
北方白菜型冬油菜的膜脂脂肪酸组分和ATPase活性对温度的响应
2018-01-18 01:18孙万仓武军艳刘自刚何辉立
作物学报 2018年1期
方 彦 孙万仓,* 武军艳 刘自刚 董 云,3 米 超 马 骊 陈 奇 何辉立
北方白菜型冬油菜的膜脂脂肪酸组分和ATPase活性对温度的响应
方 彦1,2孙万仓1,2,*武军艳2刘自刚2董 云2,3米 超2马 骊2陈 奇2何辉立2
1甘肃省干旱生境作物学重点实验室, 甘肃兰州 730070;2甘肃省油菜工程技术研究中心, 甘肃兰州 730070;3甘肃省农业科学院作物研究所, 甘肃兰州 730070
为了解北方白菜型冬油菜膜脂脂肪酸和ATPase活性与抗寒性的关系, 以抗寒性强的冬油菜品种陇油7号和抗寒性弱的品种天油2号为材料, 研究了不同温度处理(25°C、10°C、2°C、–5°C)后叶片和根系膜脂脂肪酸组分和ATPase酶活性的变化。结果表明, 低温胁迫下2个冬油菜品种叶片和根系膜脂脂肪酸组分相同, 叶片中不饱和脂肪酸以亚麻酸为主, 根系不饱和脂肪酸以亚油酸为主。随处理温度的降低, 2个冬油菜品种叶片不饱和脂肪酸含量呈先降低(10°C, 2°C)后增加(–5°C)的趋势; 陇油7号根中不饱和脂肪酸含量逐步增加, 天油2号则逐步降低; 在低温条件下(2°C, –5°C), 陇油7号膜脂U/S比值、IUFA值高于天油2号; ATPase活性表现为陇油7号逐渐高于天油2号。说明2个冬油菜品种的膜脂在低温响应上存在一定差异, 低温下不饱和脂肪酸含量和ATPase活性的提高是强抗寒冬油菜品种在北方旱寒区严酷环境条件下能安全越冬的重要原因。
冬油菜; 脂肪酸; ATP酶活性; 抗寒性
植物细胞膜是细胞与外界环境联系的直接界面, 对环境变化极为敏感[1], 受到低温刺激时膜脂的组成、结构和状态等会发生改变[2-3]。当达到一定的温度条件时, 膜脂由流动液相转变为凝固的晶体固相[4-6], 由此引起膜透性增大, 细胞液外渗, 破坏细胞膜内外的酸碱平衡, 最终导致细胞死亡[7-8]。抗寒锻炼能提高植物的抗寒性, 抗寒锻炼中, 生物膜自身能够随环境温度的变化对膜脂的不饱和度进行适应性调整[9], 各膜脂成分的相对含量及其脂肪酸组成发生改变[10-12]。已有研究表明[13], 膜脂不饱和度的增加有利于维持细胞结构的稳定性, 使植物耐冷性提高。王萍等[14]研究表明脂肪酸不饱和度能够反映水稻品种对低温的适应性, 与其抗冷性一致。王洪春等[4]研究表明, 水稻种子干胚膜脂的脂肪酸不饱和度受胚形成期温度的影响, 抗冷水稻品种的脂肪酸不饱和指数高于冷敏感品种。王金胜等[15]研究证明温度变化能影响玉米幼苗线粒体膜脂脂肪酸的组成, 品种耐冷性越强, 脂肪酸不饱和指数越大。膜脂的流动性与膜结合酶密切相关[16]。植物在遭受低温冻害时, 细胞膜的离子主动运输体系是损伤的最直接部位, ATPase具有致电泵的作用, 在离子转运过程中具有重要的调控作用[17]。ATP依赖的离子类型有Ca2+/Mg2+-ATPase和K+/Na+-ATPase[18]。膜脂肪酸组成会影响膜结合酶活力[19], 膜结合酶活性的大小能够作为抗冷性强弱评估的指标[20]。刘炜[21]对低温下不同抗寒性小麦品种细胞内Ca2+的稳态平衡能力研究结果表明, 低温下质膜Ca2+-ATPase的活性大小及其稳定性是决定植物抗寒能力的关键因素。王红等[22]指出抗寒锻炼能使小麦幼苗在低温下保持较高的Ca2+-ATPase活性。王精明等[23]研究表明, 低温锻炼能提高冷胁迫下水稻幼苗质膜和液泡膜Mg2+- ATPase活性。
北方旱寒区环境严酷, 寒冬漫长, 极端低温低, 国内外引入的冬油菜抗寒品种种植在该区均难以越冬, 只有强冬性白菜型冬油菜适宜在该区种植[24]。研究表明[25], 秋末和初冬的低温锻炼能够诱导越冬植物产生耐寒性, 以适应越冬期间的低温。在这个适应的过程中, 植物体内发生一系列的生理生化变化, 包括细胞膜系统的变化、与植物抗寒性密切相关的抗氧化酶系统的变化等, 这些生理生化的变化是评定植物抗寒性的主要指标。本课题组前期曾对低温条件下冬油菜电导率、丙二醛、过氧化物酶类等膜脂稳定性相关指标分析比较[26-28]。目前未见有关冬油菜细胞膜脂脂肪酸组分和ATP酶活性在低温胁迫下与抗寒性关系的报道。本文研究了2个抗寒性不同的冬油菜品种膜脂脂肪酸组成及ATP酶活性在不同温度下的变化, 以期探讨北方白菜型冬油菜对气温变化的膜脂响应, 为阐明北方白菜型冬油菜的抗寒机理和抗寒性的早期鉴定提供理论依据。
1 材料与方法
1.1 试验材料
强抗寒性白菜型冬油菜品种陇油7号, 在我国北方寒区越冬率在80%以上; 弱抗寒性品种天油2号, 越冬率40%~80%[24]。种子由甘肃省油菜工程技术研究中心提供。
1.2 试验设计
采用盆栽试验, 将种子播于营养钵, 置人工气候箱培养。待幼苗长至5~6片真叶时放入温度试验箱, 于25°C (CK)、10°C、2°C和–5°C处理, 72 h后分别取叶片和根, 一部分用液氮迅速冷冻后, 锡纸包裹放入–80°C超低温冰柜用于ATP酶活性测定, 一部分105°C 5 min钝化脂酶, 剪碎混匀, 用于膜脂提取。设3次重复试验。
