
冀北山地不同海拔天然山杏种群的结构与动态
2018-02-13 07:26程子敏徐学华李晓刚李玉灵
西北植物学报 2018年12期
程子敏,王 南,穆 枫,徐学华,李晓刚,李玉灵
(河北农业大学 林学院,河北保定071000)
植物种群结构与动态是种群生态学研究的重要内容[1],树木生长对气候的响应是沿海拔梯度发生变化的,研究不同海拔种群的结构和动态不仅可以反映种群的生存现状,而且能够揭示植物与外界环境相互作用的结果,对植物资源的保护以及利用有着重要的意义[2-3]。种群的径级结构不仅可以了解种群现在的结构动态,而且还可以分析种群过去的结构以及预测未来种群结构的动态变化,有助于揭示种子散布以及幼苗建立等特征在时间尺度上对植物种群动态变化的影响[4]。数量动态指数、静态生命表、存活曲线和谱分析是研究种群结构与动态的重要方法。其中数量动态指数能够弥补植物种群结构动态评价中较为粗放的等级归类划分的缺陷[5]。种群数量动态变化特征往往通过编制其生命表来予以定量化[6],通过生命表的编制得出种群的死亡率、消失率等重要参数值,同时绘制其存活曲线,从而为种群数量统计和种群生态学研究提供更多的信息[7]。谱分析是描述由不同的振幅和相应的谐波组成复杂的周期现象,种群天然更新的波动性和年龄更替过程的周期性可以通过种群不同龄级的株数分布波动来表现[8]。近年来已有学者对不同地区[9-10]、不同生境[11-14,4]、不同干扰[15-16]下的种群结构与动态进行过研究,但针对不同海拔种群结构与动态的研究相对较少。
山杏(Armeniacasibirica)为蔷薇科植物,通常为落叶乔木或灌木,具有喜光、抗旱、抗低温、耐贫瘠、耐盐碱和根系发达的特点,是冀北山地春季开花植物和山地造林的重要树种,具有一定的生态、景观和观赏价值,以及明显的防风固沙和水土保持效益[17]。其形成的天然山杏种群在该地区沿海拔梯度有较大面积的分布,但目前针对山杏的研究多集中在山杏的遗传变异[18-21]、栽培、嫁接技术[22]、光合、蒸腾速率[23-24]和持水性能[25-26]等方面。但该地区山杏种群的数量动态规律沿海拔梯度如何发生变化?以及山杏种群对冀北山地天然灌木群落稳定性的维持机制尚不明确。因此本文以冀北山地3个海拔区域的天然山杏种群为研究对象,从种群的径级结构、静态生命表、存活曲线、死亡曲线以及谱分析等方面进行研究,旨在阐明不同海拔天然山杏种群的结构及数量动态特征,进而揭示海拔对山杏种群数量动态的影响以及山杏种群对群落稳定性的维持机制,以期为该地区天然山杏种群及群落的保护和管理提供科学依据。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于河北省承德市丰宁县。地势由东南向西北呈阶梯状增高,海拔800~1 500 m。气候属于中温带半湿润半干旱大陆性季风型高原山地气候,年均温0.9~6.2 ℃,年降水量350~550 mm。土壤以砂石土为主。植被类型主要有针叶林、针阔混交林、落叶阔叶林、灌丛和灌草丛等。