
滇龙胆香叶醇合酶基因的克隆与表达分析
2018-01-16 18:38刘倩倩李彩霞谷从璟张晓东
广东农业科学 2017年9期
刘倩倩,李彩霞,赵 静,谷从璟,张晓东
(玉溪师范学院资源环境学院,云南 玉溪 653100)
香叶醇是一种非环式单萜醇,广泛应用于香精香料工业[1-5]。此外,香叶醇也是大部分药用植物、香料植物产生香味的主要化学成分[6],除了用于日用化学品、卷烟和食品加工等领域[7],还可用作驱虫剂、抗菌剂、抗肿瘤剂和汽油替代品[5,7-8],市场供不应求。目前,香叶醇主要通过植物提取和化学合成手段生产。由于天然植物香叶醇含量低,化学合成的原料为化工产品不可再生且易导致环境污染,使用现代生物技术手段生产香叶醇具有非常好的前景。因此,需要对香叶醇生物合成途径关键基因进行克隆和功能研究。
随着基因克隆和DNA测序技术的迅速发展,香叶醇生物合成途径相关基因相继被发现和克隆。香叶醇合成酶基因是香叶醇生物合成途径的主要基因之一。研究表明,香叶醇合成酶(GES,EC 3.1.7.3)能够催化香叶基焦磷酸(GPP)生成香叶醇,是单萜生物合成途径的限速酶[9-11]。在植物中,起初人们认为从GPP到香叶醇的反应是由磷酸酶或单萜合成酶催化的。在罗勒中,ObGES基因的分离,为单萜合成酶参与精油重要成分香叶醇的合成提供了首个证据[11]。接下来,在精油中能够累积香叶醇的细毛樟[12]、缬草[13]、甜蛇草[13]、藿香[6]和紫苏[14]等植物中,分离和鉴定了其中的GES基因。Dong等[13]分别从缬草和甜草中分离出VoGES和LdGES基因,在大肠杆菌中表达后,酶活分析结果表明其对底物GPP的Km分别为32、51μmol/L;GFP融合试验结果表明VoGES主要定位于质体,而LdGES分别定位于质体与细胞质。将长春花CrGES基因在大肠杆菌中过表达,其纯化蛋白能够催化GPP生成香叶醇,Km为58.5μmol/L;CrGES基因在叶和芽中特异性表达,并且其表达被茉莉酸甲酯(JA)诱导[9,11]。在结构方面,Sato等[1]研究发现,紫苏香叶醇合酶结构域IV-1和IV-4对香叶醇的形成非常重要,结构域IV-4中的5个氨基酸通过定位正电荷在碳阳离子中间体的分布,来决定羟基引入的位置。
在基因工程方面,普遍采用大肠杆菌、模式植物和酵母来生产香叶醇。使用大肠杆菌生产香叶醇中,ObGES是一个代谢调节阀[15]。Chen等[10]将喜树去除叶绿体转运肽的tCaGES和FPPS在大肠杆菌中表达,通过优化培养基组成、发酵时间和添加重金属离子等条件,可使香叶醇产量达到48.5(±0.9)mg/L。