1.3 膜脂提取
参照文献[29]方法并改进提取膜脂。称取高温热杀钝化脂酶的混匀样品100 mg, 研磨后移入15 mL离心管, 加5%盐酸甲醇溶液2 mL, 氯仿甲醇溶液(体积比1∶1) 3 mL, 100 µL十九烷酸甲酯内标, 置85°C水浴锅中1 h, 待温度降到室温后在离心管中加入1 mL正己烷, 振荡萃取 2 min, 静置1 h等待分层。取上层清液100 µL, 用正己烷定容到1 mL, 0.45 µm滤膜过滤后上机测试。
1.4 膜脂中脂肪酸成分分析
采用ThermoFisher Trace 1310 ISQ气相色谱质谱联用仪分析脂肪酸成分, 柱温为: 80°C保持1 min, 以10°C min–1的速率升温至200°C, 继续以5°C min–1的速率升温至250°C, 最后以2°C min–1的速率升到270°C, 保持3 min, 进样口温度为290°C, 采取不分流进样, 重复3次。根据样品保留时间定性分析, 确定样品中脂肪酸的种类, 然后按照峰面积归一法计算求得各脂肪酸在总脂肪酸中的相对含量, 并计算不饱和脂肪酸含量/饱和脂肪酸含量(U/S)与膜脂脂肪酸不饱和指数(IUFA)。
U/S=[(16:1)+(18:2)+(18:3)]/[(14:0)+(15:0)+(16:0) +(17:0)+(18:0)]
IUFA=[(16:1)+(18:2)×2 + (18:3)×3]×100%
式中, 小括号中的数值代表不同的脂肪酸种类, 16:1为棕榈油酸, 18:2为亚油酸, 18:3为亚麻酸, 14:0为肉豆蔻酸, 15:0为十五烷酸, 16:0为棕榈酸, 17:0为十七烷酸, 18:0为硬脂酸。
1.5 Ca2+/Mg2+-ATPase和K+/Na+-ATPase活性的测定
将冷冻样品在预冷的0.05 mol L–1Tris-HCl, pH 7.4磷酸盐缓冲液(PBS)中漂洗, 以滤纸拭干后准确称取0.5 g, 放入5 mL匀浆管, 以1∶4 (g mL–1)加入预冷的匀浆介质PBS冰浴充分研碎, 制成20%的匀浆液, 以13 000×4°C冷冻离心15 min, 重复此步骤2次, 取上清液备用。采用南京建成生物研究所生产的ATPase试剂盒按其说明测定ATPase活性。
1.6 统计分析
采用Microsoft Excel 2007处理图形, 用SPSS 19软件统计分析试验数据。
2 结果与分析
2.1 不同温度环境下冬油菜叶片形态的变化
当温度在零度以上时, 2个冬油菜品种均生长正常, 叶片深绿色, 没有受冻害的症状; 但在–5°C处理后, 表现出不同的受害症状, 陇油7号受害较轻, 仅表现出叶色变浅、下部叶片叶尖、叶缘卷曲等较轻微受害症状, 而天油2号出现严重受冻害症状, 叶缘呈水渍状冻斑(图1)。

图1 不同温度处理后冬油菜幼苗形态的变化
2.2 不同温度处理下冬油菜膜脂脂肪酸成分分析
不同温度处理下冬油菜叶片和根系膜脂脂肪酸组分相同(表1), 主要有十五烷酸(C15:0)、棕榈酸(16:0)、硬脂酸(18:0)、亚油酸(18:2)和亚麻酸(18:3), 约占总脂肪酸的95%~99%, 还有少量的肉豆蔻酸(C14:0)、十七烷酸(C17:0)和棕榈油酸(C16:1), 占1%~5%。说明十五烷酸、棕榈酸、硬脂酸、亚油酸、亚麻酸5种脂肪酸在冬油菜膜脂的组成上以及膜对外界温度发生变化时适应性方面比其他脂肪酸发挥着更为重要的作用。其中叶片中不饱和脂肪酸以亚麻酸为主, 约占总脂肪酸含量的14.33%~ 29.64%; 根系不饱和脂肪酸主要为亚油酸, 约占总脂肪酸含量的11.55%~16.20%。叶片和根系饱和脂肪酸均以棕榈酸为主, 约占总脂肪酸含量的36.69%~69.77%。

表1 不同温度处理下冬油菜脂肪酸组分及其相对含量
2.3 不同温度环境下冬油菜膜脂脂肪酸成分含量的变化
2.3.1 饱和脂肪酸含量的变化 由表2可以看出, 随着处理温度的降低, 2个冬油菜品种根和叶片中十五烷酸含量逐渐降低, 棕榈酸含量逐渐增加, 叶片中硬脂酸含量先上升后下降, 根中硬脂酸呈逐渐降低趋势。同一温度处理下, 根系饱和脂肪酸含量高于叶片。当温度较高时(25°C、10°C) 2个品种根系和叶片中饱和脂肪酸含量相差不大, 但在低温(–5°C)处理下, 天油2号叶片和根系膜脂饱和脂肪酸相对含量均高于陇油7号。
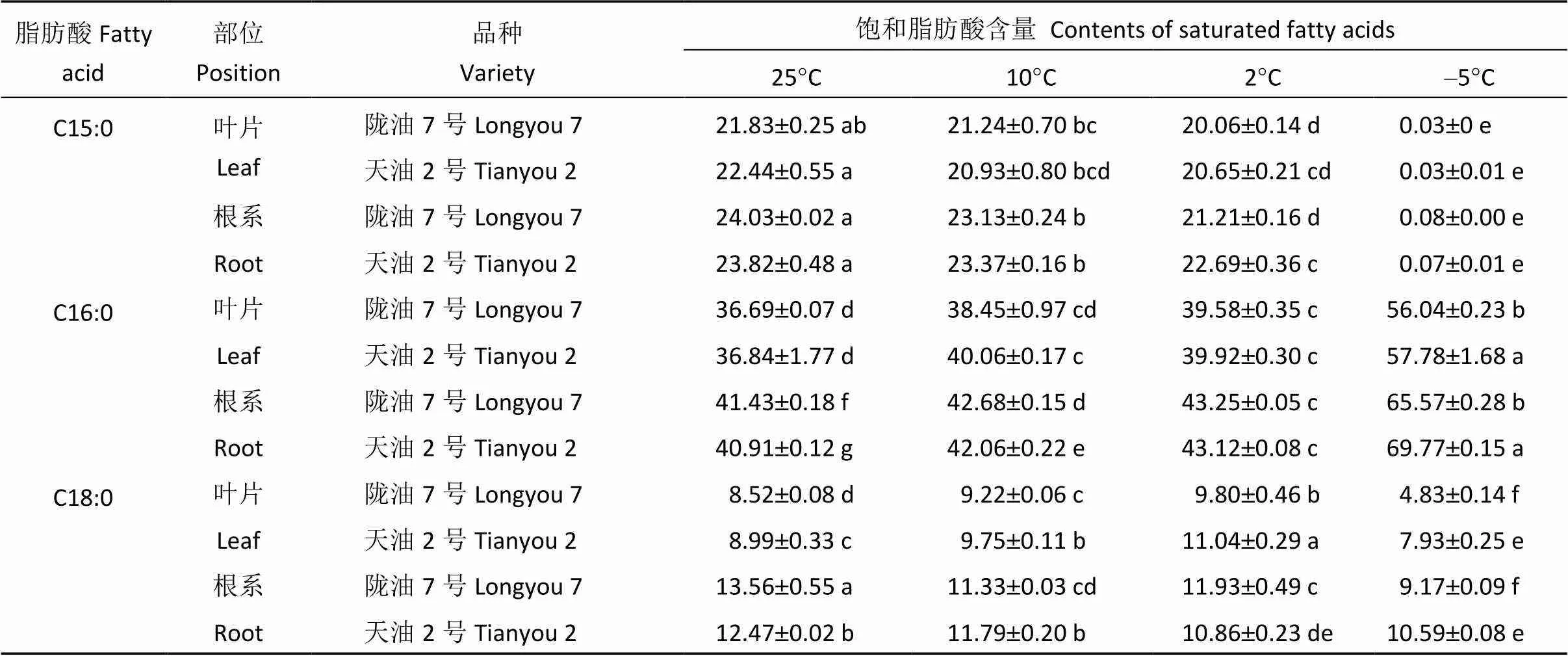
表2 不同温度下冬油菜叶片和根系饱和脂肪酸含量
表中小写字母表示2个品种同一部位的同一种脂肪酸含量在不同温度间的差异显著性(<0.05)。
Different small letters in the table show significant difference at the 0.05 level between the same position of Longyou 7 and Tianyou 2 of same fatty acids under different temperatures.