典型乔木树种以油松(Pinustabuliformis)、白桦(Betulaplatyphylla)、华北落叶松(Larixprincipis-rupprechtii)和蒙古栎(Quercusmongolica)为主;灌木主要有山杏、绒毛绣线菊(Spiraeavelutina)、木香薷(Elsholtziastauntoni)、胡枝子(Lespedezabicolor)、小叶鼠李(Rhamnusparvifolia)等;草本植物以细叶苔草(Carexrigescens)、细裂叶蒿(Artemisiatanacetifolia)、丛生隐子草(Cleistogenescaespitosa)、牛尾蒿(Artemisiadubia)、狗牙根(Cynodondactylon)和披碱草(Elymusdahuricus)等为主。以天然山杏为优势种的灌木群落在该地区有较大面积的分布。
1.2 研究方法
1.2.1样地调查在全面踏查的基础上,采用样地调查法于2017年7~8月对丰宁县山地进行实地调查,选取山杏种群相对集中、林分保持良好且无人为干扰的典型阳坡地段,沿海拔700 m~1 400 m设置3块50 m×50 m的标准样地,按海拔梯度划分为3个生境:872 m(H1)、1 131 m(H2)、1 328 m(H3)。不同海拔天然山杏种群的树高、胸径、密度和盖度等均具有较大的差异。采取相邻网格样方分割法将每个样地划分成25个10 m×10 m的样方,利用游标卡尺和钢卷尺对样方内所有山杏及其幼苗进行每木检尺,记录每株山杏的地径、树高以及幼苗的地径与苗高等,同时测定样地的海拔、坡度和植被盖度等。样地基本概况见表1。
1.2.2种群径级结构的划分由于受环境因素的影响,该地区天然山杏常呈簇状生长,因此,调查时以基株为单位进行统计,若一个树桩上有多个分株,则按径级最大的一株为准。本研究采用空间替代时间的方法,以种群立木径级结构代替年龄结构来分析种群动态的变化[27-28]。根据山杏生物学特性与年轮的变化规律,以及梭梭(Haloxylonammodendron)[11]种群径级的划分方法进行山杏种群径级的划分,基径≤2 cm时,基径按每0.5 cm划分为一个等级,0 cm<Ⅰ≤0.5 cm,0.5 cm<Ⅱ≤1 cm,1 cm<Ⅲ≤1.5 cm,1.5 cm<Ⅳ≤2 cm;基径>2 cm时,基径按每1 cm划分为一个等级,2 cm<Ⅴ≤3 cm,3 cm<Ⅵ≤4 cm,4 cm<Ⅶ≤5 cm,…,13 cm<ⅩⅤⅠ≤14 cm,14 cm<ⅩⅤⅡ≤15 cm。H1、H2和H3海拔山杏种群分别划分为15、17和11个径级。
1.2.3种群动态量化方法采用陈晓德[29]量化方法对种群动态进行定量描述,具体计算方法如下:
(1)
(2)
式中:Vn为种群从n到n+1级的个体数量变化,取正、负、零时分别反映种群两相邻年龄级个体数量的增长、衰退和稳定的结构动态关系;Vpi为整个种群结构的数量变化动态指数;Sn、Sn+1分别为第n和n+1级种群个体数;K为种群大小级数量;式(2)仅适用于不考虑未来外部环境干扰的种群结构动态的比较,若要考虑外部干扰时,则种群结构动态还与大小级数量(K)及各大小级个体数(S)两因素相关,因此将式(2)修正为:
