
中缅树鼩社群行为的观察研究
2018-01-31 04:32侯东敏任晓英王政昆叶芳艳朱万龙
绿色科技 2018年2期
关键词:竞技场
侯东敏 任晓英 王政昆 叶芳艳 朱万龙
摘要:为了探究中缅树鼩(Tupaia belangeri)的社群行为规律,对中缅树鼩的社群行为进行了观察与统计分析。采用竞技场观察了中缅树鼩的社群行为。结果表明,中缅树鼩的社群行为中,成年雌性之间的友好行为较成年雌雄个体之间增加,成年雄性之间的攻击行为较成年雌雄个体之间增加。大多数雌性树鼩会选择争斗中占优势的雄性,说明雌性在选择配偶时可能和雄性的适合度有关。雄性或雌性均能优先识别出之前与之配对的树鼩,这可能与气味识别等有关;成年雌性间的友好相处和成年雄性间攻击行为较成年雌雄个体之间增加,符合中缅树鼬单雄配偶系社会结构。
关键词:中缅树鼩;社群行为;竞技场
中图分类号:Q95
文献标识码:A
文章编号:1674-9944(2018)2-0004-03
1 引言
社群行为对于动物种群的结构稳定至关重要,对动物社群行为的研究能够更为系统地理解其生态功能[1]。张建军等[2,3]对布氏田鼠( Lasiopodomys brandtii)社群行为的研究表明,雄性布氏田鼠的攻击行为并不因为雌性的存在而减少,而且雌性选择配偶会受到雄性等级的影响。除了对啮齿类动物社群行为的研究外,对非人灵长类的社群行为也进行了相关研究,如对黄山短尾猴( Macaca thibetana huangshanensis)研究发现,其社群等级关系能够通过友好行为得以体现[4]。除此之外,还有学者对根田鼠(Microtus oeconomus )[5]、大仓鼠( Tscherskia triton)[6]、黑线姬鼠(Apodemus agrarius)[7]等其他行为进行了研究。
中缅树鼩(Tupaia belangeri)属攀鼩目(Scanden-tia)树鼩科(Tupaiidae),分布于南亚、东南亚以及中国南部和西南部地区,为东洋界特有的昼行性小型哺乳动物。目前被广泛地应用于肿瘤学、免疫学、神经生物学和生殖生物学等方面的科学研究[8,9]。为满足生物学和医学领域相关科研实验的需求,急需对树鼩进行室内驯化饲养。本研究旨在探究中缅树鼩的社群行为,为中缅树鼩的成功驯养提供科学的行为学理论依据。
2 材料与方法
2.1 样本采集
本研究的实验动物中缅树鼩均为处于非繁殖期、且体重均无显著差异的实验室繁殖个体(由中国医学科学院医学生物学研究所提供)。实验前动物均独立饲养,实验或者观察时尽量避开喂食、进食、休息和活动的高峰期。
2.2 社群行为的观察
采用由三个相邻的独立箱(规格为:400 mm×400mm×550 mm,材质为透明亚克力和不透明亚克力)构成并装有实时监控摄像头的竞技场进行观察。饲养房内装有15 W暖色白炽灯作为行为观察时的照明。
配对个体间行为观察:实验前将实验动物分别放于竞技场三个相邻观察箱两端,适应15~20 min后,将中间不透明隔板换为透明隔板,并打开位于中间观察箱两侧的活动门,两侧原来被隔离的个体处于一个联通的大空间内,监测观察10 min(实验时间为10:00~16:00)。以20 s为1个观察时间单位,某种行为在1个时间单位内出现便计为1。最后计算在10 min内该行为出現的总数。实验动物组合为中缅树鼩雌性和雌性、雄性和雄性、雌性和雄性,实验所用动物实验后均隔一周再进行下次实验。实验结束后,用清水和75%的酒精擦洗竞技场,以备下次实验使用。
多个(1雌性2雄性、1雄性2雌性)个体间行为观察:实验前将实验动物分别放于竞技场三个相邻观察箱,适应15~20 min后,将中间不透明隔板换为透明隔板,并打开位于中间的观察箱两侧的活动门,监测观察24 h,记录各个体间出现的行为,实验中每个实验动物在各个观察箱内位置各放一次,即每组实验重复3次,每次实验间隔7d。实验所用动物在实验后均隔一周再进行下次实验。实验结束后,用清水和75%的酒精擦洗竞技场观察箱,以待下次实验。优势中缅树鼩是指在两个个体争斗中,防御行为较少而攻击行为明显多且尾巴一直翘着的个体,反之则为弱势中缅树鼩。
