楸的几个种源,在苗木高生长前、中期出现烂梢现象,说明北美种源苗期不耐本地高温高湿气候。
(3)鹅掌楸幼苗性状苗高、地径、鲜质量与采种母树冠幅呈显著负相关关系,幼苗侧根数与采种母树胸径呈显著负相关关系,说明母树胸径越大,子代幼苗的侧根数呈现显著减少的趋势。
在鹅掌楸良种选育过程中,可以利用这些相关关系,选育出适宜生产要求的种质材料,如用于宽冠幅的城市绿化苗木可选择苗高、地径和鲜质量较小的苗木,而高密度用材林可选择这些指标相对较大的植株,将会缩短选育周期,提高选育成效。
[1] 吴东驰.优良高山用材观赏珍贵树种——鹅掌楸[J].农业现代化研究,1986,7(3):42-46.
Wu D C.Excellent mountain timber ornamental precious tree species-Liriodendron[J].Research on agricultural modernization,1986,7(3):42-46.
[2] 顾万春.主要阔叶树种速生丰产技术[M].北京:中国科技出版社,1993,64-78.
Gu W C.Fast growing and high yielding techniques of main broad leaved tree species[M].Beijing:China Science and Technology Press,1993,64-78.
[3] 王章荣.鹅掌楸属树种杂交育种与利用[M].北京:中国林业出版社,2005.
Wang Z R.With the use ofLiriodendronhybrid breeding[M].Beijing:China Forestry Publishing House,2005.
[4] Wolfe J A.Late cretaceous-cenoroic history of deciduousness and the terminal cretaceous event[J].Paleobiology,1987,13(2):215-226.
[5] Wolf e J A.Paleocene floras from the Gulf of Alaska region[J].Professional Paper-U.S.[J].Geological Survey,1997,8(2):100-108.
[6] Parks C R,Wendel J F,Sewell M M,et al.Genetic control of isozyme in genusLiriodendron[J].Journal of Heredity,1990,81(4):317-323.
[7] Harlow W M,Harrar E S.Textbook of dendrology[M].McGraw-Hill Book Company,1994,397-400.
[8] 方炎明,尤录祥.中国鹅掌楸天然群体与人工群体的生育力[J].植物资源与环境,1994,3(3):9-13.
Fang Y M,You L X.Fecundity of natural and cultivated populations ofLtriodendronchinense(Hemsl.) Sarg[J].Journal of Plant Resources and Environment,1994,3(3):9-13.
[9] 尹增芳,樊汝汶.中国鹅掌楸与北美鹅掌楸种间杂交的胚胎学研究[J].林业科学研究,1995,8(6):605-610.
Yin Z F,Fan R W.The embryological studies of interspecific hybrids betweenLiriodendrontulipiferaL,andL.chinense(Hesml.)Sarg[J].Forestry Research,1995,8(6):605-610.
[10] 尹增芳,樊汝汶.中国鹅掌楸花粉败育过程的超微结构观察[J].植物资源与环境,1997,6(1):1-7.
Yin Z F,Fan R W.The ultrastructure observation on the abortion ofLiriodendronchinensepollen[J].Journal of Plant Resources and Environment,1997,6(1):1-7.
[11] 孙亚光,李火根.利用SSR分子标记检测鹅掌楸雄性繁殖适合度与性选择[J].分子植物育种,2008,6(1):79-84.
Sun Y G,Li H G.Variations of male reproductive success and sexual selection among individuals of the experimental population ofLiriodendronrevealed by SSR markers[J].Molecular Plant Breeding,2008,6(1):79-84.
[12] 潘文婷,姚俊修,李火根.鹅掌楸属树种自交衰退的SSR分析[J].林业科学,2014,50(4):32-38.
Pan W T,Yao J X,Li H G.Analysis of selfing depression inLiriodendronwith SSR markers[J].Scientia Silvae Sinicae,2014,50(4):32-38.
[13] 刘洪鄂,沈湘林,曾玉亮.鹅掌楸、北美鹅掌楸及其杂种在形态和生长性状上的遗传变异[J].浙江林业科技,1991,11(5):18-23.
Liu H E,Shen X L,Zeng Y L.Genetic variations in morphological and growth characteristics of Chinese,american tulip-trees and their hybrid[J].Journal of Zhejiang Forestry Science and Technology,1991,11(5):18-23.
