
扬州地区烟粉虱生物型检测及发生动态
2018-01-29 00:22刘晓娜杜以梅杜予州
江苏农业科学 2017年23期
关键词:扬州市
刘晓娜+杜以梅+杜予州

摘要:连续2年检测扬州地区烟粉虱生物型,并对其危害程度和发生动态进行系统调查,以期为扬州地区烟粉虱的防治提供依据。利用细胞色素氧化酶I(cytochrome axidase I,简称COI)分子标记检测扬州地区烟粉虱的生物型,采用定时定点的方法调查烟粉虱的危害程度和发生动态。结果表明,2014年和2015年扬州地区发生的烟粉虱均为Q型;烟粉虱从6月初到11月初在主要蔬菜上的危害等级一直保持在4级,在其他时间的危害程度不高。2014年扬州地区大棚蔬菜上的烟粉虱始见于4月上旬,消亡期为12月下旬,在7月下旬有1个危害高峰;2015年烟粉虱始见期为4月中下旬,到11月底仍有烟粉虱危害,其间有2个危害高峰,分别在5月底至6月初和7月底至8月中旬。2014年扬州地区烟粉虱的发生危害及虫口密度高于2015年,其发生的虫口密度在不同年份之间有波动,而且在不同的月份之间也存在差异。
关键词:烟粉虱;生物型;危害程度;发生动态;扬州市
中图分类号: S433.39文献标志码: A文章编号:1002-1302(2017)23-0100-03
该虫为多食性害虫,寄主范围广,寄主适应性强,在热带、亚热带和温带地区一年发生11~15代,具有世代重叠的特性。烟粉虱可以在温暖地区的野外杂草和花卉上越冬,但在寒冷地区的烟粉虱则主要在保护地作物上越冬。烟粉虱是一类快速进化的复合种,其中B型烟粉虱(也称Middle East-Asia 1)和Q型烟粉虱(Mediterranean)在全球范围内危害较为严重,入侵性也最强[1-2]。2001年在扬州地区首次发现B型烟粉虱,随后在江苏地区迅速扩散危害,造成严重的经济损失[3]。但2005年在江苏省发现Q型烟粉虱后,Q型烟粉虱的种群数量逐年增加,而B型烟粉虱的种群数量却逐年递减,到2010年Q型烟粉虱已成为优势种[4]。孙伟等连续多年对江苏省范围内烟粉虱的发生危害进行调查发现,烟粉虱的发生危害不仅在不同年份之间存在波动,而且在不同地区之间也存在着显著差异[5-6]。为进一步了解和掌握扬州地区烟粉虱生物型及全年的发生危害动态,于2014、2015年对扬州不同地区烟粉虱的生物型进行检测,并以扬州蒋王蔬菜基地作为系统调查点,对烟粉虱在其主要嗜好寄主植物上的发生危害动态做详细的调查,以期为扬州地区烟粉虱综合防治提供科学依据。
1材料与方法
1.1扬州地区烟粉虱生物型检测
1.1.1材料供试虫源:于2014、2015年在扬州市邗江区、广陵区、江都区、高邮市、仪征市、宝应县的番茄(Lycopersicon esculentum)、黄瓜(Cucumis sativus)、辣椒(Capsicum annuum)、茄子(Solanum melongena)等主要蔬菜和过渡寄主葎草(Humulus scandens)上采集烟粉虱成虫。将采集到的样本浸泡于装有无水乙醇的1.5 mL Eppendorf管中,于-20 ℃条件下保存。
主要试剂有蛋白酶K、rTaq DNA聚合酶、10×Buffer,购自宝生物工程(大连)有限公司;dNTPs,购自中美泰和生物技术(北京)有限公司。
1.1.2方法采用细胞色素氧化酶I(cytochrome axidase I,简称COI)分子标记快速检测法,该方法引用福建省农业科学院植物保护研究所申请的专利201110413571《烟粉虱生物B型和Q型特异性引物和快速鉴定方法》。这个专利主要针对B型和Q型分别设计了2对特异性引物,构建了一套适合烟粉虱生物型快速鉴定的双引物PCR扩增体系。该方法可以快速鉴定田间外来入侵烟粉虱的生物型。若该方法不能检测出烟粉虱样本的生物型,笔者采用通用引物,即上游引物 C1-J-2195(5′-TTGATTTTTTGGTCATCCAGAAGT-3′)和下游引物L2-N-3014(5′-TCCAATGCACTAATCTGCCATA
TTA-3′)对烟粉虱mtDNA COI基因进行扩增并送中美泰和生物技术有限公司测序,将测序结果进行Blast比对分析,确定其生物型。本研究共检测扬州地区288个样本。