(2′)
(3)
1.2.4种群静态生命表编制以冀北山地不同海拔天然山杏种群各径级的株数为依据,编制其静态生命表。生命表中:x为径级;ax为x径级的个体数;因本研究中各径级的个体数相差较大,生命表中会出现死亡率为负值的情况,因此,采用匀滑技术[30]对数据进行处理,得出存活数ax*,存活量lx=ax*/a0×1 000,标准化存活数的对数为Inlx,死亡量dx=lx-lx+1,死亡率qx=dx/lx,区间寿命Lx=(lx+lx+1)/2,总寿命Tx=∑lx,期望寿命ex=Tx/lx,消失率Kx=lnlx-lnlx+1。

表1 冀北山地天然山杏样地基本概况
1.2.5谱分析方法谱分析方法能够揭示种群数量的周期性波动,山杏种群的天然更新过程可以通过不同径级株数分布的波动从而得以体现。谱分析是Fourier级数的展开,复杂的周期现象可以由不同振幅和相对应的谐波组成,因此,写成正弦波形式[31]:
(1)
式中:A0为周期变化的均值;Ak(k=1,2,3,…,P)为各谐波的振幅,标志各个周期的作用大小,其值的差异,反应了各周期作用大小的差别;θk为各谐波的相角,ωk为各谐波的频率;Nt为t时刻的种群大小,n为时间系列总长度。
将山杏种群各年龄个体分布视为一个时间系列t,以Xt表示t年龄序列时的个体数;n同样为时间序列总长度;P=n/2为谐波的总个体数为已知;T为正弦波的基本周期,即时间系列t的最长周期,则可以用下式来估计Fourier分解中的各个参数,即:
(2)
Ak2=ak2+bk2
(3)
ωk=2πk/T
(4)
θk=arctg(ak/bk)
(5)
(6)
(7)
2 结果与分析
2.1 不同海拔天然山杏种群的径级结构及动态
种群的径级结构可以代替年龄结构来反映种群的生存状况。冀北山地不同海拔天然山杏种群的径级结构见图1。由图1可知,各海拔山杏种群径级结构大致呈倒“J”型分布,3个海拔山杏种群发展均趋于稳定态势。H1(872 m)、H2(1 131 m)、H3(1 328 m)山杏种群的Ⅰ径级和Ⅱ径级所占比例分别为50.37%、72.19%和45.11%,其后随径级的增大个体数呈下降的趋势。虽然不同海拔梯度各径级个体比率差异较大,但总体上,H1、H2、H3山杏种群径级结构相似,个体数集中于小中径级,表明3个海拔山杏种群均以幼年和青壮年植株为主,种群有充足的中小个体作为储备资源,为增长型的年龄结构。H1、H2和H3山杏种群各径级株数总和分别为2158株、1262株和133株,株数总和随海拔的增高呈减少趋势,这与该地区特殊的地形起伏变化以及海拔升高温度降低和山杏自身的生物学特性、生态学习性有关。不同海拔天然山杏种群径级结构分布模式各不相同,H1最大为径级,对应基径大小为12.47 cm,H2最大为ⅩⅤⅡ径级,对应基径大小为14.10 cm,H3海拔山杏种群有11个径级,对应最大的基径大小为8.76 cm,表明海拔越高,天然山杏种群径级分布范围越小,种群密度也越小。但最大径级呈现出H2>H1>H3的特征,可能与海拔变化所引起的种群密度的差异有关。