2.3.2 不饱和脂肪酸含量的变化 随着温度降低, 2个冬油菜品种叶片中亚油酸含量逐渐降低(表3), 亚麻酸含量逐渐上升。根系亚油酸含量, 在处理温度大于0°C时逐步增加, 温度低于0°C时显著降低, 天油2号降低幅度大于陇油7号; 降温过程中, 陇油7号根系亚麻酸含量逐渐上升, 天油2号呈逐渐降低趋势。温度小于0°C时, 陇油7号根系不饱和脂肪酸含量显著高于天油2号。
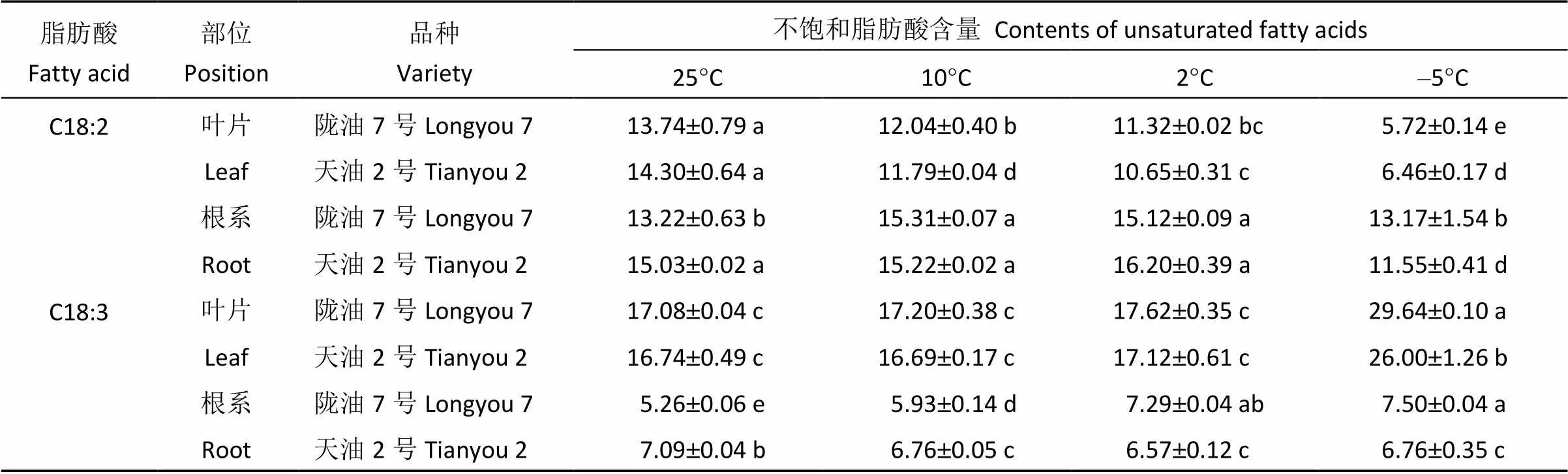
表3 不同温度下冬油菜叶片和根系不饱和脂肪酸含量
表中小写字母表示2个品种同一部位的同一种脂肪酸含量在不同温度间的差异显著性(<0.05)。
Different small letters in the table show significant difference at the 0.05 level between the same position of Longyou 7 and Tianyou 2 of same fatty acids under different temperatures.
2.3.3 不同温度环境下冬油菜脂肪酸不饱和程度的变化 由图2可知, 随温度的下降, 2个冬油菜品种叶片中的U/S比值呈先下降后增加的趋势。陇油7号叶片的U/S比值始终高于天油2号; 品种的根系U/S比值表现出较大变化, 随温度的下降, 陇油7号逐渐增加, 而天油2号在温度较高时(25°C、10°C)基本没有变化, 当温度降低到2°C时急剧下降; 降温过程中, 陇油7号根系U/S比值逐渐高于天油2号, 说明在降温过程中, 陇油7号叶片和根系能够通过调整脂肪酸相对含量, 积累相对较多的不饱和脂肪酸适应降温对其造成的伤害, 而天油2号根系在低温条件下不饱和脂肪酸合成受阻。
随着温度的降低, 2个品种IUFA值均呈逐步上升的趋势。温度较高时(25°C、10°C), 2个品种IUFA值间差异不显著; 低温条件下(2°C)时, 两品种间IUFA值差异明显; 胁迫温度小于0°C时, 陇油7号IUFA值显著高于天油2号(图3)。
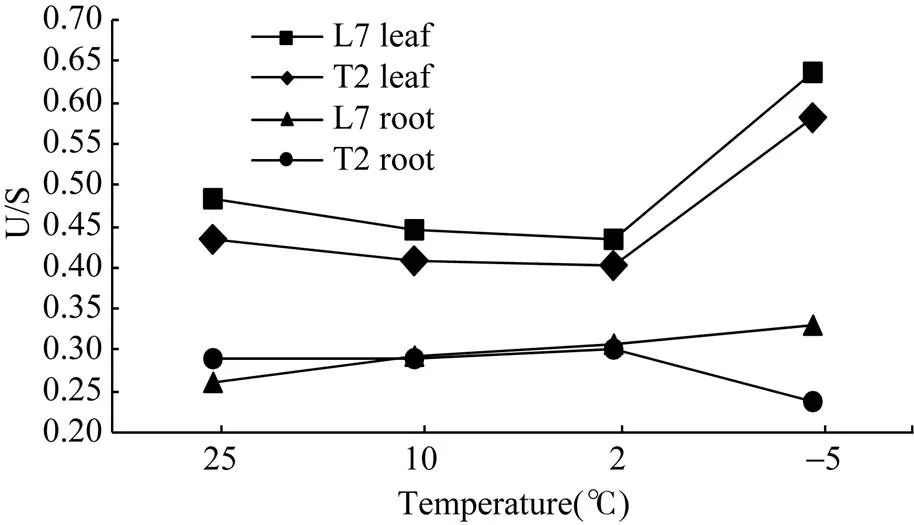
图2 不同温度下冬油菜U/S比值的变化
L7: 陇油7号; T2: 天油2号。L7: Longyou 7; T2: Tianyou 2.

