
沙地生境不同苜蓿品种形态特征对低温的响应及其与抗寒性关系
2018-02-13 01:42朱爱民张玉霞王显国田永雷丛百明候文慧
草地学报 2018年6期
朱爱民, 张玉霞, 王显国, 田永雷, 丛百明, 王 鑫, 候文慧
(1. 内蒙古民族大学, 内蒙古 通辽 028041; 2. 中国农业大学动物科技学院, 北京 100083;3. 内蒙古农业大学草原与资源环境学院, 内蒙古 呼和浩特 010018)
紫花苜蓿(MedicagosaticaL.)是一种多年生豆科(Leguminosaesp.)优质牧草[1],具有很强的适应性,在我国大部分地区均有栽培种植,因其具有品质好、抗逆性好、并且产量高等优点,在我国畜牧业产业中占据了重要地位[2-3]。随着国家粮改饲政策的调整,我国牧草的种植将不断扩大,其中内蒙古自治区提出到2020年发展1 000万亩节灌饲草地[4]。紫花苜蓿作为重要的冷季型牧草,在中国北方地区种植,越冬期的低温胁迫是影响其安全越冬的主要环境因素[5-7],低温锻炼期和越冬期寒害限制了北方草原牧区苜蓿的大规模生产,认识和研究紫花苜蓿越冬期抗寒机理,已经成为草原牧区研究热点之一。
低温锻炼期苜蓿首先感受到低温的是地上茎和叶,因此首先对低温做出反应的是苜蓿的茎和叶,其次是苜蓿的地下根系。目前,关于低温胁迫下苜蓿生长特性的研究很多,主要集中在苜蓿的茎、叶、茎叶比、根冠比、分枝数、株高、茎粗、秋眠芽数、根重、根长和叶片结构等方面。苜蓿低温锻炼期即秋末至初冬时期,在此期间日照短且温度逐渐降低。CofFindaffer,Nittler[8-9]研究表明,不同苜蓿品种在适应低温期间生长特性表现差异较大,抗寒性不同,耐寒能力强的苜蓿品种其茎分枝且匍匐生长。也有学者[10]研究发现苜蓿株高与抗寒性密切相关。崔国文等[11]在研究苜蓿生长特性与苜蓿抗寒性关系时表明,苜蓿叶片宽长比与苜蓿的越冬存活率呈显著负相关关系,抗寒性高的苜蓿品种其叶片宽长比越大,越冬率也就越高,叶片宽长比越小则苜蓿抗寒性就越低,越冬率越低。
根系是牧草重要的吸收器官和合成器官,根系的生长状况及其活力水平直接影响牧草的品质和产量。苜蓿作为宿根牧草,主要依赖根部的活力渡过越冬期,所以,苜蓿根系活力在很大程度上不仅能衡量其生长状况,更能反映出植株抗寒性的强弱。研究表明,宿根花卉、“野牛草”(Buchloedactyloides)等植物在低温胁迫下[12-14],根系活力均与植物抗寒性呈显著的相关关系。可见根系活力是衡量植物抗寒性的重要指标,并且实践中应结合更多的抗寒指标,综合评价和判断植物抗寒性。科尔沁沙地冬季寒冷、降雪较少,选择种植抗寒性强的苜蓿品种尤为重要。本研究通过对低温锻炼期沙地苜蓿各形态指标的测定,同时采用根系活力法协同Logistic回归方程计算苜蓿半致死温度,分析苜蓿的抗寒性与各形态指标的相关关系,为沙地苜蓿种质筛选、品种抗寒性鉴定及后续高抗寒性品种的选育提供理论依据。
1 材料与方法
1.1 试验区自然概况
试验地在内蒙古自治区赤峰市阿鲁科尔沁旗草源合作社公司(116°21′120°58′ E,41°17′45°24′ N),温带半干旱大陆性气候。年平均气温0~6℃,≥10℃积温3 000~3 200℃,无霜期140~150 d,年平均降水量350~400 mm,蒸发量是降水量的5倍左右,年平均风速34.4 m·s-1。试验田为新开垦草地,土壤类型为沙土。