[14] 陈世群,杨永兰,郑先宝,等.鹅掌楸扦插繁殖技术初探[J].重庆林业科技,1993,36(1/4):30-33.
Chen S Q,Yang Y L,Zheng X B,et al.Preliminary study on on cuttage reproduce ofLiriodendronchinense[J].Chongqing forestry science and technology,1993,36(1/4):30-33.
[15] 杨志成.杂种鹅掌楸扦插试验初报[J].林业科学研究,1994,7(6):697-700.
Yang Z C.Preliminary study on hybrid tulip tree cutting rooting rates[J].Forestry Research,1994,7(6):697-700.
[16] 廖明,韦小丽,朱忠荣,等.鹅掌楸播种苗生长发育规律及育苗技术研究[J].贵州林业科技,2005,32(1):20-23.
Liao M,Wei X L,Zhu Z R,et al.Study on growth regularity and cultivation technology ofLiriodendronchineseseedlings[J].Guizhou Forestry Science and Technology,2005,32(1):20-23.
[17] 王章荣.鹅掌楸属杂交育种成就与育种策略[J].林业科技开发,2008,5(22):1-4.
Wang Z R.Achievements and strategies of hybrid breeding inLiriodendron[J].China Forestry Science and Technology,2008,5(22):1-4.
[18] 仝伯强,鲁仪增,李文清.不同无纺布袋类型对北美鹅掌楸袋装栽培后生长的影响[J].山东林业科技,2013,43(3):45-47.
Tong B Q,Ru Y Z,Li W Q.No effect of different woven bag types on growth ofLiriodendrontulipiferaafter bagged cultivation[J].Journal of Shandong Forestry Science and Technology,2013,43(3):45-47.
[19] 彭秀,耿养会,李秀珍,等.鹅掌楸容器育苗轻基质配方研究[J].湖北林业科技,2013,42(1):5-8.
Peng X,Geng Y H,Li X Z,et al.Study on container seedling cultural techniques of light media forLiriodendronchinense[J].Hubei Forestry Science and Technology,2013,42(1):5-8.
[20] 董纯,谭德仁,汪长江,等.马褂木地理种源试验研究报告[J].湖北林业科技,1999,28(1):3-10.
Dong C,Tan D R,Wang C J,et al.Research report on the geographical provenance test result ofLiriodendron[J].Hubei Forestry Science and Technology,1999,28(1):3-10.
[21] 李斌,顾万春,夏良放,等.鹅掌楸种源遗传变异与选择评价[J].林业科学研究,2001,14(3):237-244.
Li B,Gu W C,Xia L F,et al.Genetic variation and provenance selection of Chinese Tuliptree[J].Forestry Research,2001,14(3):237-244.
[22] 许洋,李迎超,冯慧,等.不同种源栓皮栎种子表型性状的变异分析[J].安徽农业科学,2015,43(25):164-167.
Xu Y,Li Y C,Feng H,et al.Variation of seed traits in different Provenance ofQuercusvariabilis[J].Journal of Anhui Agricultural Sciences,2015,43(25):164-167.
[23] 刁松锋,邵文豪,姜景民,等.基于种实性状的无患子天然群体表型多样性研究[J].生态学报,2014,34(6):1451-1460.
Diao S F,Shao W H,Jiang J M,et al.Phenotypic diversity in natural populationsofSapindusmukorossibased on fruit and seed traits[J].Acta Ecologica Sinica,2014,34(6):1451-1460.
[24] 杨传平,杨书文,夏德安,等.长白落叶松生长性状的地理变异规律与模式的研究[J].东北林业大学学报,1991,40(S2):9-18.
Yang C P,Yang S W,Xia D A,et al.Study on the geographic variation rule and pattern of growth characters ofLarixolgensis[J].Journal of Northeast Forestry University,1991,40(S2):9-18.
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
小星星·阅读100分(低年级)(2022年9期)2022-09-23 07:30:52
现代园艺(2021年12期)2021-12-07 03:59:22
鸭绿江(2021年17期)2021-10-13 07:06:16
辽宁林业科技(2021年1期)2021-03-16 09:17:08
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
现代农业科技(2017年24期)2018-01-22 21:35:06