图3 不同温度下冬油菜IUFA的变化
柱形图上不同字母表示2个品种在处理间存在显著性差异(< 0.05)。
Different letters above the bar mean significant difference between Longyou 7 (L7) and Tianyou 2 (T2) (< 0.05).
2.4 温度对冬油菜ATPase活性的影响
2.4.1 温度对冬油菜Ca2+/Mg2+-ATPase活性的影响
叶片与根中Ca2+/Mg2+-ATPase活性变化不同(图4和图5)。随着温度的降低, 叶片中Ca2+/Mg2+- ATPase活性由高到低, 而根中则是由低到高。在温度较高时(25°C、10°C), 叶片中的Ca2+/Mg2+-ATPase活性大于根系, 低温时(2°C、–5°C), 根系Ca2+/Mg2+- ATPase活性大于叶片。品种间叶片与根中Ca2+/ Mg2+-ATPase活性变化差异较大, 当温度在10°C以上时, 天油2号叶片与根中Ca2+/Mg2+-ATPase活性均高于陇油7号, 温度降至2°C以后, 陇油7号叶片与根中Ca2+/Mg2+-ATPase活性大于天油2号。这是由于温度较高时(即适宜温度时), 天油2号生长发育快, 而当温度降低到非适宜生长时, 天油2号地上部生长受阻, 适应性降低, Ca2+/Mg2+-ATPase活性降低。陇油7号则相反, 适宜温度时, 生长发育缓慢, 在温度下降到非适宜条件时, 其地下部生长正常, 适应性强。说明强抗寒的冬油菜在低温抗寒锻炼过程中, 质膜ATP酶获得了耐低温的特性, 以致在零度以下, 能保持更高的活性反应。

图4 不同温度下冬油菜叶片Ca2+/ Mg2+-ATPase活性
柱形图上不同字母表示2个品种在处理间存在显著性差异(< 0.05)。
Different letters above the bar mean significant difference between Longyou 7 (L7) and Tianyou 2 (T2) (< 0.05).
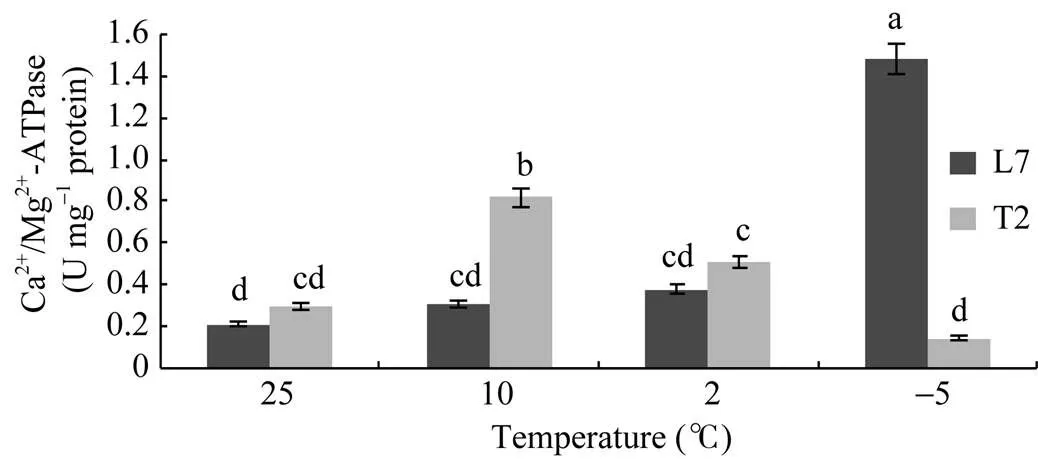
图5 不同温度下冬油菜根系Ca2+/Mg2+-ATPase活性
柱形图上不同字母表示2个品种在处理间存在显著性差异(< 0.05)。
Different letters above the bar mean significant difference between Longyou 7 (L7) and Tianyou 2 (T2) (< 0.05).
2.4.2 温度对冬油菜K+/Na+-ATPase活性的影响
陇油7号叶片K+/Na+-ATPase活性随着温度的降低而减小, 温度为–5°C时, K+/Na+-ATPase活性比常温(25°C)降低了77.4% (图6); 天油2号叶片K+/Na+- ATPase活性随着温度降低呈“升降升”的趋势, 在胁迫温度下均高于常温(25°C)。

图6 不同温度下冬油菜叶片K+/Na+-ATPase活性
柱形图上不同字母表示2个品种在处理间存在显著性差异(< 0.05)。
Different letters above the bar mean significant difference between Longyou 7 (L7) and Tianyou 2 (T2) (< 0.05).
随着温度降低, 陇油7号根系K+/Na+-ATPase活性逐渐提高(图7), 天油2号根系K+/Na+-ATPase活性逐渐降低, 当温度降至–5°C时, 2个冬油菜品种根系K+/Na+-ATPase活性差异显著, 陇油7号为天油2号的4.92倍。
3 讨论
3.1 质膜脂肪酸饱和性影响北方冬油菜抗寒性
质膜不饱和脂肪酸含量与饱和脂肪酸含量比值与植物抗寒性密切相关[30]。低温胁迫可使不同植物质膜脂肪酸组成比例改变, 不饱和脂肪酸含量增加[31-32]。膜脂中不饱和脂肪酸含量的增加能降低膜脂的相变温度, 增加膜脂的流动性, 使膜脂在低温下保持稳定, 以此提高植物的抗寒性[33-34]。膜脂脂肪酸不饱和指数( IUFA)是细胞膜中不饱和脂肪酸含量的总和,是衡量膜脂脂肪酸不饱和程度的重要指标之一, 可用来评价植物的抗寒性[35-36]。本研究在低温条件下(2°C, –5°C), 抗寒性强的陇油7号膜脂不饱和脂肪酸含量/饱和脂肪酸含量比值、IUFA值始终高于抗寒性弱的天油2号, 说明陇油7号在降温过程中能随温度变化对膜脂脂肪酸组分含量进行适应性调整, 低温下脂肪酸去饱和化程度强于天油2号, 能积累较多的不饱和脂肪酸, 增强细胞膜体系的稳定性, 可见, 质膜脂肪酸因品种抗寒性而异。

图7 不同温度下冬油菜根系K+/Na+-ATPase活性
柱形图上不同字母表示2个品种在处理间存在显著性差异(< 0.05)。
Different letters above the bar mean significant difference between Longyou 7 (L7) and Tianyou 2 (T2) (< 0.05).