1.2 供试材料
供试材料见表1。

表1 紫花苜蓿品种及来源Table 1 Alfalfa varieties and sources
1.3 试验方法
1.3.1大田试验设计 大田小区布局及管理:采用随机区组设计,小区试验面积20 m2(5 m×4 m),8个品种,3次重复,共设24个小区,小区之间设50 cm过道。所有小区底肥均施750 kg·hm2安琪酵母有机肥(有效活菌数≥0.20 亿(g-1,有机质含量≥60.0%),300 kg·hm2过磷酸钙(P2O5≥14.0 %),7 kg·hm2硫酸钾(K2O≥50.0%),60 kg·hm2尿素(N≥46.0%)。2016年8月4日播种,苜蓿播种量为22.5 kg·hm2,人工撒播,试验田正常管理。低温锻炼期(2016年9月15日、10月1日和10月15日)进行相关生长指标的测定,10月15日测鲜草产量。
1.3.2低温处理试验设计 实验室低温处理:11月20号取样,每小区取长势一致的越冬器官若干(苜蓿根系),于实验室中4℃冰箱保存,备用。将备用1年生苜蓿越冬器官用蒸馏水冲洗干净,每个品种平均分成7份,每份40株,每份越冬器官并排整齐摆放在厚度为3 mm,长、宽为30 cm×20 cm的纯棉布上,包裹好,均匀喷洒30 ml蒸馏水,使棉布保持潮湿状态,最后用长×宽为30 cm×30 cm锡箔纸包好。其中1份放入4℃恒温冰箱作为对照,其余6份放入低温冰箱(温度误差±1℃)进行处理,处理温度分别为-10,-15,-20,-25,-30和-35℃。低温程控箱设置为:以4℃为起点,先以4℃·h-1的速率升温,到达设定温度后保持8h,后以4 ℃·h-1的速率降温,取出后置于4℃下保持12 h,然后进行根颈活力测定,指标测定时采用随机取样。
1.4 生长指标和根颈活力测定
苜蓿干草产量:选择样区内长势一致的区域(避免取根样的区域),取2 m2样方刈割测量,留茬高度5 cm,3次重复取其平均值,即为苜蓿干重。鲜草每次测产时,称取200 g鲜草装袋带回实验室,105℃杀青 15 min后,在 65℃下烘干至恒重,计算鲜干比(200 g/干草重量g×100%),并进一步折算为每公顷干草产量。
株高:选择样区内长势一致的区域,样区内随机测定10株,测量从地面至植株顶端的自然长度,10次重复取其平均值。
根颈粗:使用游标卡尺直接测量,最大根颈处(产生分蘖和秋眠芽处),10次重复取其平均值。
秋眠芽:每小区取长势一致的苜蓿10株,分别数出每株苜蓿根颈处长出的秋眠芽数,记录并统计后计算平均单个苜蓿品种每株秋眠芽数。
分枝数:每小区取长势一致的苜蓿10株,分别数出每株苜蓿根颈处长出大于15 cm的枝条,记录并统计后计算平均单个苜蓿品种单株枝条数,即为苜蓿有效单株分枝数。
地上干物质量及根干重:每次刈割测定地上部株高时,同时挖取地下根系,连续5株,地上部分和地下部分分离,洗净后放在烘箱里于65℃烘干后分别进行称量。
根长:每次刈割测定地上部株高时,同时挖取地下根系,连续10株,用直尺测定苜蓿根系长度,记录并计算平均单株根长。
根冠比(%)=(地上干物质量/地下干物质量)×100%。