Liu等[3]在大肠杆菌BL21中过表达罗勒(Ocimum basilicum)香叶醇合酶基因ObGES、巨冷杉(Abies grandis)AgGPPS和一个异源甲羟戊酸途径,使用补料分批发酵方式,使香叶醇产量达到78.8 mg/L;使用水-有机双相发酵系统,通过酰基转移酶处理,最终获得2.0 g/L香叶醇的产量,这是迄今已报道的最高产量。Fischer等[16]分别将ObGES基因转化葡萄、烟草、拟南芥和细菌,结果香叶醇产量分别为烟草>葡萄>拟南芥>细菌,均远低于ObGES基因在酵母中的产量499(±188)μg/g鲜重,表明香叶醇合酶的功能特性不仅取决于其氨基酸序列,也取决于表达细胞的背景。Ritala等[17]通过将缬草质体定位的VoGES基因转入烟草毛根中来生产香叶醇,结果表明游离香叶醇含量高达31.3 μg/g干重,香叶醇衍生物主要包含己糖和戊糖香叶醇结合物、羟化香叶醇等6种形式,脱糖反应后香叶醇含量高达204.3 μg/g干重。Liu等[18]将密码子优化的罗勒ObGES基因整合到酵母基因组中,产量达到0.93 mg/L。Jiang等[5]通过基因筛选、截短CrGES蛋白N端的转运肽、在酵母中共表达截短的CrGES和FPPS方法,采用分批补料发酵技术,使香叶醇产量达到1.68 g/L。Zhao等[5]则通过在酵母LEU2原养型菌株中删除OYE2(编码NADPH氧化还原酶)、动态调控ERG20的表达、使用乙醇进行补料分批发酵,使香叶醇产量达到1.69 g/L,是目前已报道酵母的最高产量。
滇龙胆(Gentiana rigescens)是云南道地药材,主要用于治疗肝炎和胆囊炎[19-21]。目前,滇龙胆是龙胆泻肝片、苦胆草片和小儿清热片等200多种中药的主要原料[22]。本研究以滇龙胆为材料,利用RT-PCR技术成功地克隆了香叶醇合酶基因,对其编码蛋白基因功能进行预测,进行原核表达,并采用荧光定量PCR方法分析该基因在根和叶中的表达,为下一步研究其在香叶醇及滇龙胆药效成分龙胆苦苷合成中的功能奠定基础。
1 材料与方法
1.1 试验材料
滇龙胆植物材料、主要药品试剂和仪器同文献[23],采样日期为2016年5月15日。
1.2 试验方法
1.2.1 滇龙胆RNA提取和基因克隆 滇龙胆RNA的提取和基因克隆步骤参照张玲等[23]的方法,GrGES基因特异引物见表1。以cDNA为模板进行PCR扩增,PCR产物经电泳分离、胶回收、末端加A、连接到pMD19T载体,获得重组载体pMD19-GrGES。
1.2.2GrGES基因及其编码蛋白的生物信息学分析 采用张玲等[23]的方法对GrGES基因及其编码蛋白进行序列分析。
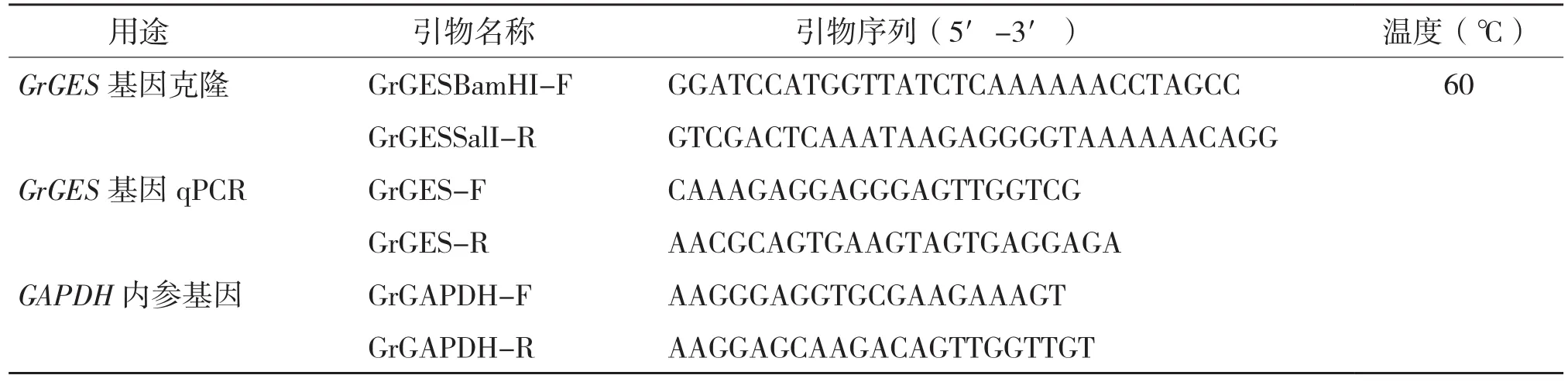
表1 GrGES基因克隆和荧光定量分析的引物序列