3.2 根系与叶片的膜脂在低温响应上存在差异, 根对北方白菜型冬油菜抗寒性的影响较大
本试验中, 随处理温度的降低, 2个冬油菜品种叶片不饱和脂肪酸含量变化均呈先降低后增加的趋势。在–5°C时不饱和脂肪酸大幅增加, 所增加的不饱和脂肪酸主要是棕榈油酸和亚麻酸; 根中不饱和脂肪酸含量变化趋势因品种不同, 陇油7号逐步增加, 天油2号则逐步降低; 根中不饱和脂肪酸含量变化以亚油酸为主。品种的不饱和脂肪酸含量变化趋势不同。2°C处理时天油2号叶片和根系不饱和脂肪酸含量均明显下降, 而陇油7号则叶片下降、根系显著上升, 说明两个抗寒性不同的品种的根系与叶片的膜脂在低温响应上存在差异, 抗寒性强的品种根系在降温过程中逐步获得了较稳定的膜脂不饱和度水平, 在低温下始终保持膜的流动性, 维持正常的生理功能, 以此抵御低温伤害。
3.3 ATP酶活性与冬油菜抗寒性的关系
低温胁迫会影响细胞膜系中的功能蛋白如ATP酶活性等[37], 然后对细胞生理、生化代谢过程造成破坏[38], ATP酶是膜结合功能性蛋白质, 在细胞中主要执行K、Na、Ca、Mg和H离子的传递功能[39-40]。简令成等[41]对小麦幼苗寒害的研究指出, 低温胁迫下, 质膜完整性受到破坏后透性发生变化, K+等外渗, 打破了离子平衡, 引起代谢失调。寒冷胁迫能刺激细胞质内Ca2+浓度增高[42], 但胞内Ca2+浓度过高或长时间维持高浓度会干扰细胞能量代谢系统和许多生理功能, 导致细胞死亡[43]。Ca2+-ATPase在维持细胞质内Ca2+稳态方面起重要作用[44]; 已有研究表明[45-46], 冷害首先是通过影响植物细胞膜的Mg2+- ATP 酶活性, 使之降低或完全失活, 从而破坏植物的生活力。在本研究中, 陇油7号叶片Ca2+/ Mg2+- ATPase和K+/Na+-ATPase活性均随着胁迫温度的降低而降低, 而其根系Ca2+/Mg2+-ATPase和K+/Na+- ATPase活性均随着胁迫温度的降低而升高; 天油2号叶片Ca2+/Mg2+-ATPase和K+/Na+-ATPase活性均随着胁迫温度的降低呈现波动状态, 其根系Ca2+/ Mg2+-ATPase和K+/Na+-ATPase活性均随着胁迫温度的降低而下降; 当温度达到–5°C时, 陇油7号根系ATPase显著高于天油2号。表明在低温胁迫下, 陇油7号能更好地维持根系的ATPase活性, 根系能有效地把低温刺激所产生的过量胞质离子及时地运到胞外, 使胞内离子维持稳态平衡, 保持细胞正常生理代谢, 抵御低温逆境胁迫的能力增强; 天油2号在低温下, 根系ATPase活性显著下降, 致使被升高的胞质无机离子不能及时外运, 造成细胞伤害, 引起代谢失调, 抗低温胁迫的能力降低, 导致抗寒性减弱。
3.4 低温胁迫下不饱和脂肪酸含量和ATPase活性的提高是品种抗寒的重要原因
生物膜的流动性和膜结合酶活力都受膜脂成分的影响[19], 抗寒锻炼能通过改变植物细胞膜的脂肪酸组成来提高植物的抗寒性[47-48]。本研究中, 低温胁迫对2个冬油菜品种叶片和根系脂肪酸饱和度及ATP酶活性均产生显著影响, 强抗寒性品种陇油7号在零度以下低温胁迫时, 不饱和脂肪酸含量/饱和脂肪酸含量比值、IUFA、Ca2+/Mg2+-ATPase和K+/ Na+-ATPase活性均大于抗寒性较弱的天油2号, 说明抗寒性强的冬油菜品种在长期低温适应性生长过程中, 自身产生了适应低温的机制, 当温度降低时, 其膜脂脂肪酸不饱和度增加, 膜脂的流动性增强, 从而有利于膜脂维持稳定的状态; 同时, 脂肪酸组分含量的改变, 致使膜结合酶在低温下的活性增强, 有利于维持细胞质正常的无机离子水平, 避免细胞内无机离子中毒, 保护细胞免受伤害。王育启等[49]、李素丽等[50]在水稻、甘蔗的研究中也得到类似研究结果。因此低温下不饱和脂肪酸含量和ATPase活性的提高是北方强抗寒冬油菜品种主要抗寒特征。
4 结论
低温胁迫下2个冬油菜品种的膜脂脂肪酸组分相同, 但脂肪酸含量因品种抗寒性而异。低温条件下, 抗寒性强的品种脂肪酸不饱和度、IUFA值和ATPase活性均高于抗寒性较弱的品种, 而且, 在降温过程中, 抗寒性强的品种根系不饱和程度持续增加, 说明根中不饱和脂肪酸代谢合成在北方白菜型冬油菜抗寒(越冬)中起着非常重要的作用。低温胁迫下不饱和脂肪酸含量和ATPase活性的提高是抗寒品种抗寒的重要原因。
[1] 王洪春. 植物抗性生理. 植物生理学通讯, 1981, (6): 72–81 Wang H C. Physiological responses of plants to anti-environment., 1981, (6): 72–81 (in Chinese with English abstract)
[2] Kasamo K, Kagita F, Yamanishi H, Sakaki T. Low temperature-induced changes in the thermotropic properties and fatty acid composition of the plasma membrane and tonoplast of cultured rice (L.) cells., 1992, 33: 609–616
[3] Lyons J M, Wheaton T A, Pratt H K. Relationship between the physical nature of mitochondrial membranes and chilling sensitivity in plants., 1964, 39: 262–268
[4] 王洪春, 汤章城, 苏维埃, 王文英, 李锦树. 水稻干胚膜脂脂肪酸组分差异性分析. 植物生理学报, 1980, 6: 227–236 Wang H C, Tang Z C, Su W A, Wang W Y, Li J S. Analysis of difference in fatty acid composition of membrane lipids of dry rice embryo., 1980, 6: 227–236 (in Chinese with English abstract)
[5] 杨玲, 钱建东, 方芳, 苏维埃. 佛手柑叶磷脂酰甘油相变和脂肪酸组成的差异. 植物生理学通讯, 1995, 31: 196–197 Yang L, Qian J D, Fang F, Su W A. Difference of thermal phase transition and fatty acid composition of phosphatidylglycerols in fingered citron leaves., 1995, 31: 196–197 (in Chinese with English abstract)
[6] Lyons J M. Chilling injury in plants., 1973, 24: 445–466
[7] 赵金梅, 周禾, 孙启忠, 郭强, 李芳. 植物脂肪酸不饱和性对植物抗寒性影响的研究. 草业科学, 2009, 26(9): 129–134 Zhao J M, Zhou H, Sun Q Z, Guo Q, Li F. Study on the influence of plant fatty acid desaturation on cold tolerance., 2009, 26(9): 129–134 (in Chinese with English abstract)
[8] Larus B, Sangwan V, Omann F, Dhindsa R S. Early steps in cold sensing by plant cells: the role of actin cytoskeleton and membrane fluidity., 2000, 23: 785–794
[9] 夏明, 刘亚学, 阿拉木斯, 李志勇. 低温下苜蓿叶片膜脂脂肪酸组分的研究. 中国草地, 2002, 24(6): 28–31 Xia M, Liu Y X, AL M S. Li Z Y. Fatty acid composition of cell membrane of alfalfa leaves at chilling temperature., 2002, 24(6): 28–31 (in Chinese with English abstract)