根系活力测定用TTC染色法测定(张志安和陈展宗方法[15]):采用TTC氧化还原法,称取根颈0.1 g,浸没在由0.4% TTC和磷酸缓冲液(pH 7.0)各5 mL的溶液内,在37℃下暗保存4 h,此后加入1 mol·L-1硫酸2 mL停止反应。将根放入研钵中,加乙酸乙酯3~4 mL充分研磨,将红色提取液移入刻度试管,用乙酸乙酯冲洗数次并定容至10 mL,用分光光度计在波长485 nm下比色,以空白试验(将根先用硫酸处理)作参比测出吸光度,查标准曲线,即可求出TTC还原量。
根系活力(mg.g-1.h-1)=四氮唑还原量(mg)·[根重(g)×时间(h)]-1。
将各处理温度下的相对根系活力分别用SPSS17.0软件拟合Logistic方程[16-20],求拐点温度,即为低温半致死温度(LT50)。
1.5 数据分析
试验数据用Microsoft Excel软件处理、做图和制作表格,SPSS17.0软件进行方差显著性分析,Logistic方程拟合和相关性分析。其中不同时期不同品种间株高、单株分枝数、秋眠芽数等指标数据采用方差显著性分析,低温处理下根颈活力值拟合Logistic方程。
2 结果与分析
2.1 沙地苜蓿对低温的地上生长响应
2.1.1沙地苜蓿低温锻炼期株高变化 根据表2可知,低温锻炼期不同时间测定8个苜蓿品种株高变化存在差异。9月15日测定“肇东”和“公农1号”苜蓿品种植株高度显著高于10月1日和10月15日测定株高(P<0.05),说明“肇东”和“公农1号”苜蓿品种秋季休眠早;10月15日测定“东苜1号”和“草原3号”苜蓿株高显著高于9月15日和10月1日测定株高(P<0.05),说明“东苜1号”和“草原3号”秋季休眠较晚,10月15日仍继续生长。
低温锻炼期8个苜蓿品种间株高变化不同。9月15日测定“公农1号”品种株高显著高于其他苜蓿品种(P<0.05),达38.8 cm,苜蓿株高最低的是草原3号品种,为24.5 cm,显著低于其他苜蓿品种(P<0.05)。10月1日测定和10月15日测定株高最高的均是“龙牧801”品种。

表2 沙地苜蓿低温锻炼期株高变化Table 2 Changes in plant height of alfalfa during cold hardening in sandy land/cm
注:不同小写字母表示差异显著性水平P< 0.05,”/”左边表示同列相同时期测定不同苜蓿品种间差异显著性,右边表示同一品种不同时间测定差异显著性,下同
Note:The different letters indicate significant differences at the 0.05 levels,on the left side of”/” indicate significant difference of the same column between different alfalfa varieties that determined at the same time,on the right side of”/” indicate significant difference between the same varieties that determined at the same time,the same as below