1.2.3 原核表达载体的构建 使用限制酶BamHI和SalI,分别对已测序质粒pMD19-GrGES和载体pET32a进行双酶切,然后分别回收目的基因和载体片段,按4∶1(摩尔比)的比例连接过夜,然后转化感受态细胞E.ColiDH5α,涂布于氨苄青霉素(100 mg/L)+IPTG(24 mg/L)+ X-gal(20 mg/L)的LB固体平板,37℃过夜培养后挑取阳性菌落;摇菌、提质粒,通过酶切检测大小正确后,进行DNA测序,最终获得原核表达载体pET32a-GrGES。
1.2.4GrGES基因原核表达 采用42℃热激法将pET32a-GrGES重组质粒转化E.ColiRosetta(DE3)表达菌感受态细胞,挑单菌落于含抗生素的LB液体培养基中,37℃过夜培养。次日以1∶100(菌种∶培养基)比例接种到新鲜的LB液体培养基(不含抗生素)中,37℃摇床培养3 h,然后在37℃、1 mmol/L IPTG的诱导下进行表达,以同条件下空载体pET32a转化菌做对照;分别诱导0、2、4和6 h后,超净台中取2 mL。4℃离心1 min集菌,倒掉上清,加超纯水100 μL、5 × 蛋白上样缓冲液25 μL,振荡悬菌,100℃煮5 min裂解菌体。4℃、12 000 r/min离心10 min。取20 μL样品进行上样,然后进行SDS-PAGE(5%浓缩胶和12%分离胶)电泳检测。
1.2.5 滇龙胆GrGES基因的实时荧光定量表达分析 滇龙胆GrGES基因的组织特异性表达分析参考张玲等[23]的方法,对三年生滇龙胆的根和叶进行采样、RNA提取、逆转录,然后进行qPCR反应。选择滇龙胆GrGAPDH基因作为内参基因。qPCR引物见表1。定量数据采用比较Ct值的2-△△Ct法进行分析处理。
2 结果与分析
2.1 滇龙胆GrGES基因的克隆
使用GrGES基因特异性引物和滇龙胆幼叶的第一链cDNA模板进行PCR反应,扩增出大约2 000 bp的DNA片段(图1)。通过TA克隆、酶切检测和DNA测序方法,获得正确的重组质粒pMD19-GrGES。
2.2 GrGES基因的生物信息学分析
2.2.1GrGES基因序列分析 将GrGES基因测序结果提交GenBank数据库,获得序列登录号KJ917168。利用Genetyx软件对GrGES基因进行序列分析,结果表明该基因长1 767 bp,共编码588个氨基酸,这与长春花CrGES基因大小类似[9]。
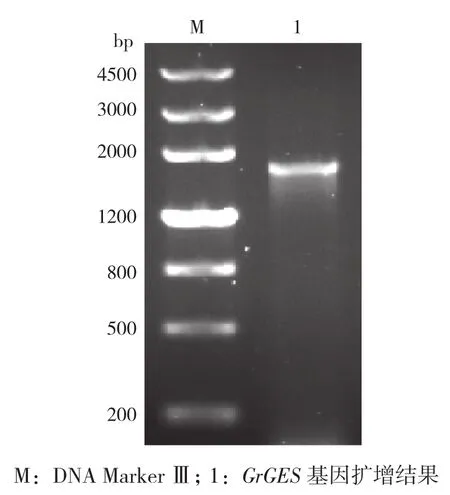
图1 滇龙胆GrGES基因的PCR扩增结果

图2 滇龙胆GrGES蛋白与其他药用植物GES蛋白的多序列比对分析