[10] 孙汉洲, 赵芳, 李志辉. 用细胞膜膜脂脂肪酸成分分析法筛选抗寒巨桉种源. 中南林学院学报, 2000, 20(3): 59–62 Sun H Z, Zhao F, Li Z H. Selection of the cold-resistant species provenances of eucalyptus grand is by using the method of composition analysis of fatty acids of cell membrane., 2000, 20(3): 59–62 (in Chinese with English abstract)
[11] 杨玉珍, 罗青, 彭方仁. 不同种源香椿叶片脂肪酸组分含量与抗寒性关系. 林业科技开发, 2011, 25(5): 21–25 Yang Y Z, Luo Q, Peng F R. A relationship between cold resistance and content of fatty acid composition in leaves offrom different provenances., 2011, 25(5): 21–25 (in Chinese with English abstract)
[12] 宋芳琳, 张苗苗, 苏金乐, 柴军舰, 张广信. 自然降温过程中猴樟和香樟幼苗的抗寒性比较. 西部林业科学, 2012, 41(6): 48–52 Song F L, Zhang M M, Su J L, Chai J J, Zhang G X. The comparison on cold resistance betweenandseedlings in natural decreasing process of air temperature., 2012, 41(6): 48–52 (in Chinese with English abstract)
[13] Murelli C, Rizza F, Albini F M, Dulio A, Terzi V, Cattivelli L. Metabolic changes associated with cold acclimation in contrasting cultivars of barley., 1995, 94: 87–93
[14] 王萍, 张成军, 陈国祥, 王静, 施大伟, 吕川根. 低温对水稻剑叶膜脂过氧化和脂肪酸组分的影响. 作物学报, 2006, 32: 568–572 Wang P, Zhang C J, Chen G X, Wang J, Shi D W, Lyu C G. Effects of low temperature on lipid peroxidation and fatty acid composition of flag leaf in rice (L.)., 2006, 32: 568–572 (in Chinese with English abstract)
[15] 王金胜, 郭定成. 温度与膜脂脂肪酸对不同抗冷性玉米幼苗ATP酶的影响. 山西农业大学学报, 1994, 14: 290–292 Wang J S, Guo D C. Temperature and fatty acid of membrane lipids effected on activity of ATPase in mitochondria of maize seedling belonging to four chilling-resistant level., 1994, 14: 290–292 (in Chinese with English abstract)
[16] Caldogno R F, Maria I, Michelis D, Maria C, Pugliarello. Active transport of Ca2+in membrane vesicles from pea: evidence for a H+/ Ca2+antiport., 1982, 639: 287–295
[17] 宋立新, 王瑞秀. 离子转运ATPase的结构和功能. 生理科学进展, 1989, 20: 334–338 Song L X, Wang D X. Structure and function of ion transporter ATPase., 1989, 20: 334–338 (in Chinese with English abstract)
[18] 郑翠兵. 盐胁迫下甜菜碱对甜菜光合作用及抗氧化能力的影响. 黑龙江大学硕士学位论文, 黑龙江哈尔滨, 2011Zheng C B. Effects of Betaine on Photosynthesis and Antioxidant Ability of Sugarbeet Under Salt Stress. MS Thesis of University of Heilongjiang, Harbin, China, 2011 (in Chinese with English abstract)
[19] 薛刚, 刘凤霞. 干旱对棉花根和下胚轴质膜脂肪酸组分及其相关酶活性的影响. 植物生理学通讯, 1997, 33(2): 97–100 Xue G, Liu F X. Effects of drought stress on fatty acid composition and involved enzyme activity of plasma membrane in cotton root and hypocoty., 1997, 33(2): 97–100 (in Chinese with English abstract)
[20] 李俊明, 张敬贤, 崔四平, 魏建昆, 张海明, 耿庆汉. 膜脂组成和膜结合酶活性与玉米幼苗抗冷性的关系. 华北农学报, 1992, 7(3): 50–53 Li J M, Zhang J X, Cui S P, Wei J K, Zhang H M, Geng Q H. The relationship among membrane lipids membrane linked enzymes and maizecold-resistance., 1992, 7(3): 50–53 (in Chinese with English abstract)
[21] 刘炜. 低温下冷敏感植物春小麦和抗寒植物冬小麦细胞内Ca2+的稳态平衡能力的比较研究. 山东师范大学硕士学位论文, 山东济南, 2000 Liu W. Difference of Ca2+-homeostasis between Chilling- Sensitive Spring Wheat and Chilling-Resistant Winter Wheat under Low Temperature. MS Thesis of Shandong Normal University, Jinan, China, 2000 (in Chinese with English abstract).
[22] 王红, 孙德兰, 卢存福, 简令成. 抗寒锻炼对冬小麦幼苗质膜Ca2+-ATPase的稳定作用. 植物学报, 1998, 40: 1098–1101 Wang H, Sun D L, Lu C F, Jian L C. Stability effects of cold-acclimation on the plasmolemma Ca2+-ATPase of winter wheat seedlings., 1998, 40: 1098–1101 (in Chinese with English abstract)
[23] 王精明, 李美茹. 低温对水稻幼苗根细胞质膜、液泡膜Mg2+-ATP酶活性的影响. 湖北农学院学报, 2000, 20: 295–297 Wang J M, Li M R. Effect of low temperature on the activity of Mg2+-ATP in root plasma membrane and tonoplast membrane of rice seedlings., 2000, 20: 295–297 (in Chinese with English abstract).