2.1.2沙地苜蓿低温锻炼期分枝数变化 低温锻炼期沙地苜蓿单株分枝数呈先增加后减少的趋势(表3)。10月1日测定8个苜蓿品种单株分枝数均高于9月15日测定分枝数,但未达到差异显著水平(P>0.05)。除“龙牧801”和“肇东”苜蓿品种外其他苜蓿品种9月15日和10月1日测定单株分枝数均显著高于10月15日测定单株分枝数(P<0.05),说明10月1日后苜蓿进入休眠,不产生分枝,且部分枝条凋零干枯减少。
低温锻炼期8个苜蓿品种间单株枝条数变化不同。如表3所示,9月15日测定“草原2号”苜蓿单株分枝数显著高于其他苜蓿品种(P<0.05),达到每株2.58个。10月1日测定苜蓿单株分枝数较多的是“草原2号”、“龙牧801”和“东苜1号”品种;10月15日测定“龙牧801”苜蓿品种单株分枝数显著多于其他苜蓿品种(P<0.05)。

表3 沙地苜蓿低温锻炼期单株分枝数变化Table 3 Variation of branch number of alfalfa during cold hardening period in sandy land/个·株-1
2.1.3沙地苜蓿低温锻炼期地上干物质量变化 如表4所示,低温锻炼期沙地苜蓿地上干物质量变化存在差异。“龙牧801”、“公农一号”、“农菁1号”和“农菁8号”苜蓿地上干物质量随测定时间的推迟呈逐渐增加的趋势。9月15日测定“草原2号”苜蓿地上干物质量显著大于10月1日和10月15日测定干物质量(P<0.05)。10月1日测定“肇东”、“东苜1号”和“草原3号”苜蓿地上干物质量显著高于9月15日和10月15日测定干物质量(P<0.05)。
低温锻炼期8个苜蓿品种间地上干物质量变化不同。9月15日测定“龙牧801”苜蓿品种地上干物质量最大;10月1日测定地上干物质量较大的苜蓿品种是“龙牧801”、“东苜1号”和“农菁8号”,分别为5.05 g、4.98 g和4.65 g;10月15日测定“农菁8号”苜蓿地上干物质量最大。
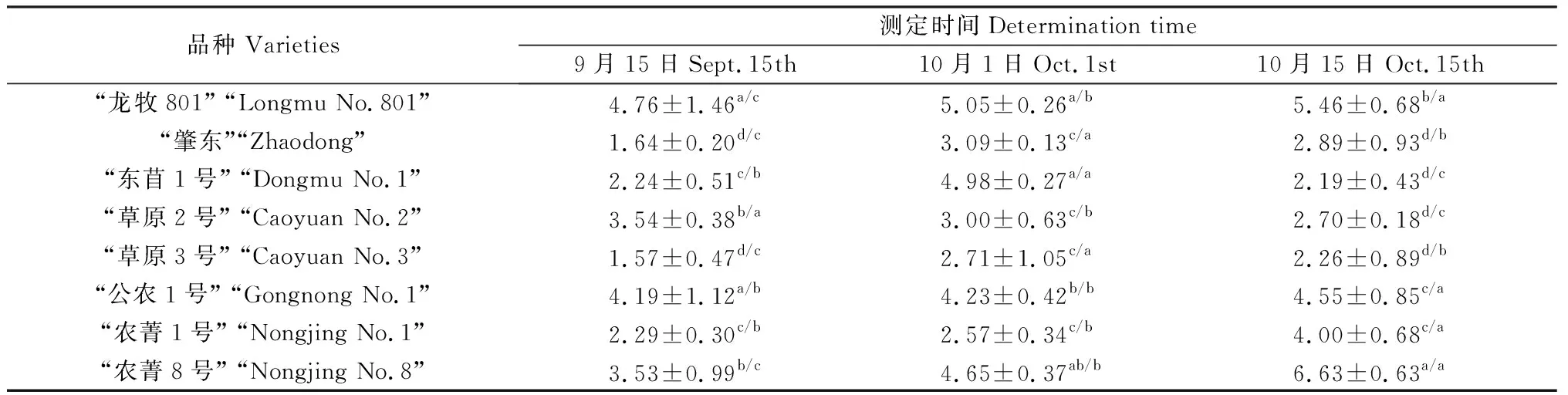
表4 沙地苜蓿低温锻炼期地上干物质量变化Table 4 Dry matter quality changes of alfalfa above ground during cold hardening period in sandy land/g·5株-1
2.1.4苜蓿品种干草产量比较 如图1所示,“肇东”和“农菁8号”苜蓿品种干草产量显著高于其他苜蓿品种(P<0.05),分别达到2 681.33 kg·hm-2和2 645.01 kg·hm-2,干草产量最低的是“草原3号”苜蓿,为2 248.08 kg·hm-2,显著低于“龙牧801”、“肇东”、“东苜1号”、“农菁1号”和“农菁8号”,其中“龙牧801”、“东苜1号”、“草原2号”、“公农1号”和“农菁1号”苜蓿品种间干草产量无差异显著性(P>0.05)。