1.2扬州地区烟粉虱危害程度及发生动态调查
于每年2月开始定期对扬州市蒋王蔬菜基地的番茄、茄子、辣椒进行系统调查,每7 d调查1次。对不同的寄主均采用“Z”字形5点采样,每点随机调查5株,每株分别摘取上、中、下3张叶片,将不同寄主的叶片放入不同的袋子中带回实验室进行镜检,记录每张叶片中烟粉虱卵、若虫和伪蛹的数量以及叶面积,然后折算成单位叶面积的虫口密度并划分危害等级。具体方法:(1)将烟粉虱的危害程度划分为5级。(2)设定1个标准叶面积为10 cm2。调查时,将调查叶片的平均叶面积换算为标准叶面积。(3)以1个标准叶面积上的平均烟粉虱数量(卵、若虫和伪蛹的总数量)为分级单位,即虫量(头)/标准叶面积。(4)分级标准[平均虫量(头)/标准叶面积(10 cm2)]:0级为无虫,1级≤1头/10 cm2,2级为2~3头/10 cm2,3级为 4~6头/10 cm2,4级≥7头/10 cm2,分别记为“-”“+”“++”“+++”“++++”。
2结果与分析
2.1扬州地区烟粉虱生物型检测
連续2年对扬州市邗江区、广陵区、江都区、高邮市、仪征市、宝应县蔬菜基地不同寄主植物上的烟粉虱进行生物型检测。结果表明,扬州地区主要蔬菜和过渡寄主葎草上的烟粉虱均为Q型烟粉虱,没有发现B型烟粉虱(表1)。这表明扬州地区主要蔬菜上的B型烟粉虱已被Q型所取代。
2.2扬州地区烟粉虱的发生危害程度
2014年的调查数据表明,扬州地区烟粉虱4月初开始在番茄上出现,至5月初危害较轻,危害程度一直保持在1级;5月中旬危害开始逐渐加重,危害程度上升至2级,5月下旬后危害程度升至4级,其危害在7月中旬达到高峰,与蔡力等2013年的调查结果[7]相比提前40 d左右,此间主要种植的蔬菜有番茄、茄子和黄瓜等;从5月下旬至10月底,烟粉虱在番茄、茄子、黄瓜等主要蔬菜上的危害程度一直维持在4级水平;从11月初开始,烟粉虱的危害程度开始下降,此时已开始种植下一季番茄,11月10日左右危害程度降为3级,到11月末烟粉虱的危害程度降到1级,直到12月中旬调查结束。endprint
2015年的调查数据表明,烟粉虱从4月中下旬开始在番茄上发生,较2014年发生晚,直到5月中旬,危害程度一直处在1级;5月中下旬危害程度开始上升至2级,此时寄主蔬菜主要以番茄为主;从5月下旬开始,烟粉虱的危害程度迅速上升至4级,直到10月初,此时除了种植番茄以外还有黄瓜、茄子等蔬菜;从10月中旬开始危害程度降为3级,此时大棚里主要种植的是茄子、豇豆等蔬菜;此后到12月初,烟粉虱主要转至十字花科蔬菜危害,其危害程度较轻,危害级别为1级。
从扬州地区2014年和2015年的调查数据可以看出,烟粉虱在番茄、茄子、黄瓜等主要蔬菜上的危害程度趋势基本一致,但2015年较2014年发生晚,发生危害程度略轻。
2.3扬州地区烟粉虱的发生动态
2014年的调查结果表明,从4月3日发现烟粉虱到5月中下旬,在叶片上的虫口密度一直很低,为0.03~0.15头/cm2;从5月底开始,烟粉虱的虫口密度开始增加,为 0.83头/cm2 左右;在6月20日至7月24日的1个月时间里,虫口密度由4.09头/cm2迅速增长为30.68头/cm2,达到危害最高峰,此间田间主要种植茄子;随后虫口密度迅速下降,到8月9日虫口密度降至3.64头/cm2;此后烟粉虱的虫口密度除了在10月27日有小幅度的增长外,一直维持在较低水平,为0~0.59头/cm2,直到调查结束(图1)。
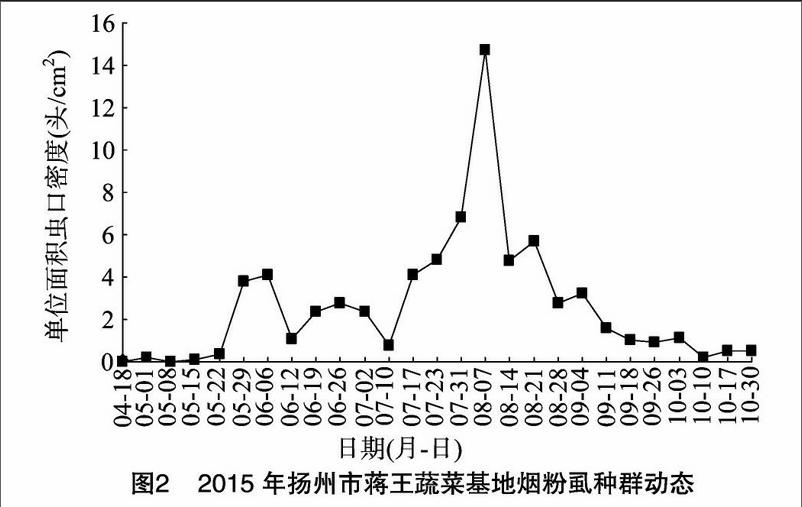
2015年的调查数据表明,4月18日开始发现烟粉虱危害番茄,直到5月22日,叶片上的虫口密度一直不高,在 0.02~0.36头/cm2之间;到5月下旬至6月初,发生危害形成1个小高峰,其虫口密度在6月6日达到4.11头/cm2,此时田间主要种植番茄;6月中旬番茄收获,主要寄主更换为黄瓜,虫口密度有所下降,6月12日的虫口密度为 1.08头/cm2。随后20 d左右的时间,虫口密度没有大幅变化,在2.35~2.77头/cm2;7月10日虫口密度达到1个月之内的最小值为0.75头/cm2;7月中旬以后,烟粉虱的危害加重,虫口密度快速上升,8月7日达到最高峰,虫口密度为1472头/cm2;随后,虫口密度迅速下降,到8月28日虫口密度降到2.77头/cm2,之后的虫口密度逐渐下降,此间烟粉虱的寄主蔬菜主要是茄子;在10月30日茄子收获后,大多数烟粉虱转到在十字花科蔬菜上危害,但数量很少(图2)。