H1、H2、H3分别表示海拔高度为872 m、1 131 m、1 328 m。 径级划分标准为,当基径≤2 cm时,基径按每0.5 cm划分 为一个等级,0 cm<Ⅰ≤0.5 cm,0.5 cm<Ⅱ≤1 cm, 1 cm<Ⅲ≤1.5 cm,1.5 cm<Ⅳ≤2 cm;基径>2 cm时, 基径按每1 cm划分为一个等级,2 cm<Ⅴ≤3 cm,3 cm< Ⅵ≤4 cm,4 cm<Ⅶ≤5 cm,…,13 cm<ⅩⅤⅠ≤14 cm, 14 cm<ⅩⅤⅡ≤15 cm。下同。图1 冀北山地不同海拔天然山杏种群的径级结构 H1, H2 and H3 show the altitude of 872 m, 1 131 m and 1 328 m, respectively. The diameter divided standard is, when the diameter≤2 cm, diameters are divided into one grade per 0.5 cm, 0 cm<Ⅰ≤0.5 cm,0.5 cm<Ⅱ ≤1 cm,1 cm<Ⅲ≤1.5 cm,1.5 cm<Ⅳ≤2 cm; when the diameter>2 cm, diameters are divided into one grade per 1 cm, 2 cm<Ⅴ≤3 cm,3 cm<Ⅵ≤4 cm,4 cm<Ⅶ≤5 cm, …,13 cm<ⅩⅤⅠ≤14 cm,14 cm<ⅩⅤⅡ≤15 cm. The same as blowFig.1 Diameter structure of natural Armeniaca sibirica populations at different altitudes in mountain areas of northern Hebei Province

2.2 不同海拔天然山杏种群的静态生命表
静态生命表是利用特定的时间段所调查的种群所有的个体编制而成的。根据表3可知,H1、H2和H3海拔天然山杏种群的总寿命均呈现随径级的增大不断减小的趋势,显示逐步达到其生理寿命。不同海拔同一径级以及同一海拔不同径级山杏种群的期望寿命均存在波动。不同海拔天然山杏种群Ⅰ径级期望寿命均小于Ⅱ和Ⅲ径级,表明山杏种群在Ⅰ径级受到环境筛选的影响较大,到Ⅱ和Ⅲ径级种群个体适应性逐渐增强。随着山杏种群个体的生长,种群种内和种间竞争增加,期望寿命开始降低。但H1、H2和H3海拔山杏种群分别在Ⅹ径级、Ⅻ径级和Ⅶ径级期望寿命再次出现峰值,分别为2.88、2.67和2.50,说明山杏种群在经过中期激烈的种内种间竞争之后生存能力逐渐增强。此后,由于种群个体生理衰老等因素的影响,各海拔山杏种群期望寿命随径级的增加呈现下降的趋势,显示种群逐步到达生理寿命。
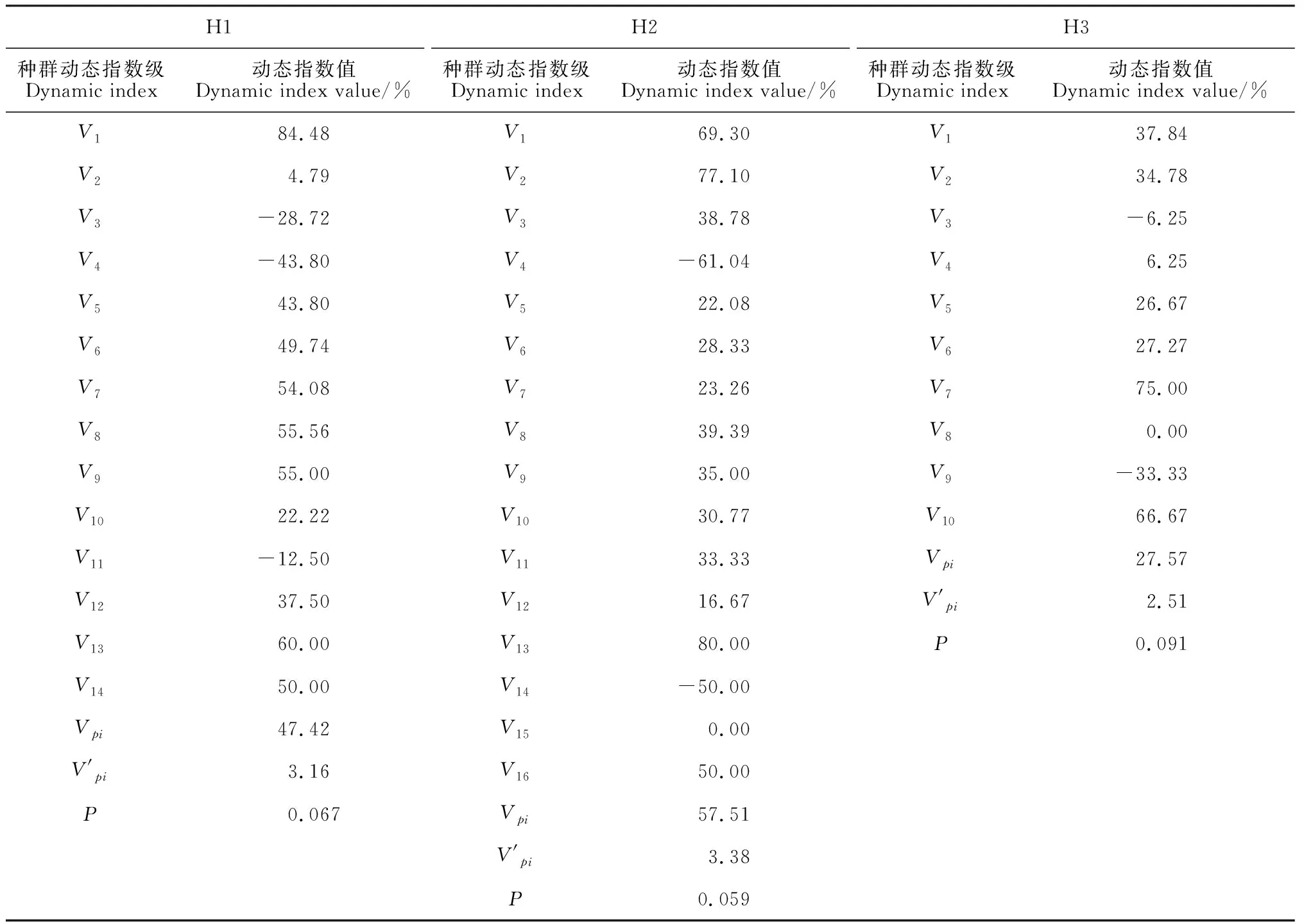
表2 冀北山地不同海拔天然山杏种群径级结构的动态变化指数



表3 冀北山地不同海拔天然山杏种群静态生命表
2.3 不同海拔天然山杏种群的存活曲线
