
生菜SSR遗传多样性及其与营养品质性状的关联分析
2018-02-13 12:14王书珍黄兴学王斌才周国林汪爱华
江苏农业科学 2018年24期
王书珍, 黄兴学, 王斌才, 张 霖, 周国林, 汪爱华
(1.经济林木种质改良与资源综合利用湖北省重点实验室/黄冈师范学院生命科学学院,湖北黄冈 438000;2.武汉市农业科学技术研究院蔬菜科学研究所,湖北武汉 430065)
生菜(LactucasativaL.)(2n=18)是菊科莴苣属1~2年生草本植物,原产地中海沿岸,现为全世界范围广泛种植食用的低糖、低脂类叶用类蔬菜,因能够分解亚硝胺等致癌物质被称为“抗癌蔬菜”,其根据叶片的长势分为结球生菜、半结球生菜、散叶生菜等多种类型[1-2]。生菜富含蛋白质、维生素、矿物质、有机酸、核黄素、膳食纤维等活性成分,具有较高的营养和经济价值[3]。近年来,随着生活水平的提高,人们对蔬菜、水果以及农作物等的品质要求不断提高,对生菜的品质要求也是逐渐提高[4]。
品质性状是数量性状或数量-质量性状,其受多基因控制,且其表型极易受到周围环境的影响[4]。控制品质性状的基因常为隐性基因,传统的品质检测繁琐且费用较高,因此农作物的品质改良工作进展缓慢。然而,寻找与品质性状紧密连锁的分子标记并用于分子标记辅助育种中,将大大提高品质性状的遗传改良效率。微卫星(simple sequence repeat,SSR)分子标记,是基因组中1~6个核苷酸多次串联重复组成的序列,数量丰富、共显性遗传、多态性高,已经广泛应用于遗传多样性、关联分析、基因定位、遗传连锁图谱构建、物种进化等研究中[5-6]。
本研究选用51份生菜资源构建自然群体,采用SSR标记分析其遗传多样性,并结合关联分析,挖掘蛋白质、硝酸盐、可溶性糖、硝酸盐含量等营养品质性状的优异等位变异,为生菜品质性状遗传改良育种奠定分子基础,也为分子标记辅助育种提供理论依据。
1 材料与方法
1.1 试验材料
从国家生菜种质资源库中挑选51份表型差异较大的资源,分别于2015、2016年秋季种植在武汉市蔬菜科学研究所的试验基地(114°20′E、30°37′N),种植2季,常规日常管理和病虫害防治,
定植株行距为25 cm,各品种种植30~50株。
1.2 营养品质性状测定
每个品种选取长势一致且健康的植株5株,在同一位置采摘叶片,低温保存后迅速进行营养品质的测定,各试验重复3次。蛋白质含量的测定采用微量凯氏定氮法[7];采用硝酸盐试粉法测定新鲜叶片组织中的硝酸盐含量[8]。采用蒽酮法测定生菜嫩叶的可溶性糖含量[9]。利用近红外光谱技术测定生菜叶片纤维素的含量[10]。
1.3 基因组DNA提取及PCR扩增
每个品种随机选5株,每株采集3张健康嫩叶,硅胶干燥并自封袋封存。采用改良的CTAB法提取基因组DNA,稀释到100 ng/μL[11]。依据NCBI数据库的生菜EST序列信息,设计合成80对SSR引物。PCR反应体系(15 μL):1 μL的基因组DNA(100 ng/μL),20 mmol/L正反向引物各0.15 μL,7.5 μL的2×TaqPCR Mastermix,灭菌的去离子水补足体积。PCR扩增程序:94 ℃预变性5 min;94 ℃变性40 s,最适退火温度下退火40 s,72 ℃延伸50 s,35个扩增循环;72 ℃延伸 5 min。采用6%的变性聚丙烯酰胺凝胶检测PCR扩增产物,依据20 bp DNA marker的电泳谱带,统计不同个体相应位点上等位基因的大小。
1.4 数据统计与分析
根据电泳图谱构建“0/1”二元矩阵:同一电泳位置上有带记为“1”,无带记为“0”。对蛋白质、硝酸盐、可溶性糖、纤维素含量的测定数据进行方差分析和聚类分析,统计各性状的平均值、变异系数、重复力(R)大小[12]。使用Arlequin 3.1软件统计等位基因数(Na)、期望杂合度(HE)、观察杂合度(HO)等多样性参数[11,13]。利用PICCalc 0.6软件计算各标记的多态性信息(PIC)含量。采用NTSYS-PC(version 2.2)软件计算遗传相似系数,根据遗传距离构建聚类图[14]。根据遗传距离矩阵分析SSR标记位点与形态性状的相关性[12]。
2 结果与分析
2.1 生菜营养品质性状分析
本研究所选的51份生菜资源变异较大,方差分析表明,4个营养品质性状在不同的生菜资源间均达到极显著水平。生菜叶片蛋白质平均含量高达10.914 mg/g,硝酸盐平均含量为1.016 mg/g,可溶性糖平均含量为2.286%,而纤维素平均含量为0.019%(表1)。蛋白质含量的变化幅度最大,为 2.211~35.423 mg/g,极差高达33.213 mg/g。4个营养品质性状的变异系数(CV)在0.124%~0.624%间变化,其中蛋白质含量的变异系数最大,最小的是纤维素含量。4个品质性状的重复力(R)变化范围为0.865~0.973,重复力最低的是纤维素含量,最高的是蛋白质含量(表1)。
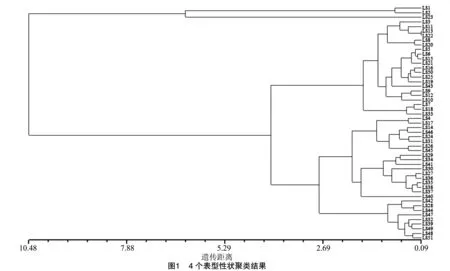
依据4个营养品质性状构建的聚类分析图,51份生菜资源被分为两大类,LS1、LS2、LS23等3份生菜构成第1类,其余的48份被聚为第2类(图1)。在遗传距离3.73处,第二大类又被分为2个小类,分别包括LS3、LS11等在内的21份资源和包括LS4、LS17等在内的27份资源。营养品质性状类似的生菜品种被聚到一起,研究发现聚类结果与地理起源并不一致。