[24] 孙万仓, 刘海卿, 刘自刚, 武军艳, 李学才, 方彦, 曾秀存, 许耀照, 张亚宏, 董云. 北方寒旱区白菜型冬油菜安全越冬的临界指标分析. 作物学报, 2016, 42: 609–618 Sun W C, Liu H Q, Liu Z G, Wu J Y, Li X C, Fang Y, Zeng X C, Xu Y Z, Zhang Y H, Dong Y. Critical index analysis of safe over-wintering rate of winter rapeseed () in cold and arid region in north China., 2016, 42: 609–618 (in Chinese with English abstract)
[25] 徐燕, 薛立, 屈明. 植物抗寒性的生理生态学机制研究进展. 林业科学, 2007, 43(4): 88–94 Xu Y, Xue L, Qu M. Physiological and ecological mechanisms of plant adaptation to low temperature., 2007, 43(4): 88–94 (in Chinese with English abstract)
[26] 刘自刚, 孙万仓, 杨宁宁, 王月, 何丽, 赵彩霞, 史鹏飞, 杨刚, 李学才, 武军艳, 方彦, 曾秀存. 冬前低温胁迫下白菜型冬油菜抗寒性的形态及生理特征. 中国农业科学, 2013, 46: 4679–4687 Liu Z G, Sun W C, Yang N N, Wang Y, He L, Zhao C X, Shi P F, Yang G, Li X C, Wu J Y, Fang Y, Zeng X C. Morphology and physiological characteristics of cultivars with different levels of cold-resistance in winter rapeseed (L.) during cold acclimation., 2013, 46: 4679–4687 (in Chinese with English abstract)
[27] 杨宁宁, 孙万仓, 刘自刚, 史鹏辉, 方彦, 武军艳, 曾秀存, 孔德晶, 鲁美宏, 王月. 北方冬油菜抗寒性的形态与生理机制. 中国农业科学, 2013, 47: 452–461 Yang N N, Sun W C, Liu Z G, Shi P H, Fang Y, Wu J Y, Zeng X C, Kong D J, Lu M H, Wang Y. Morphological characters and physiological mechanisms of cold resistance of winter rapeseed in northern China., 2013, 47: 452–461 (in Chinese with English abstract)
[28] 朱惠霞, 孙万仓, 邓斌, 燕妮, 武军艳, 范惠玲, 叶剑, 曾军, 刘雅丽, 张亚宏. 白菜型冬油菜品种的抗寒性及其生理生化特性. 西北农业学报, 2007, 16(4): 34–38 Zhu H X, Sun W C , Deng B, Yan N, Wu J Y, Fan H L, Ye J, Zeng J, Liu Y L, Zhang Y H. Study on cold hardiness and its physiological and biochemical characteristics of winter turnip rape ()., 2007, 16(4): 34–38 (in Chinese with English abstract)
[29] 史春会. 低温胁迫下不同类型秋菊品种主成分分析及抗寒性评价. 河南农业大学硕士学位论文, 河南郑州, 2011 Shi C H. Principal Component Analysis and Cold Tolerance Evaluation of Membrane Lipid Component in Different Varieties of Autumn Chrysanthemum Cultivers under Low Temperature. MS Thesis of Henan Agricultural University, Zhengzhou, China, 2011 (in Chinese with English abstract)
[30] Yoshida S. Reverse change in plasma membrane properties upon deacclimation of mulberry trees., 1986, 27: 83–89
[31] 余泽宁. 龙眼叶片膜脂脂肪酸组分与龙眼耐寒性的关系. 亚热带植物科学, 2003, 32(2): 15–17 Yu Z N. Cold tolerance of longyan in relation to membrane fatty acid composition in leaves., 2003, 32(2): 15–17 (in Chinese with English abstract)
[32] 王萍, 张成军, 陈国祥, 王静, 施大伟, 吕川根, 左敏, 刘蓉蓉. 低温对水稻幼苗类囊体膜脂肪酸组分和膜脂过氧化的影响. 中国水稻科学, 2006, 20: 401–405 Wang P, Zhang C J, Chen G X, Wang J, Shi D W, Lyu C G, Zuo M, Liu R R. Effects of low temperature on fatty acid composition of thylakoid membranes and lipid. peroxidation in leaves of rice seedlings,, 2006, 20: 404–405 (in Chinese with English abstract)
[33] 谢冬微, 王晓楠, 付连双, 孙健, 关涛, 李卓夫. 低温胁迫对冬小麦分蘖节膜脂脂肪酸的影响. 麦类作物学报, 2013, 33: 746–751 Xie D W, Wang X N, Fu L S, Sun J, Guan T, Li Z F. Effects of low temperature stress on membrane fatty acid in tillering node of winter wheat., 2013, 33: 746–751 (in Chinese with English abstract)
[34] Douglas O, Brain D P. Responses of plant to low, nonfreezing temperature: protein metabolism, and acclimation., 1982, 33: 347–372
[35] 梁及芝, 李志辉, 黄志文, 朱日光, 黄晓明. 耐寒性桉树早期选择: II. 巨桉种源和家系幼林生长. 中南林学院学报, 2000, 20(3): 75–79 Liang J Z, Li Z H, Huang Z W, Zhu R G, Huang X M. Early selection in cold-resistant eucalyptus: II. Analysis on growth of the young forests of eucalyp tus grand is provenances and families., 2000, 20(3): 75–79 (in Chinese with English abstract)
[36] 李志辉, 汤珧华, 孙汉洲, 左海松. 耐寒性桉树早期选择Ⅲ: 巨桉种源和家系膜脂肪酸组成、含量与抗寒性关系. 中南林学院学报, 2000, 20(3): 80–85 Li Z H, Tang Z H, Sun H Z, Zuo H S. Early selection in cold-resistant eucalyptus iii. the relation of composition and content of fatty acid and ability of cold-resistant of eucalyptus grand is provenance and families., 2000, 20(3): 80–85 (in Chinese with English abstract)
[37] 刘建. 两种桉树对低温胁迫的响应机制研究. 南京林业大学博士学位论文, 江苏南京, 2008 Liu J. Studies on Response Mechanisms of Two Eculypts to Low Temperature Stress. PhD Dissertation of Nanjing Forestry University, Nanjing, China, 2008 (in Chinese with English abstract)
[38] 简令成. 植物抗寒机理研究的新进展. 植物学通报, 1992, 9(3): 17–22 Jian L C. Advances of the studies on the mechanism of plant cold hardiness., 1992, 9(3): 17–22 (in Chinese with English abstract)
[39] 刘鸿先, 曾韶西, 李平. 植物抗寒性与酶系统多态性的关系. 植物生理学通讯, 1981, (6): 6–11 Liu H X, Zeng S X, Li P. The relationship between plant cold tolerance and enzyme system., 1981, (6): 6–11 (in Chinese with English abstract)
[40] Hammerten R W, Krzeminski K A, Mays R W, Ryan T A, Wollner D A, Nelson W J. Mechanism for regulation cell surface distribution of Na+, K+-ATPase in polarized epithelial cells., 1991, 254: 847–849
[41] 简令成, 孙龙华, 卫翔云, 王红, 张红. 从细胞膜系统的稳定性与植物抗寒性关系研究到抗寒剂的研制. 植物学通报, 1994, 11: 1–22 Jian L C, Sun L H, Wei X Y, Wang H, Zhang H. From the study on the stability of cellular membrane system in relation with plant cold hardiness to the creation of plant cold resister., 1994, 11: 1–22 (in Chinese with English abstract)