利用Gen Bank数据库蛋白比对程序BLASTp对GrGES蛋白进行比对分析,结果表明滇龙胆GrGES与长春花CrGES蛋白序列相似性最高(70.65%),与杨树PtGES(45.12%)蛋白相似性相对较低。选择与GrGES蛋白相似性较高的序列,采用DNAMAN 8进行多序列比对分析,结果(图2)表明,GrGES蛋白与已知GES蛋白序列具有较高保守性。利用MEGA7.0.26将GrGES氨基酸序列与BLASTp中相似性较高的长春花(Catharanthus roseus,AFD64744.1)、缬草(Valeriana officinalis,AGB05612.1,AGB05613.1,AHE41084.1)、油橄榄(Olea europaea,AFI47926.1)、芝麻(Sesamum indicum,XP_011080637.1)、桂花(Osmanthus fragrans,AMB57285.1)、甜舌草(Phyla dulcis,ADK62524.1)胡黄连(Picrorhiza kurrooa,AHX97739.1)、喜树(Camptotheca acuminata,ALL56347.1)、罗勒(Ocimum basilicum,Q6USK1.1)、葡萄(Vitis vinifera,NP_001267920.1)、杨树(Populus trichocarpa,AEI52903.1)等11种植物的GES蛋白序列进行系统发育分析,结果(图3)显示,滇龙胆GrGES蛋白与长春花CrGES聚为同一进化枝,表明二者亲缘关系很近,GrGES蛋白参与萜类生物碱龙胆苦苷生物合成途径。
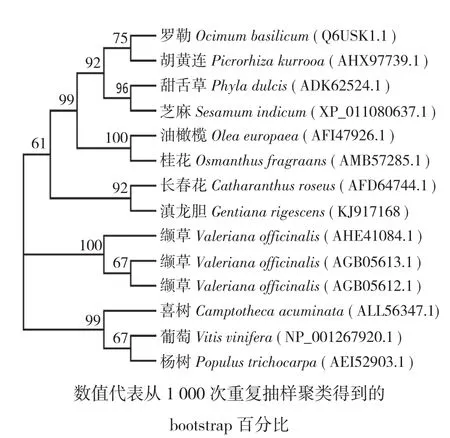
图3 滇龙胆GrGES蛋白与其他植物GES蛋白的系统发育分析
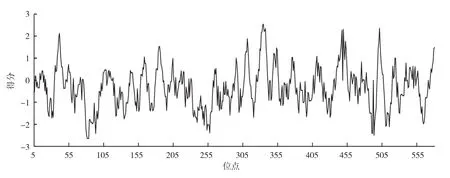
图4 滇龙胆GrGES蛋白的疏水性分析
2.2.2 GrGES蛋白的理化特性分析 使用ProtParam工具对GrGES蛋白进行分析,结果表明GrGES蛋白单体相对分子量为67.84 ku,pI为5.56,与长春花CrGES蛋白类似[9];带负电氨基酸残基(Asp + Glu)为83,带正电氨基酸残基(Arg + Lys)为68,化学方程式为:C3031H4778N806O894S32。不稳定指数为55.21,属不稳定蛋白;脂肪指数92.06,总平均疏水性(GRAVY)-0.29,为亲水蛋白(图4)。GrGES蛋白包含20种基本氨基酸,其中亮氨酸含量最高(12.80%);其次是谷氨酸和丝氨酸(9.50%和8.00%);半胱氨酸含量最低(1.40%)。