存活曲线能够更为直观地表达种群个体在各径级的存活过程[32]。根据生命表所示数据,以径级为横坐标,标准存活量的对数lnlx为纵坐标绘制存活曲线。结合图2,采用2种数学模型:指数方程Nx=N0e-bx和幂函数Nx=N0x-b对Deevey-Ⅱ型和Deevey-Ⅲ型进行检验,运用SPSS统计软件建立相应的模型,通过对比F检验值和相关系数R2值得出,3个海拔山杏种群的指数模型R2以及F值均大于幂函数模型的相应值(表4),因此认为,3个海拔天然山杏种群存活曲线均趋于Deevey-Ⅱ型,说明山杏种群在各自生命过程中死亡率大致均衡,种群趋于相对稳定的状态。
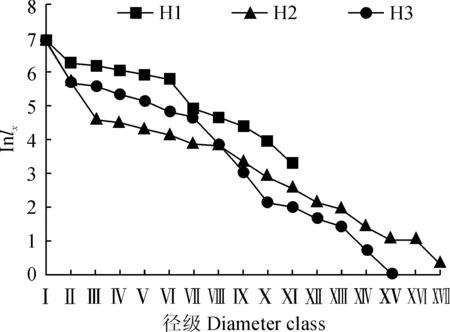
图2 冀北山地不同海拔天然山杏种群的存活曲线Fig.2 Survival curves of natural A. sibirica population at different altitudes in mountain areas of northern Hebei Province
2.4 不同海拔天然山杏种群的死亡率和消失率曲线
以不同海拔天然山杏种群的各径级为横坐标,静态生命表中死亡率(qx)和消失率(Kx)的值为纵坐标,绘制不同海拔天然山杏种群的死亡率及消失率曲线(图3)。由图3看出,随径级的增大不同海拔天然山杏种群的死亡率和消失率曲线呈波动型变化趋势且基本保持一致。H1、H2和H3海拔山杏种群在Ⅰ径级存在资源竞争和环境筛选,种群个体大量死亡,表现出较高的死亡率。随着种群个体的生长,对环境的适应能力增强,出现了相对稳定的时期。H1、H2和H3海拔山杏种群死亡率分别在Ⅸ径级、Ⅷ径级和Ⅵ径级出现二次峰值,且呈现出H1(0.60)>H3(0.58)>H2(0.39)的趋势,表明H1和H3海拔山杏种群中期种内种间竞争较为激烈。后期由于生理衰老等原因的影响,造成了各海拔山杏种群死亡峰期的出现。从整体上看,除Ⅰ径级外,3个海拔梯度天然山杏种群死亡率波动峰值处于不同径级。
2.5 不同海拔天然山杏种群的谱分析
不同海拔山杏种群的天然更新动态是通过不同径级的株数分布波动表现的,为了计算方便,本研究采用生命表的径级划分作为波谱分析的极差,由于H1、H2和H3山杏种群分别分为15、17、11个径级,所以采用内插法分别补充为16级、18级和12级,因此各海拔总波序(K=n/2)分别为8、9、6。由于各径级的个体数存在一定的差异,因而先对数据进行处理,以Xt′=In(ax+1)来代替公式中的Xt。利用波谱分析计算公式计算各个波形的振幅AK值(K=1,2,3,…,P)(表5)。A1为基波,A2~A8、A2~A9、A2~A6为不同海拔各个谐波,且每个谐波的周期为基本周期的1/2、1/3、…、1/P)。
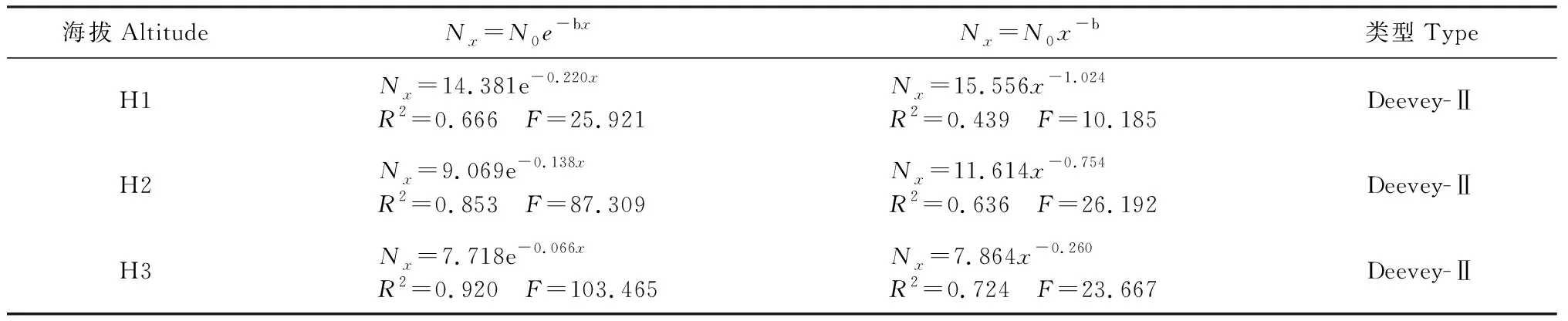
表4 不同海拔天然山杏种群存活曲线的拟合方程
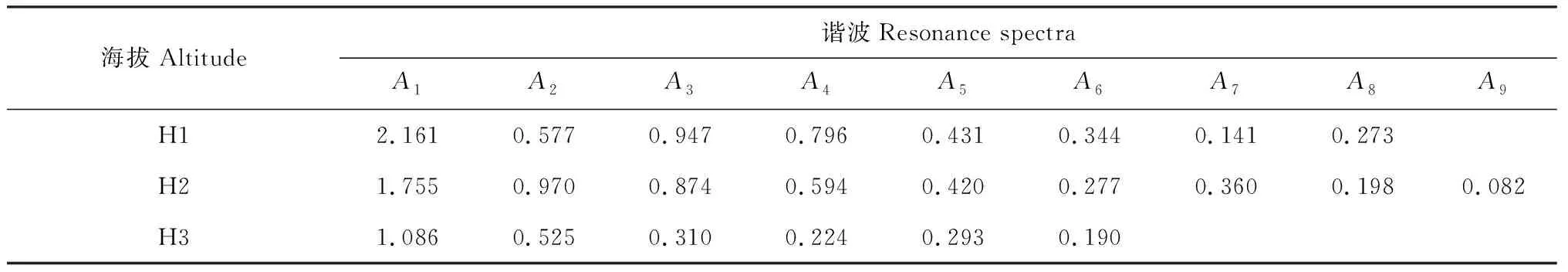
表5 冀北山地不同海拔天然山杏种群的周期性波动