[42] 许大全. 光合作用学. 北京: 科学出版社, 2013. pp 215–218 Xu D Q. Photosynthesis Science. Beijing: Science Press, 2013, pp 215–218
[43] 曾韶西, 李美茹. 冷和盐预处理提高水稻幼苗抗寒性期间细胞Ca2+-ATPase酶活性的变化. 植物学报, 1999, 41: 156–160 Zeng S X, Li M R. Changes of Ca2+-ATPase activities in cell of rice seedlings during the enhancement of chilling resistance induced by cold and salt pretreatment., 1999, 41: 156–160 (in Chinese with English abstract)
[44] Bonza M C, Michelis M I D. The plant Ca2+-ATPase repertoire: biochemical features and physiological functions.(Stuttg), 2011, 13: 421–430
[45] 简令成, 董合铸, 孙龙华. 番茄子叶细胞内三磷酸腺苷酶活性的超微结构定位及其在冷害中的变化. 植物学报, 1981, 23: 257–261 Jian L C, Dong H Z, Sun L H. Ultrastructural localization of adenosine tri phosphatase activity in cotyledon cells of tomato and its changes during chilling stress., 1981, 23(3): 257–261 (in Chinese with English abstract)
[46] 戴金平, 沈征言, 简令成. 低温锻炼对黄瓜幼苗几种酶活性的影响. 植物学报, 1991, 33: 627–632 Dai J Y, Shen Z Y, Jian L C. The effects of cold acclimation on several enzyme activities in cucumber seedling., 1991, 33: 627–632 (in Chinese with English abstract)
[47] Thomashow M F. Plant cold acclimation: freezing tolerance genes and regulatory mechanisms., 1999, 50: 571–599
[48] Uemura M, Steponkus P L. Cold acclimation in plants: relationship between the lipid composition and the cryostability of the plasma membrane., 1999, 112: 245–254
[49] 王育启, 王洪春. 生物膜组分对膜功能和膜相变的调控. 植物生理学报, 1981, 7: 185–192 Wang Q Y, Wang H C. Regulation and control of function and phase transition in membrane by composition of biomembrane lipid., 1981, 7: 185–192 (in Chinese with English abstract)
[50] 李素丽, 杨丽涛, 李志刚, 李杨瑞, 韩春旺, 梁兆宙. 不同冷敏感型甘蔗茎尖Ca2+和Ca2+-ATP酶活性对低温的响应. 中国农业大学学报, 2011, 16(2): 14–21 Li S L, Yang L T, Li Z G, Li Y R, Han C W, Liang Z Z. Response of Ca2+and Ca2+-ATPase activity in the stem tip of sugarcane to low temperature stress., 2011, 16(2): 14–21 (in Chinese with English abstract)
Response of Membrane Fatty Acid Composition and ATPase Activity inL. to Temperature in North China
FANG Yan1,2, SUN Wan-Cang1,2,*, WU Jun-Yan2, LIU Zi-Gang2, DONG Yun2,3, MI Chao2, MA Li2, CHEN Qi2, and HE Hui-Li2
1Gansu Provincial Key Laboratory of Arid Land Crop Sciences, Lanzhou 730070, Gansu, China;2Rape Engineering and Technology Research Center of Gansu Province, Lanzhou 730070,Gansu, China;3Crops Institute of Gansu Academy of Agricultural Sciences, Lanzhou 730070, Gansu, China
L. cultivars Longyou 7 (cold tolerant) and Tianyou 2 (cold sensitive) were used to investigate the variations of membrane fatty acid composition and ATPase activity at the temperatures of 25°C, 10°C, 2°C, and –5°C. There was the same membrane fatty acid composition in leaf and root of two cultivars under temperature stresses, with linolenic acid as the main component of unsaturated fatty acid in leaf, but linoleic acid in root. With decrease of treatment temperatures, the content of unsaturated fatty acid in leaf initially decreased at 10°C and 2°C, then increased at –5°C. The content of unsaturated fatty acid rose up gradually in root of Longyou 7, but reduced in Tianyou 2. At low temperature (2°C, –5°C), the ratio of U/S and IUFA in Longyou 7 were significantly higher than those in Tianyou 2. The activity of ATPase was gradually increased in Longyou 7 than in Tianyou 2. It suggested there are differences in membrane lipids in response to temperature for two winter rapeseed cultivars, the increase of unsaturated fatty acid content and ATPase activity is the main cause sustaining winter rapeseed cultivars to over winter.
winter rapeseed; fatty acid; ATPase activity; cold tolerance
2017-01-20;
2017-09-10;
2017-09-28.
10.3724/SP.J.1006.2018.00095
通信作者(Corresponding author): 孙万仓, E-mail: 18293121851@163.com
E-mail: ffyv@163.com
本研究由国家现代农业产业技术体系建设专项(CARS-12), 国家自然科学基金项目(31460356, 31560397), 国家重点基础研究发展计划项目(973计划) (2015CB150206), 国家重点研发计划项目(2016YFD0101300)和甘肃农业大学盛彤笙创新基金(GSAU-STS-1428)资助。
This study was supported by the National Modern Agro-industry Technology System (CARS-13), the National Natural Science Foundation of China (31460356, 31560397), the National Key Basic Research Program of China (973 Program) (2015CB150206), the National Key Research and Development Program (2016YFD0101300), and the Sheng Tongsheng Innovation Fund of Gansu Agricultural University (GSAU-STS-1428).
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20170928.1458.008.html
猜你喜欢
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
农业工程学报(2022年5期)2022-06-22
热带作物学报(2022年5期)2022-06-01
农技服务(2021年6期)2021-09-22
中国生殖健康(2020年2期)2020-12-09
山西农业科学(2020年10期)2020-10-16
中国生殖健康(2018年2期)2018-11-06
浙江农业科学(2017年8期)2017-08-30
中学科技(2016年5期)2016-05-12