从调查的数据可以看出,2014年扬州地区烟粉虱危害高峰期的虫口密度明显高于2015年的,是2015年的2倍多;2014年烟粉虱只出现了1个明显的危害高峰,而2015年则出现了2个危害高峰,1个危害高峰期在5月底至6月初,另1个危害高峰期在7月底至8月中旬, 出现的时间和2014年
基本一致。
3讨论
2001年,在扬州仪征花卉生产基地的一品红上首次发现B型烟粉虱,这也是江苏省的首例[3],随后其不断的扩散,给农业生产造成了严重的损失。2005年江苏省检测到Q型烟粉虱,此后Q型烟粉虱的种群数量逐年递增,而B型烟粉虱却逐年递减[4]。2008—2013年扬州地区都未检测到B型烟粉虱[7],2014—2015年的检测结果也同样如此。短短几年的时间,扬州地区B型烟粉虱完全被Q型烟粉虱所取代,可能的原因有:(1)Q型烟粉虱有更强的寄主适应性,杂草是Q型烟粉虱的主要寄主,这一点在调查中也有体现[8-9];(2)有研究报道,Q型烟粉虱对新烟碱类农药有更强的抗性[10];(3)可能与扬州地区蔬菜种植布局及品种有关。由于目前扬州地区烟粉虱都为Q型,它对新烟碱类农药更有抗性,所以在对其进行化学防治时,应注意不同药剂之间合理的混用轮用,以延缓抗性产生。
调查结果表明,2014、2015年扬州地区烟粉虱的危害动态大体一致,5月中旬之前危害程度一直为1级,从6月初到11月初一直保持4级的危害程度,这与孙伟等的调查结果[5,11]一致。通过2年的种群动态调查可以看出,2014年扬州地区烟粉虱危害高峰期的虫口密度明显高于2015年,是2015年的2倍多;2014年在扬州蒋王蔬菜基地的烟粉虱只出现了1个危害高峰,而2015年则出现了2个危害高峰,这可能与扬州地区这2年的天气(温度、湿度、降水和风等)和寄主蔬菜的生长状况等因素有关。因为温度、湿度可以影响烟粉虱的生长发育、存活和繁殖[12-13],也可以影响寄主蔬菜的生长状况,从而影响烟粉虱的发生动态。而2014年扬州地区早春的平均气温高于2015年,有利于烟粉虱的发生危害。扬州地区2015年的降水量明显高于2014年,且在4月中旬、5月中旬至7月中旬以及8月上旬出现多个持续降水天气,这一天气条件不利于烟粉虱发生危害。此外,由于扬州蒋王蔬菜基地温室设施老旧,防雨、保温、保湿的效果较差,而2015年雨水较多,寄主蔬菜受涝害较重,长势较差。这些因素导致2014年烟粉虱的发生危害和虫口密度明显高于2015年。
值得注意的是,烟粉虱在-2 ℃以下时开始出现死亡,随着温度的下降和处理时间的延长,烟粉虱的死亡率迅速上升,-6 ℃时烟粉虱处理7 h以上全部死亡[12]。而在扬州地区,最低气温是要低于烟粉虱致死温度的,且持续时间较长,故在扬州地区烟粉虱只能在温室中越冬,不能在野外越冬。因此,在防治温室大棚内的烟粉虱时,要注意保持棚内空气流通,适当掀开大棚的薄膜以降低棚內温度,有效地抑制烟粉虱的生长繁殖。通过调查发现,在寄主轮换时烟粉虱数量骤减,而在蒋王地区作物的种植是没有间隔期的,这就导致烟粉虱种群数量骤减后又迅速增长。因此,在烟粉虱的防治工作中,要注意彻底清理残枝落叶,减少危害下一茬作物的虫源基数,适当调整作物种植间隔期。
参考文献:
[1]Dinsdale A,Cook L,Riginos C,et al. Refined global analysis of Bemisia tabaci (Hemiptera:Sternorrhyncha:Aleyrodoidea:Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries.[J]. Annals of the Entomological Society of America,2010,103(2):196-208.endprint