图3 冀北山地不同海拔天然山杏种群的死亡率 (qx)和消失率(Kx)曲线Fig.3 Death rate and disappearance rate of natural A. sibirica populations at different altitudes in mountain areas of northern Hebei Province
从表5看出,不同海拔天然山杏种群天然更新波动明显,各海拔天然山杏种群的基波A1最大,分别为2.161、1.755和1.086,呈现出随海拔的增高不断减小的现象。基波表现了种群基本周期的波动状态,其周期长度是种群本身所固有的,由种群自身特性所决定的,这表明不同海拔天然山杏种群的数量动态变化受其整个生命周期过程自身特有的生物学特性的控制趋向较为明显。但由于所调查3个山杏种群的径级结构分布差异较大,因此无法对3个海拔的周期波动进行对比分析。但3个海拔天然山杏种群除受到基波的影响外,还表现出明显的小周期波动,H1海拔山杏种群分别在A2和A8出现波动, H2和H3海拔山杏种群分别在A7和A5处出现波动,但小周期的波动并不明显,说明各海拔山杏种群的发展过程较为平稳。
3 讨 论
3.1 不同海拔天然山杏种群结构及生存现状
植物种群数量动态不仅反映不同个体的组配情况,同时也反映了种群的结构、发展趋势以及与环境之间的相互关系,是揭示种群结构现状和更新策略的重要途径之一。经过对3个海拔天然山杏径级结构的对比分析,显示 3个海拔山杏种群的整体数量呈现出H1>H2>H3的数量动态特征,且径级分布表现出一定的差异性,主要是由于海拔升高,致使山杏种群对周围气候变化更加敏感,所承受的非生物环境压力增大,低温大风、土壤贫瘠以及生长期缩短等限制因子增多,因此天然山杏种群数量随海拔增高不断减少,同时说明山杏种群在海拔梯度上的径级分布与种群发育历史、物种特性以及环境因素有关[33]。
种群若缺乏可更新的幼龄个体将导致种群趋于衰退[34]。从天然山杏种群的幼苗更新数量来看,天然山杏的幼苗萌发更新能力极强,种群密度相对较大且具有一定的幼苗储备,种群表现出一定的更新发展潜力。但由于H3海拔天然山杏种群地处丰宁接坝和坝上交接地带,温度骤降,环境变化较为复杂,植被类型开始由灌丛向草原植被转变。虽无人为干扰,但幼苗难以承受低温和适应复杂多变的生存环境,成活率较低,且缺乏大径级植株,种群虽暂时处于稳定的状态,但未来的增长潜力较为不足。结合不同海拔天然山杏种群数量动态规律以及样地实地调查,表明海拔的高低是影响山杏生长与数量动态的重要因素。此外,本研究采用径级划分和种群结构动态指数两种方式来综合分析不同海拔山杏种群的结构,比仅用单一的年龄结构分析可能更为准确。
3.2 不同海拔天然山杏种群动态趋势
植物种群的发展是一个漫长而复杂的过程,不仅涉及到种内和种间的竞争,同样涉及到物种和无机环境之间的适应过程。静态生命表分析结果显示,3个海拔天然山杏种群Ⅰ径级期望寿命值较低,死亡率较高,这可能是由于幼苗存活能力较低,在营养竞争和环境筛选的过程中,部分幼苗被淘汰。此后成为Ⅱ径级以上的植株,山杏种群呈现相对稳定的发展态势,但随着植株个体以及树冠的增大,生存需要的光照、水分、养分和空间已不能满足植株个体的需求,种间竞争强烈,种群内的自疏和它疏作用增强,死亡率开始增加,期望寿命开始下降。随着种群个体的生长,存活植株开始进入自身数量调节后的稳定生活期,生存空间和养分、水分等相对充足。此后随着山杏种群逐渐步入衰老期,期望寿命值降到最低,死亡率达到峰值。