2.2.3 GrGES蛋白二级和三级结构分析 利用Predictprotein对GrGES蛋白进行二级结构进行预测,结果表明该蛋白二级结构中α-螺旋占62.41%,环占39.57%,延伸带占0.68%;暴露于外部的蛋白占31.63%,被包埋的蛋白占59.18%,介于二者的中间部分占9.18%。利用Swiss-Model Workspace软件自动模式对GrGES蛋白的三级结构进行预测,结果如图5所示。该3D模型以灰杨(Populus×canescens)异戊二烯合酶[3n0f.1]为模板,在第106~573氨基酸处建模,序列相似性为44.28%。
2.2.4 GrGES蛋白结构域分析 使用InterPro在线软件对GrGES蛋白保守结构域进行预测,结果显示GrGES蛋白具有其他植物GES蛋白萜类环化酶的活性位点(底物结合口袋cd00684,305R、314W、335I、337I、338L、339L、342D、346D、417F、484R、485L、487D、488D、491T、 495E、564Y、569D、571Y)、金属结合位点,包含2个保守结构域(图6),分别为萜类合酶N端结构域(IPR001906,81~273)和萜类合酶结构域(IPR005630,263~588)。
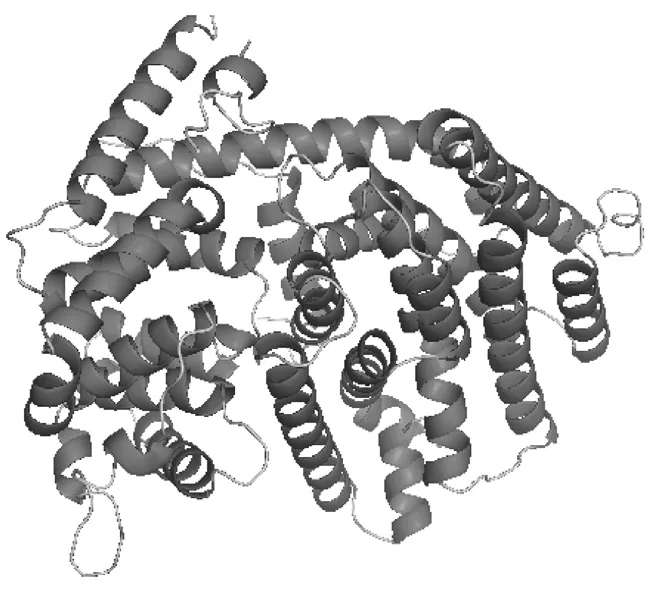
图5 滇龙胆GrGES蛋白的3D结构预测
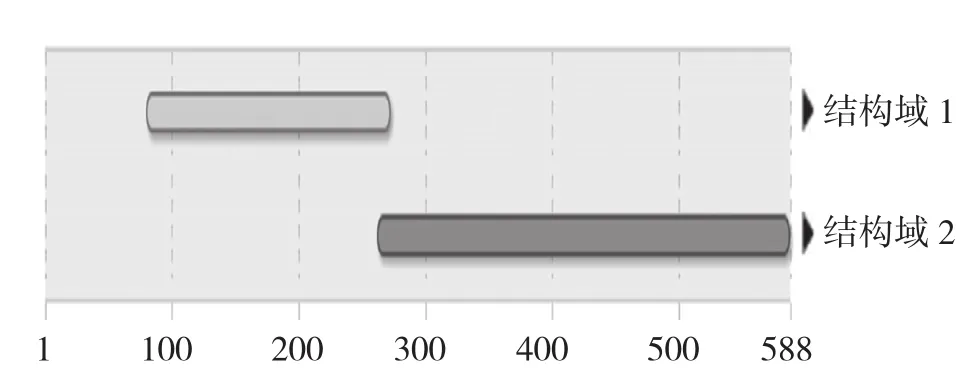
图6 滇龙胆GrGES蛋白保守结构域的预测
2.2.5 GrGES蛋白信号肽预测 使用SignalP 4.1对GrGES蛋白信号肽进行预测,未发现信号肽,表明该蛋白为非分泌型蛋白。采用ChloroP v1.1对GrGES蛋白叶绿体转运肽进行分析,结果显示GrGES蛋白含有1个叶绿体转运肽,长度为44个氨基酸。利用TMHMM对GrGES蛋白跨膜螺旋区进行预测,结果(图7)显示GrGES蛋白不含有跨膜区域。
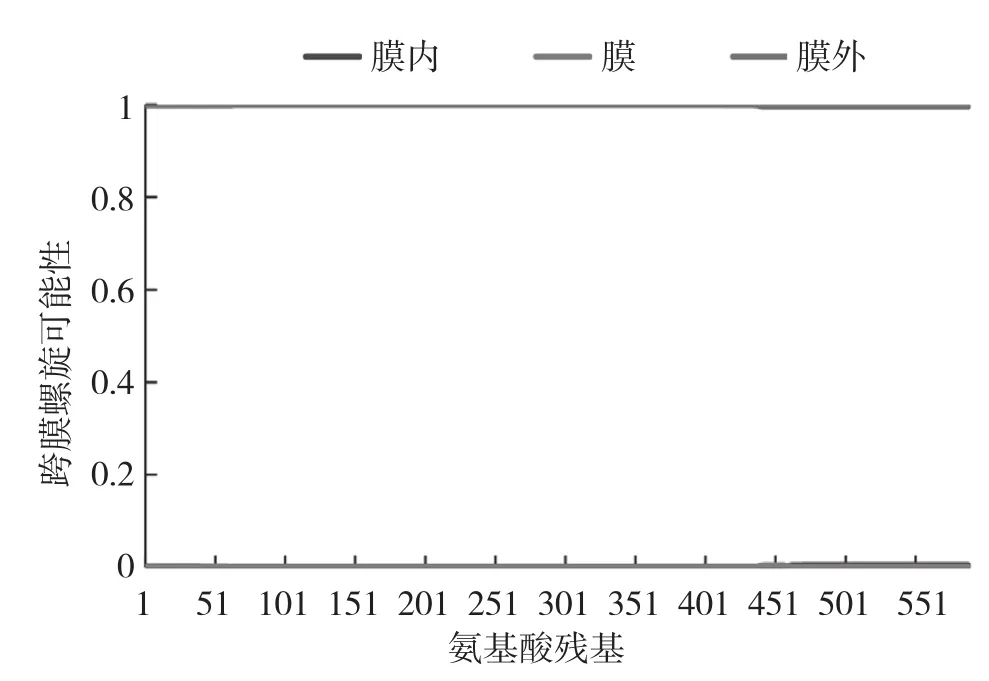
图7 滇龙胆GrGES蛋白可能跨膜螺旋的检测
2.2.6 GrGES蛋白亚细胞定位预测 使用WoLF PSORT对GrGES蛋白进行亚细胞定位预测,结果显示GrGES蛋白在叶绿体和细胞质中的定位系数分别为12.0和2.0,表明该蛋白定位于叶绿体。
2.2.7GrGES基因稀有密码子分析 使用在线软件对GrGES基因稀有密码子进行预测,结果显示该基因中稀有密码子为1.70%,仅含有1个二联稀有密码子,因此可采用E.coli表达菌Rosetta(DE3)进行原核表达。
2.3 GrGES基因原核表达载体的构建
使用快切酶BamHI和SalI双酶切检测重组质粒pET32a-GrGES,可切出载体和目的基因(图8),表明GrGES基因已成功插入载体pET32a。对酶切检测正确的质粒,进行DNA测序,表明未出现移码现象。