表1 各品种间品质性状变异情况
2.2 SSR标记的多态性分析
在所设计合成的80个EST-SSR标记中有34个能够特异性扩增出目的大小的DNA片段,且重复性强;12个以(AG)n或者(CT)n为重复基序的标记在51份生菜资源中是多态的。多态性SSR标记的退火温度为53~58 ℃,微卫星基序重复次数介于24~27次之间。12个标记共扩增出等位基因类型62种,片段大小为124~237 bp。每个标记检测到的等位基因数为3~9个,平均为5.167个。标记LswSS10检测到的等位基因数最多,为9个,其次是LswSS6,检测到8个等位基因位点。LswSS1、LswSS9、LswSS11等3个SSR标记扩增效率最低,仅扩增出3个等位基因(表2)。
观察杂合度(HO)和期望杂合度(HE)的变化范围分别为0.00~0.273和0.545~0.842,平均值分别为0.106和0.687(表2)。12个标记间的连锁不平衡度并未达到极显著水平,并且12个位点处均出现HO小于HE的情况,即杂合子严重缺失。LswSS4、LswSS5、LswSS7、LswSS8等4个位点的HO均为0,即此4个位点在检测的生菜资源中均是纯合位点。多态性信息含量(PIC)值变化范围为0.428~0.803,平均为 0.614。除了LswSS11标记,其余11个位点的PIC值均大于或等于0.500,即生菜资源内存在丰富的遗传多样性。
2.3 营养品质性状与SSR标记关联分析
将4个营养品质性状的距离矩阵与SSR标记的距离矩阵进行相关性分析,拟合方程为y=0.464 8x+1.322,二者的相关系数为-0.02841,相关性并不显著。为了分析每个标记位点对4个营养品质性状的贡献率,将4个营养品质性状数据进行聚类,12个SSR扩增带谱也分别聚类,再将蛋白质、硝酸盐、可溶性糖和纤维素含量等4个性状的距离矩阵与单个SSR标记聚类的距离矩阵进行相关性分析。在进行的两两相关分析中,并未找到与蛋白质含量和硝酸盐含量性状关联的SSR分子标记。LswSS1和LswSS11两个标记与生菜叶片可溶性糖含量显著相关,相关系数分别为-0.214和 -0.260(表3)。与纤维素含量相关的SSR标记有3个,分别为LswSS1(r=-0.366)、LswSS3(r=-0.208)以及LswSS12(r=-0.261)。然而,包括LswSS2、LswSS4、LswSS5、LswSS6、LswSS7、LswSS8、LswSS9、LswSS10等在内的8个SSR标记与四个营养品质性状的相关性都不显著。

表2 12个SSR多态性标记信息

表3 与品质性状相关联的SSR标记信息
3 讨论与结论
关联分析(association analysis)是利用基因或者标记间的连锁不平衡(linkage disequilibrium,LD)关系,对表型性状与基因或标记位点间的相关性进行分析,从而鉴定与表型变异相关的基因位点[15]。关联分析采用的是自然群体,研究周期短,且可以同时分析同一位点上的多个等位基因,自然群体在长期进化过程中积累了大量的重组信息,做出来的关联图谱精确度也比较高[16]。于志远等利用关联分析方法检测到11个与大豆蛋白质含量极显著关联的SSR标记,解释率为2.65%~9.08%[17]。冯英娜等筛选到与茄子农艺相关性状显著相关(P<0.05)的SSR标记17个[18]。严玫等采用关联分析法检测出与花生品质性状关联的SSR位点4个,总等位变异位点40个[19]。
生菜叶片的营养品质性状属于连续变异的数量性状,是基因和环境共同作用的结果,其遗传基础复杂,目前由于很难找到表现型和基因型之间的对应关系,因此品质性状机理和遗传改良等研究十分困难。本研究将51份生菜资源种植在条件一致的苗圃地,采用完全随机区组设计,最大程度上消除了环境因素引起的个体表型差异。本研究共检测到的12个多态性位点均出现观察杂合度显著低于期望杂合度的现象,可能的原因是在生菜长期选育过程中,部分位点出现了极大的纯合化。最后利用关联分析法筛选出2个与生菜叶片可溶性糖含量相关的SSR标记,3个与纤维素含量相关的SSR标记,并且LswSS1标记与2个营养品质性状均相关。
本研究筛选到的营养品质性状关联的SSR标记对于后期克隆生菜营养品质性状相关目的基因意义重大,本研究结果也为生菜品质性状的遗传改良和新品种选育奠定了基础。然而,关联位点与性状间关系及作用机理和方式尚不清楚,而进行全基因组的关联分析则能找出更多与品质性状关联的分子标记,后续仍需更加深入的研究。
猜你喜欢
区域治理(2022年40期)2022-11-27
中老年保健(2022年6期)2022-08-19
新世纪智能(数学备考)(2021年9期)2021-11-24
农家参谋(2020年5期)2020-06-15
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
当代陕西(2019年15期)2019-09-02
学苑创造·A版(2018年11期)2018-02-01
食品与健康(2017年3期)2017-03-15