2.3 数据处理
实验数据均采用SPSS 19.0统计软件包统计分析处理。行为统计均使用独立样本T检验( independent- samples T test)进行分析。结果均使用平均值±标准误( Mean±SE)表示,P<0.05表示差异显著,P<0.01表示差异极显著。
3 结果
3.1 配对个体间行为观察
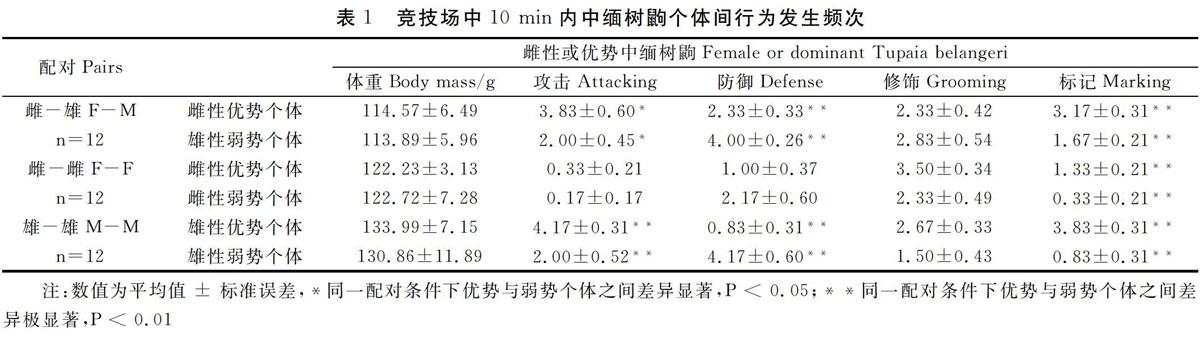
中缅树鼩异性个体之间,雌性个体的攻击行为显著高于雄性个体(t=2.45,PO.05,表1)。
中缅树鼩同性个体之间,两个雌性个体间除了标记行为优势个体极显著高于弱势个体(t= -3.35,P<0.01),攻击行为(t=0.62,P>O.05)、防御行为(t--1.66,P>O.05)和修饰行为(t=0.24,P>O.05)差异均不显著;两个雄性个体之间,优势个体攻击行为(t-3. 61,PO.05)差异不显著(表1)。
3.2 多个个体间行为观察
1雌性2雄性间攻击行为多发生在2个雄性之间,2雌性1雄性间攻击行为多发生在雄性与雌性之间。1雌性2雄性:初始时各个体均进入各自所在观察箱内的暗盒,而后出来四处窜跳,并在自己的观察箱内四处嗅探观察,尾部高翘,进行尿液标记并伴有粪便标记,少许个体用舌头舔舐观察箱和暗盒。雌性个体一直处于初始观察箱内,活动片刻便进入暗盒中;雄性个体通过隔板活动门进入到相邻观察箱内,然后又回到初始观察箱,持续几次后两个雄性个体均留在雌性个体所在的初始观察箱内并开始标记,二者开始相互攻击,雌性个体不参与攻击,两雄性中优势个体(争斗中攻击行为多)主动攻击对方,弱势者逃离到相邻观察箱。雌性个体存在时两雄性个体攻击力度和攻击频次远远强于没有雌性个体时。争斗结束后,大多数雌性个体选择争斗获胜的雄性。
1雄性2雌性:刚进入观察箱内先进入暗盒,一段时间后均钻出暗盒开始到处窜跳进行标记,随后通过活动门进入其他个体所在观察箱,到处嗅探后便立即返回,如此往复几次。雄性进入雌性个体领地后,首先进入暗盒,而后对雌性个体进行嗅闻,四处观看并不断修饰,修饰时会舔舐自己的阴部,再嗅舔雌性的阴部,如果雌性屡次拒绝,雄性就会展开攻击,雄性最终选择一个雌性个体并长期占据该雌性所在观察箱的暗盒,该雌性便与另一雌性个体到其他观察箱中,且两个雌性个体间没有出现明显的争斗行为。当雌性回到自己观察箱时,雄性将其赶出,重复几次后,雄性开始追逐雌性并进行交配。如若另一雌性进入领地,雄性立即攻击,赶走对方。
1雌性2雄性组合中配对成功的雌雄性个体再挑1雌性个体组合,成为新的2雌1雄组合,放入竞技场观察箱进行行为监测,发现无论将原来的雌性个体放在相邻还是相隔观察箱,雄性个体总会优先选择该雌性。1雄性2雌性组合中配对成功的雌雄性个体再挑1雄性个体组合,成为新的2雄1雌组合,放人竞技场观察箱进行行为监测,发现3个个体通过各自标记相互嗅闻后,雌性个体与雄性个体之间无攻击行为发生,两个雄性个体之间出现严重的争斗攻击行为,且雌性亦会优先选择先前交配成功的雄性。
4 讨论与结论
近年来,动物行为的研究逐渐进入国内外学者的视线,成为当下研究的热点问题,如对啮齿类动物和大型兽类社群行为、繁殖行为、攻击行为的研究[1]。本研究结果表明,中缅树鼩的标记行为均有产生,这说明尿液和唾液中相关的信息素成分可能是引起中缅树鼩不同行为诸如攻击、交配行为产生的原因。
社会等级是动物社群结构的表现形式,对于动物的社群及其相关行为的研究已有一些报道[10],社会等级在社群的建立和维持上起着重要作用[11]。已有研究报道,雌性树鼩具有极强的忍耐性,无论是幼仔还是同性均能友好相处。然而雄性之间的忍耐性极低,故雄性树鼩之间通常均会发生争斗攻击行为[12],本研究的观察结果与上述结论一致。在野外,中缅树鼩大多喜欢单独活动,只有在交配季节才能见到它们成对相互追逐[13],在本研究中,大多数雌性树鼩会选择争斗中占优势的雄性,说明雌性在选择配偶时可能和雄性的适合度有关。雄性或雌性均能优先识别出之前与之配对的树嗣,这可能与气味识别等有关;成年雌性间的友好相处和成年雄性间攻击行为较成年雌雄个体之间增加,符合中缅树鼩单雄配偶系社会结构[14]。