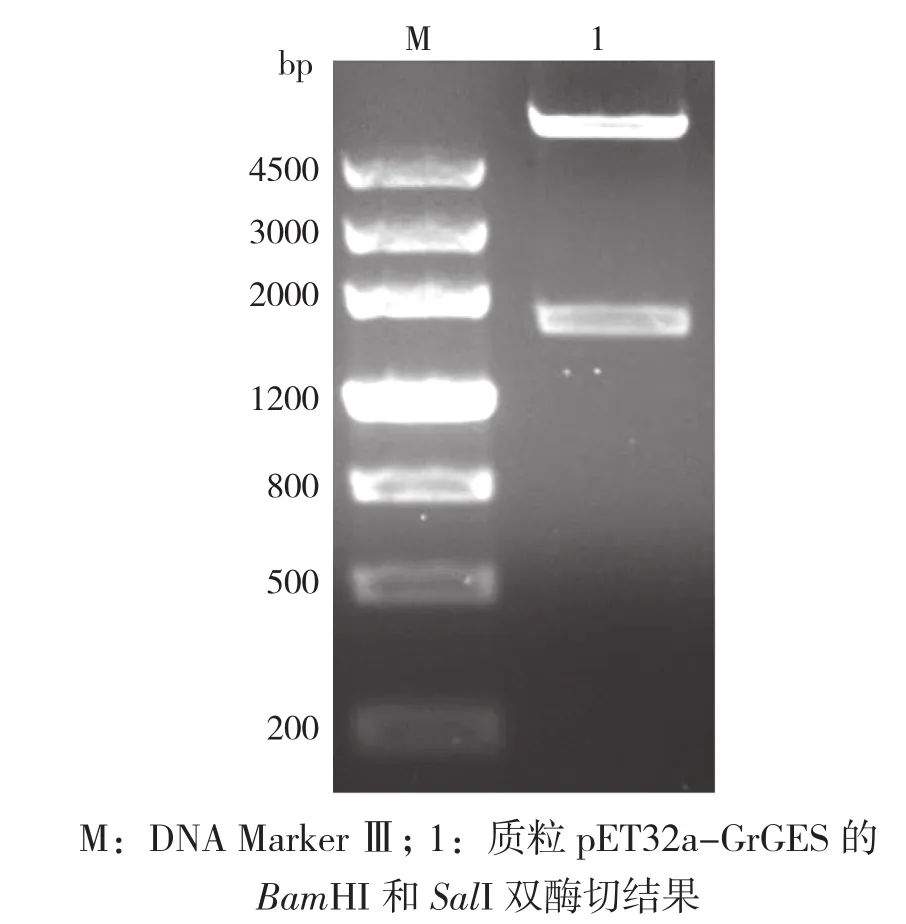
图8 质粒pET32a-GrGES双酶切检测
2.4 GrGES基因的原核表达
将重组质粒pET32a-GrGES转化E.ColiRosetta(DE3)后,在37℃、1 mmol/L IPTG下,分别诱导表达0、2、4、6 h,然后提取细菌总蛋白,进行SDS-PAGE分析。结果(图9)表明,与对照(第1~3泳道)相比,经IPTG诱导后,转化菌pET32a-GrGES在相对分子质量78.22 ku(含Trx、S·Tag、His·Tag等标签蛋白约17.71 ku)左右出现一蛋白条带,且GrGES融合蛋白表达量随诱导时间增加而逐渐增加,这表明pET32a-GrGES重组质粒在E.ColiRosetta(DE3)中已成功诱导表达出GrGES蛋白。
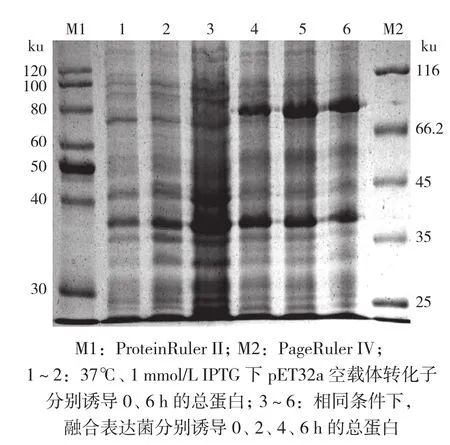
图9 37℃下不同诱导时间对GrGES蛋白表达的影响
2.5 GrGES基因的组织特异性表达分析
取三年生盆栽滇龙胆的根和叶,分别提取总RNA,反转录成第一链cDNA后,通过实时荧光定量RT-PCR分析GrGES基因在根和叶中的表达情况。结果表明,GrGES基因在叶中表达量很高,约是根的3 000倍。
3 结论与讨论
香叶醇合酶能够催化GPP合成香叶醇,其编码基因已从长春花[9]、缬草[17]、喜树[10]等许多植物中被克隆和研究,但是植物次生代谢具有很强的种属特异性,模式植物中所获得的结果往往不具有普遍性,因此需要对不同植物基因进行研究[24]。本研究从滇龙胆中成功克隆到GrGES基因,其编码蛋白GrGES包含一个高度保守的富含天冬氨酸的基序DDIFD和另一个高度保守的NSE/DTE基序(保守序列为LWDDLGTAKEE)(图2)。大多数植物GES蛋白均包含这两种基序,主要功能为参与焦磷酸底物的结合与依赖金属离子的离子化[10],这表明本研究所克隆的基因为GES基因。存在于许多植物中的N末端RRX8W基序主要参与GES蛋白的质体定位,在转运到质体后会被切除[10],但是,在GrGES中并未发现该基序。由于GrGES蛋白氨基酸疏水性的特征,使用WoLF PSORT软件对GrGES蛋白进行亚细胞定位预测时,该蛋白定位于叶绿体,使用ChloroP v1.1软件对GrGES蛋白叶绿体转运肽进行预测,结果也显示GrGES蛋白含有一个44个氨基酸组成的叶绿体转运肽。这与缬草VoGES蛋白和长春花CrGES蛋白的亚细胞定位一致[9,17],与甜舌草LdGES蛋白定位于叶绿体和细胞质不同[13]。