图1 苜蓿品种干草产量比较Fig.1 Comparison of hay yield of alfalfa varieties注:A:龙牧801;B:肇东;C:东苜1号;D:草原2号;E:草原3号;F:公农1号;G:农菁1号;H:农菁8号不同小写字母表示不同品种间差异显著水平P<0.05,下同Note:A:Longmu No.801;B:Zhaodong;C:Dongmu No.1;D:Caoyuan No.2;E:Caoyuan No.3;F:Gongnong No.1;G:Nongjing No.1;H:Nongjing No.8 Different lowercase letters indicate significant difference among different varieties at the 0.05 level,the same as below
2.2 沙地苜蓿对低温的地下生长响应
2.2.1沙地苜蓿低温锻炼期秋眠芽数变化 低温锻炼期苜蓿休眠芽数随时间的推移呈现增加的趋势(表5)。但是不同品种休眠芽的变化存在差异。9月15日测定“公农1号”和“农菁1号”苜蓿品种秋眠芽数显著高于其他苜蓿品种(P<0.05),分别是单株2.67个和2.50个。10月1日测定“草原3号”、“草原2号”和“东苜1号”苜蓿秋眠芽数相对较多,分别为单株4.08个、3.96个和4.00个。10月15日测定“龙牧801”和“农菁1号”苜蓿秋眠芽数显著高于其他苜蓿品种(P<0.05),分别为单株4.83个和4.91个。
2.2.2沙地苜蓿低温锻炼期颈粗变化 根据表6可知,低温锻炼期不同时间测定苜蓿根颈粗度变化趋于一致。10月15日测定8个苜蓿品种颈粗均大于10月1日测定颈粗,其中“草原3号”、“公农1号”、“农菁1号”和“农菁8号”颈粗达到差异显著水平(P<0.05),且10月15号测定颈粗均显著高于9月15日测定颈粗(P<0.05),除“草原3号”苜蓿品种外其他苜蓿10月1日测定颈粗均显著高于9月15日测定颈粗(P<0.05)。三次测定“龙牧801”苜蓿品种根颈均最粗。
2.2.3沙地苜蓿低温锻炼期根重变化 低温锻炼期沙地苜蓿根重呈增加的趋势(表7)。除“草原2号”苜蓿品种外10月15日测定其他苜蓿品种根重均显著大于10月1日测定的根重(P<0.05),除“肇东”苜蓿品种外10月1日测定其他苜蓿品种根重均显著大于9月15日测定的根重(P<0.05)。
低温锻炼期8个苜蓿品种间根重变化不同。9月15日测定“公农1号”、“草原2号”、“龙牧801”和“农菁8号”苜蓿品种根重显著高于其他苜蓿(P<0.05)。10月1日测定“东苜1号”苜蓿品种根重最大,“肇东”和“农菁1号”苜蓿根重相对较低。10月15日测定“草原3号”苜蓿根重最大,为3.46 g,显著大于其他苜蓿品种(P<0.05)。

表5 沙地苜蓿低温锻炼期秋眠芽数变化Table 5 Changes in the number of dormant buds during the cold hardening period of alfalfa in sandy land/个·株-1

表6 沙地苜蓿低温锻炼期颈粗变化Table 6 Changes in neck diameter of alfalfa duringcold hardening in sandy land/ mm

表7 沙地苜蓿低温锻炼期根重变化Table 7 Root weight changes during cold hardening of alfalfa in sandy land/ g