[2]Barro P D,Ahmed M Z. Genetic networking of the Bemisia tabaci cryptic species domplex reveals pattern of biological invasions[J]. PLoS ONE,2011,6(10):e25579.
[3]周福才,杜予州,陆自强. 江苏发现B型烟粉虱危害[J]. 扬州大学学报(自然科学版),2001,4(3):37.
[4]沈媛,杜予州,任顺祥,等. 江苏地区烟粉虱生物型演替研究初报[J]. 应用昆虫学报,2011,48(1):16-21.
[5]孫伟,杜予州,沈媛,等. 江苏扬州地区蔬菜烟粉虱寄主调查及种群动态[J]. 华东昆虫学报,2005,14(1):38-43.
[6]蔡力,许丽丽,杜以梅,等. 江苏地区烟粉虱发生危害及其生物型监测[J]. 应用昆虫学报,2015,52(1):124-134.
[7]蔡力. 江苏地区烟粉虱生物型监测及其遗传多样性研究[D]. 扬州:扬州大学,2014:32-38.
[8]Iida H,Kitamura T,Honda K. Comparison of egg-hatching rate,survival rate and development time of the immature stage between B-and Q-biotypes of Bemisia tabaci (Gennadius) (Homoptera:Aleyrodidae) on various agricultural crops[J]. Applied Entomology and Zoology,2009,44(2):267-273.
[9]Tsueda H,Tsuchida K. Reproductive differences between Q and B whitefly,Bemisia tabaci,on three host plants and negative interactions in mixed cohorts[J]. Entomologia Experimentalis et Applicata,2011,141(3):197-207.
[10]Horowitz A R,Kontsedalov S,Khasdan V,et al. Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance[J]. Archives of Insect Biochemistry and Physiology,2005,58(4):216-225.
[11]赵伟. 山东烟田烟粉虱的发生动态及防治技术[D]. 北京:中国农业科学院,2014,48-54.
[12]崔红旭,郑丹,蒲桂林,等. 温度对B型、Q型烟粉虱生长发育、存活和繁殖的影响[J]. 植物保护学报,2009,36(6):571-572.
[13]陈夜江,罗宏伟,黄建,等. 湿度对烟粉虱实验种群的影响[J]. 华东昆虫学报,2001,10(2):76-80.江苏农业科学2017年第45卷第23期王呈玉,张浩,崔俊涛,等. 人参根际防病促生放线菌的筛选及其活性[J]. 江苏农业科学,2017,45(23):103-106.endprint
猜你喜欢
江苏安全生产(2023年9期)2023-10-24
江苏安全生产(2022年11期)2023-01-11
江苏安全生产(2022年9期)2022-11-02
江苏安全生产(2022年7期)2022-08-24
江苏安全生产(2022年5期)2022-06-16
江苏安全生产(2021年10期)2022-01-18
华人时刊(2021年23期)2021-03-08
江苏安全生产(2021年12期)2021-03-08
军民两用技术与产品(2020年6期)2020-07-18
大众考古(2015年5期)2015-06-26