基因的组织表达特异性决定了其代谢产物的组织特异性。在长春花中,CrGES基因主要在叶和芽中特异性表达,并且其表达能够被茉莉酸甲酯诱导[11]。本研究中,GrGES基因在叶中表达量远远高于根中,说明香叶醇的生物合成主要在叶中进行。本研究为香叶醇的生物合成及滇龙胆分子生物学研究提供基因资源,并为GrGES蛋白结构和功能的研究奠定基础。
[1]Sato-Masumoto NIto M.A domain swapping approach to elucidate differential regiospecific hydroxylation by geraniol and linalool synthases from perilla[J].Phytochemistry,2014,102:46-54.
[2]Zhao J,Bao X,Li C,et al.Improving monoterpene geraniol production through geranyl diphosphate synthesis regulation inSaccharomyces cerevisiae[J].Applied Microbiology and Biotechnology,2016,100(10):4561-4571.
[3]Liu W,Xu X,Zhang R,et al.EngineeringEscherichia colifor high-yield geraniol production with biotransformation of geranyl acetate to geraniol under fed-batch culture[J].Biotechnol Biofuels,2016,9:58.
[4]Jiang G Z,Yao M D,Wang Y,et al.Manipulation of GES and ERG20 for geraniol overproduction inSaccharomyces cerevisiae[J].Metabolic Engineering,2017,41:57-66.
[5]Zhao J,Li C,Zhang Y.Dynamic control of ERG20 expression combined with minimized endogenous downstream metabolism contributes to the improvement of geraniol production inSaccharomyces cerevisiae[J].Microbial cell factories,2017,16(1):17.