2.2.4沙地苜蓿低温锻炼期根长变化 根据图2所示,低温锻炼期沙地苜蓿根长增加,且不同苜蓿品种在不同时间段根长增加幅度不同。9月15日至10月1日苜蓿根长增加达到差异显著水平的品种是“龙牧801”、“肇东”、“草原2号”、“草原3号”、“公农1号”和“农菁8号”(P<0.05)。10月1日至10月15日8个苜蓿品种根长均增加,“肇东”、“东苜1号”、“草原2号”和“农菁1号”增加差异达到显著水平。
2.2.5沙地苜蓿低温锻炼期根冠比变化 如图3所示,8个苜蓿品种低温锻炼期根冠比变化趋于一致,均呈增加趋势,且不同品种间根冠比增加幅度不同。9月15日测定除“农菁1号”外,其他苜蓿品种根冠比均显著低于10月1日测定的根冠比(P<0.05),其中根冠比较高的是“草原3号”苜蓿,为0.50。10月1日测定根冠比最高的是“农菁8号”品种,达到0.66,除“公农1号”和“农菁8号”外,其他苜蓿品种根冠比均显著低于10月15日测定的根冠比(P<0.05)。10月15日测定根冠比最高的是“草原3号”苜蓿品种,为1.32,其次是草原2号,为1.13,根冠比相对较低的是“龙牧801”品种和“公农1号”品种。说明低温锻炼期沙地苜蓿地上生物量部分向地下转移,地下生物量明显增加,根冠比增大。

图2 沙地苜蓿低温锻炼期根长变化Fig.2 Root length changes during cold hardening of alfalfa in sandy land

图3 沙地苜蓿低温锻炼期根冠比变化Fig.3 Change of root and crown ratio of alfalfa during cold hardening in sandy land
2.3 根颈活力法协同Logistic回归方程对8个苜蓿品种半致死温度分析
根据在不同低温胁迫下苜蓿根颈活力变化值求得logistic回归方程,低温半致死温度,相关系数(r)及显著性(Sig.)见表8。8个苜蓿品种显著性均小于0.01,相关系数均大于0.9,达到差异极显著水平;“东苜1号”苜蓿半致死温度最低,为-19.2℃,其次是“草原3号”苜蓿,其中“草原2号”和“农菁8号”品种苜蓿半致死温度均高于-15℃,最低半致死温度与最高半致死温度相差4.76℃;根据根颈活力法协同Logistic回归方程得半致死温度对8个苜蓿品种耐寒性排序为,“东苜1号”>“草原3号”>“农菁1号”>“公农1号”>“龙牧801”>“肇东”>“农菁8号”>“草原2号”。
2.4 苜蓿半致死温度与各形态指标相关性
根据表9可知,苜蓿半致死温度与株高、茎粗、分枝数、地上干物质和根长呈正相关关系,但均未达到显著水平(P>0.05),与秋眠芽数、根重和根冠比呈负相关关系,其中与根重的相关性达到差异显著水平(P<0.05)。此外苜蓿株高与分枝数、茎粗与地上干物质量以及根长与秋眠芽数呈显著正相关关系(P<0.05)。其他生长指标间无显著相关性(P>0.05)。

表8 8个苜蓿品种低温处理下的根颈活力回归方程及半致死温度(LT50)Table 8 Logistic equation between temperature and vitality of root neck of 8 alfalfa varieties and their LT50.