参考文献:
[1]张君,胡锦矗,行为生态学在中国的研究与进展[J].西华师范大学学报(自然科学版),2003,24(3):325~329.
[2]张建军,施大钊,不同社群条件下雄性布氏田鼠的行为[J].兽类学报,2006,26(2):159~163.
[3]张建军,施大钊,雄性等级对布氏田鼠雌性选择的影响[J].生态学杂志,2007,26(2):219—222.
[4]陈燃,李进华,朱勇,等,黄山短尾猴不同顺位等级雄性个体友好行为的比较研究[J].兽类学报,2009,29(3):246—251.
[5]聂海燕,刘季科.根田鼠攻击行为模式及其进化稳定对策分析[J].生态学报,2004,24(7):1406~1412.
[6]张健旭,張知彬,大仓鼠在繁殖期的行为关系及交配行为[J].兽类学报,1999,19(2):132—142.
[7]王刚,王传干,王振龙,等,黑线仓鼠殴斗行为模式及其与生理状态的关系[J].兽类学报,2009,29(3):286~293.
[8]王晓娟,杨春,苏建家.树鼩在医学实验研究中的新进展[J].中国比较医学杂志,2010,20(2):67~70.
[9]Xu X, Chen H, Cao X, et al. Efficient infection of tree shrew(Tupaia belangeri) with hepatitis C virus grown in cell culture orfrom patient plasma[J]. Journal of General Virology, 2007,88(9) :2504~2512.
[10]Premnath S, Sinha A, Gadagkar R.Dominance relationships inthe establishment of reproductive division of labor in a primitivelyeusocial wasp (Ropalidia marginata)[J]. Behavioral Ecology andSociobiology, 1996,39 (2):125~132.
[lllPiper W H. Social dominance in birds[J]. Current Ornithology,1997,14(1) :125~187。
[12]Yao Y G. Creating animal models, why not use the Chinese treeshrew (Tupaia belangeri chinensis)?[J]. Zoological Reasearch,2017,38(3) :118~126.
[13]王应祥,李崇云,马世来.树鼩的分类与生态.树鼩生物学[M].昆明:云南科学技术出版社,1991.
[14]Shadbolt A B.2015. Behaviour of the painted treeshrew (Tupaiapicta) in conservation set - asides of the planted forest zone ofSarawak, Malaysia[J]. Malayan Nature Journal, 2014( 664):420~439.
猜你喜欢
销售与市场·管理版(2021年9期)2021-08-30
时代英语·高二(2021年4期)2021-07-29
汉语世界(The World of Chinese)(2019年6期)2019-09-10
青年时代(2019年6期)2019-04-04
儿童故事画报·智力大王(2019年9期)2019-02-02
畅谈(2018年16期)2018-09-29
凤凰生活(2018年9期)2018-09-17
中国房地产·市场版(2017年10期)2018-01-15
足球周刊(2016年17期)2016-11-10
儿童故事画报·智力大王(2016年2期)2016-04-16