[6]欧阳蒲月,曾少华,莫小路.广藿香香叶醇合酶基因克隆及表达分析[J].西北植物学报,2016,36(5):896-901.
[7]田宁,咸漠,胡仰栋,等.产香叶醇重组大肠杆菌发酵培养基的优化[J].林产化学与工业,2015,35(4):131-137.
[8]张松,孙丽,李悦.香叶醇在医学领域的应用研究进展[J].西北药学杂志,2017,32(1):124-126.
[9]Simkin A J,Miettinen K,Claudel P,et al.Characterization of the plastidial geraniol synthase fromMadagascar periwinklewhich initiates the monoterpenoid branch of the alkaloid pathway in internal phloem associated parenchyma[J].Phytochemistry,2013,85:36-43.
[10]Chen F,Li W,Jiang L,et al.Functional characterization of a geraniol synthaseencoding gene fromCamptotheca acuminataand its application in production of geraniol inEscherichia coli[J].Journal of Industrial Microbiology & Biotechnology,2016,43(9):1281-1292.
[11]Kumar K,Kumar S R,Dwivedi V,et al.Precursor feeding studies and molecular characterization of geraniol synthase establish the limiting role of geraniol in monoterpene indole alkaloid biosynthesis inCatharanthus roseusleaves[J].Plant Science,2015,239:56-66.
[12]Yang T,Li J,Wang H X,et al.A geraniolsynthase gene fromCinnamomum tenuipilum[J].Phytochemistry,2005,66(3):285-293.
[13]Dong L,Miettinen K,Goedbloed M,et al.Characterization of two geraniol synthases fromValeriana officinalisandLippia dulcis:similar activity but difference in subcellular localization[J].Metabolic Engineering,2013,20:198-211.
[14]Ito M,Honda G.Geraniol synthases from perilla and their taxonomical significance[J].Phytochemistry,2007,68(4):446-453.
[15]Zhou J,Wang C,Yang L,et al.Geranyl diphosphate synthase:an important regulation point in balancing a recombinant monoterpene pathway inEscherichia coli[J].Enzyme and Microbial Technology,2015,68:50-55.
[16]Fischer M J,Meyer S,Claudel P,et al.Specificity ofOcimum basilicumgeraniol synthase modified by its expression in different heterologous systems[J].Journal of Biotechnology,2013,163(1):24-29.
[17]Ritala A,Dong L,Imseng N,et al.Evaluation of tobacco(Nicotiana tabacumL.cv.Petit Havana SR1)hairy roots for the production of geraniol,the first committed step in terpenoid indole alkaloid pathway[J].Journal of Biotechnology,2014,176:20-28.
[18]Liu J,Zhang W,Du G,et al.Overproduction of geraniol by enhanced precursor supply inSaccharomyces cerevisiae[J].Journal of Biotechnology,2013,168(4):446-451.
[19]Suyama Y,Tanaka N,Kurimoto S,et al.Studies on medicinal plants of Yunnan province:Constituents ofGentiana rigescens[J].Planta Medica,2014,80(10):785.
[20]Suyama Y,Kurimoto S,Kawazoe K,et al.Rigenolide A,a new secoiridoid glucoside with a cyclobutane skeleton,and three new acylated secoiridoid glucosides fromGentiana rigescensFranch[J].Fitoterapia,2013,91:166-172.
[21]董丽萍,倪梁红,赵志礼,等.龙胆属环烯醚萜类化学成分研究进展[J].中草药,2017,48(10):2116-2128.
[22]金航,张霁,张金渝,等.滇龙胆[M].昆明:云南科技出版社,2013.
[23]张玲,李彩霞,张海晨,等.滇龙胆异戊烯基焦磷酸异构酶基因的克隆与表达分析[J].广东农业科学,2015,42(17):139-146.
[24]陈晓亚,薛红卫.植物生理与分子生物学(第4版)[M].北京:高等教育出版社,2012.
猜你喜欢
基层中医药(2022年7期)2022-11-17
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
天然产物研究与开发(2018年10期)2018-11-06
中成药(2017年12期)2018-01-19
中成药(2016年4期)2016-05-17
中成药(2016年4期)2016-05-17
中国老年学杂志(2015年9期)2015-01-31
中国应用生理学杂志(2013年2期)2013-03-25
中国神经精神疾病杂志(2013年1期)2013-03-11