注:**表示在0.01水平下差异显著
Note:** indicates significant difference at the 0.01 level
注:*表示在 0.05 水平上显著相关
Note:* indicate significant correlation at the 0.05 level
3 讨论
因地制宜的选择苜蓿品种是保证苜蓿安全越冬的首要条件,在中国北方寒冷地区,品种的选择应首先考虑抗寒性。因此根据生产目标、当地气候条件、现场调查、引种试验等多方而因素综合分析,确定适宜品种,才能保证苜蓿安全越冬[4]。本试验在北方科尔沁沙地对8个苜蓿品种低温锻炼期生长特性调查表明,“肇东”和“公农1号”苜蓿品种植株在9月下旬后不在继续生长,株高呈降低趋势,说明“肇东”和“公农1号”苜蓿品种秋季休眠早,低温锻炼期3次测定“草原3号”苜蓿株高均明显低于其他苜蓿品种,说明当“草原3号”苜蓿株体接收到低温胁迫后逐渐进入休眠,可能具有较强的抗寒性。苜蓿颈粗为适应逐渐降低的温度表现为显著增粗的变化。Schwab等[21]研究表明,苜蓿根颈粗度与其抗寒性具有显著正相关关系,本试验9月15日和10月1日测定“龙牧801”苜蓿品种根颈粗度均显著大于其他苜蓿,因此可推断“龙牧801”苜蓿品种抗寒性较强。
王英哲等[22]在研究苜蓿引种和生产性能比较中表明不同紫花苜蓿品种分枝数表现不同,本试验表明苜蓿单株分枝数9月15日和10月1日相比变化不显著(P>0.05),之后单株分枝数呈减少趋势,且不同品种间苜蓿单株分枝数不同,与王英哲等[23]研究结果一致。10月1日后北方地区温度逐渐降低,抗寒性强的苜蓿品种地上物质量的积累停滞,部分营养向根系转移,“肇东”、“东苜1号”和“草原3号”苜蓿地上干物质量在10月1日后显著降低,说明此3种苜蓿对低温较敏感;“农菁8号”苜蓿品种干草产量显著高于其他苜蓿品种(P<0.05),干草产量最低的是“草原3号”苜蓿,说明“农菁8号”品种对低温的敏感度低,在逐渐降低的温度变化环境中,地上生物量积累较其它苜蓿品种高,而“草原3号”苜蓿品种则逐渐进入休眠。
苜蓿秋眠芽数越多则翌年苜蓿的分枝越多,产量亦会提高,秋眠芽的多少与越冬的关系尚不明确。本试验表明苜蓿根颈秋眠芽数随逐渐降低的温度表现为显著增加的变化(P<0.05),相关性分析表明苜蓿秋眠芽数与颈粗呈显著正相关关系,其中10月15日测定“龙牧801”和“农菁1号”苜蓿秋眠芽数显著高于其他苜蓿品种(P<0.05),说明“龙牧801”和“农菁1号”苜蓿生产性能高于其他苜蓿品种。陶雅、陈积山、吴新卫、王运涛等[24-27]研究发现苜蓿根颈粗度的大小与苜蓿的抗寒性呈显著的正相关关系,本研究与其研究结果一致,本研究表明随季节性温度的逐渐降低,苜蓿根重明显增加,根长增长,根冠比增大,10月15日测定根颈粗度最大的苜蓿品种是“龙牧801”,“草原3号”苜蓿根最重,推断两苜蓿品种抗寒性较强。
受不同低温胁迫后苜蓿植株根颈活力会明显下降,且温度越低根颈活力越弱,根颈活力法协同Logistic回归方程表明,“东苜1号”品种抗寒性较“草原3号”强,“农菁8号”品种较“草原2号”抗寒性强;8个紫花苜蓿品种抗寒性排序为“东苜1号”>“草原3号”>“农菁1号”>“公农1号”>“龙牧801”>“肇东”>“农菁8号”>“草原2号”;通过对半致死温度与苜蓿各生长指标做相关性分析表明,苜蓿半致死温度与苜蓿根重呈显著负相关关系(P<0.05),即苜蓿根系越重其半致死温度越低、抗寒性越强,而与其他生长指标无显著相关关系(P>0.05)。
4 结论
低温锻炼期“东苜1号”、“草原3号”和“公农1号”对低温较敏感,苜蓿休眠较早;科尔沁沙地可选择种植“东苜1号”、“草原3号”等抗寒性较强的苜蓿品种。
猜你喜欢
军事文摘(2021年18期)2021-12-02
内蒙古林业(2021年6期)2021-06-26
幼儿智力世界(2021年4期)2021-05-17
人民黄河(2020年12期)2020-12-30
山西农业科学(2020年9期)2020-09-14
山西农业科学(2019年12期)2019-12-19
科普童话·百科探秘(2018年7期)2018-09-13
电子制作(2017年10期)2017-04-18
新疆农垦科技(2016年2期)2016-08-21
中国果菜(2016年9期)2016-03-01