研究表明,3个海拔天然山杏的存活曲线呈Deevey-Ⅱ稳定性,这与新疆西天山峡谷不同海拔梯度上野核桃种群[4]以及河西走廊荒漠绿洲过渡区沙拐枣[35]的存活曲线研究结果相一致,原因可能与山杏种群经受的环境阻力和种内种间竞争有关,但存在交叉现象。H1、H2和H3海拔山杏种群在各自的生活史上分别出现7次、6次和3次死亡高峰,这可能与海拔升高生境变化引起山杏自身的生物学或生态学习性变化等有关,但有待于进一步研究,这对阐述山地植物在不同海拔梯度上的种群数量动态有重要的意义。
3.3 不同海拔天然山杏种群谱分析
山杏种群个体的增长和生活空间的不断扩展使得种群各径级植株的组配情况在不断发生变化,此后种群自身所创造的环境条件又反过来调节和控制种群的植株数量,产生波动的过程,这是稳定植物群落中优势种群的特征[36]。不同海拔天然山杏种群的谱分析结果显示,天然山杏种群数量动态是存在周期性波动的,而且所表现出来的波动并非单一周期,具有大周期内有小周期的谐波迭加特征,这与多数学者利用谱分析对种群数量动态的研究结果相一致[37-39]。研究发现,不同海拔山杏种群的基波随海拔的增高呈减小的趋势,这可能是与各海拔山杏种群的密度相关。由于所研究不同海拔天然山杏种群年龄不够大,其时间系列长度不足以表现出其基本周期和固定的周期长度,但其存在性是肯定的。在长白山臭冷杉[40](Abiesnephrolepis)种群研究中也有过类似的报道。除此之外,H1山杏种群分别在A2和A8处存在小周期波动,H2和H3山杏种群则分别在A7、A5处出现小周期波动。A2对应阶段,山杏种群幼苗经过高强度的环境筛选,在较高死亡率的基础上,一部分个体进入中龄阶段,种群密度开始下降,种群发展趋于稳定。A8、A7和A5处周期的波动与种群自身的生物学特性有关,经过一定的生长过程,此径级的山杏种群数量出现小波动现象。但由于各海拔山杏种群小周期的波动并不明显,表明山杏种群的发展过程较为平稳,仍能保持相对的稳定性。揭示冀北山地不同海拔天然山杏种群结构的动态变化规律及其天然更新过程的周期性,对该地区天然山杏种群和群落的保护和管理具有指导意义。
猜你喜欢
现代农村科技(2022年11期)2022-12-16
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
园艺与种苗(2021年3期)2021-05-26
现代农村科技(2021年3期)2021-04-11
中国农业信息(2017年16期)2017-11-02
运动(2017年1期)2017-09-03
种子科技(2017年8期)2017-09-03
西北农林科技大学学报(自然科学版)(2016年7期)2016-08-04
广西农学报(2015年4期)2015-09-11
