
起源
2018-01-15 13:08
生物进化 2017年4期
地球生命可能有39.5亿年的历史

地球有46亿年的历史,意味着最原始的地壳形成于距今46亿年左右。一般认为,存在于始太古代(约36亿年至40亿年前)时期的原核生物——细菌与古菌,为生物的下一步进化打下了坚实的基础。但由于始太古代的岩石稀少且保存不佳,因此,有关地球早期生命存在的证据,迄今较为匮乏。已知最古老的变质碎屑岩来自加拿大拉布拉多北部的萨格利克地块,约有39.5亿年历史。近期,研究者详细调查了这些岩石的地质特征,并测量了石墨和碳酸盐的浓度和同位素组成。最终发现,这些石墨确实是生源的,并指出石墨的结晶温度和母岩的变质温度一致,这意味着石墨并非源自后期污染物。这表明,可能早在39.5亿年前,地球上就已存在有机生命,其或是地球上已知最早的一些生命。(Nature 2017,549:516-518)
基因给鹦鹉披上五彩衣
当谈到自然界中绚丽多姿的色彩时,鸟类和花无疑是的佼佼者。许多色彩鲜艳的鸟是从食物中获得色素的,但鹦鹉却是个例外。那它们是如何产生其特有的黄色、蓝色和绿色羽毛的呢?研究者发现,经过过去150年间的人工选择,虎皮鹦鹉出现了大量影响颜色的简单孟德尔遗传性状,因此,虎皮鹦鹉是研究鹦鹉色彩的绝佳模式动物。虎皮鹦鹉身上有一类未在其他脊椎动物中发现的表现为红色到黄色的色素。而且,人们还发现一些不能产生黄色素的长尾小鹦鹉的羽毛也有黄色到绿色到蓝色的颜色变换。然而,参与鹦鹉羽毛颜色的基因和生化途径一直不得而知。近期,研究者使用了全基因组关联图谱鉴定了一个包含蓝色突变的遗传区域,该区域富含几个基因,但不确定哪种基因起主要作用。为了缩小范围,研究人员测序了234只活虎皮鹦鹉(其中105只为蓝色)和澳大利亚15个博物馆鹦鹉标本的DNA。结果,蓝色鹦鹉携带单突变基因MuPKS,编码了一个鲜为人知的聚酮合成酶。而且,实验显示,MuPKS在绿黄鹦鹉羽毛上出现高量表达,但是蓝鹦鹉羽毛中MuPKS的一个单氨基酸被替换了。之后,研究者克隆了MuPKS基因,并将其插入酵母基因组以检查酵母是否会产生黄色颜料。结果正如所料。这些实验证明,基因表达模式赋予了鹦鹉变换的羽色。(Cell 2017,171:427-439.e21)

家鸡的矮小化
人类的驯化和选育使得家鸡成为表型多样性最为丰富的动物之一。在体型大小方面,大型肉鸡或斗鸡(大于5千克)和小型观赏鸡(约0.5千克)可达到10倍以上的悬殊差异。作为重要的经济性状,家鸡的体型一直受到育种工作者的长期关注。其中,家鸡的矮小化在蛋鸡和观赏鸡选育中至关重要。针对家鸡的矮小化现象,研究者尝试采用基于高通量测序技术的群体基因组学分析方法,对我国著名矮小化观赏鸡——元宝鸡展开研究。相关结果显示,BMP10基因在元宝鸡受强烈的人工选择作用,其上游启动子区域的突变导致其在元宝鸡心脏中表达上调,是导致其矮小化的重要原因。该基因可以解释元宝鸡22.41%的体型变异。相关工作也表明,运用群体基因组分析有望在较小的群体样本水平发掘家鸡复杂性状的主效基因。

有意思的是,矮小化的家鸡品系众多,其遗传机制是否相同?针对这一问题,研究者进一步选取了世界上体型最小的鸡——塞拉玛和我国云南著名的小型鸡——大围山微型鸡开展了群体基因组学研究。群体遗传结构分析揭示,元宝鸡、塞拉玛和大围山微型鸡有着独立的遗传组分。群体历史分析显示,元宝鸡和塞拉玛在选育过程中经历了强烈的瓶颈效应。选择信号和模拟计算分析锁定IGF1和POU1F1是决定塞拉玛矮小化的重要基因。相关基因均在生长发育相关通路中发挥重要的作用。这种选择信号组合的模式提示"杂种优势"效应是导致塞拉玛成为世界上体型最小鸡种的重要原因。对三个矮小化鸡种的比较分析显示,它们矮小化的遗传机制并不相同,很可能是多次独立选育的结果。相关结果也为今后的家鸡育种工作提供重要的靶点和参考。(Molecular Biology and Evolution 2017,34:3081-3088)
帕米尔人群的源流历史
帕米尔高原,古称"葱岭",由亚洲腹地的喜马拉雅山脉、兴都库什山山脉、天山山脉和昆仑山脉交汇而成。帕米尔高原平均海拔超过4000米,在1838年被到访的英国探险者约翰·伍德首次赋予了"世界屋脊"的称号。尽管具有高寒缺氧等严酷的环境条件,帕米尔高原在历史上是古丝绸之路的重要组成部分,东西人群交流往来频繁。然而,相关的人群源流历史却一直鲜为人知。为探讨帕米尔高原人群的源流历史,研究者对整个帕米尔高原代表性的色勒库尔塔吉克、瓦罕塔吉克、帕米尔塔吉克和高原柯尔克孜及其周边平原低地人群共计382份样本进行了线粒体DNA基因组测序。分析结果显示,人群体现出东西方遗传混合的特征。大多数混合事件发生在末次盛冰期之后。进一步的分析发现塔吉克人群与柯尔克孜人群之间存在明显的遗传分化。基于近似贝叶斯计算分析的结果揭示人类移居帕米尔高原涉及多次迁移和混合,是一个复杂的过程。通过比较平原人群的mtDNA编码区序列,研究人员未能在高原人群中检出受到选择信号。(European Journal of Human Genetics 2017,doi:10.1038/s41431-017-0028-8)

褐家鼠起源于东亚南部
褐家鼠是重要的伴人鼠类之一,除两级冰盖之外,几乎遍布全球,在人类活动区域非常常见。褐家鼠也是最主要的害鼠之一,携带多种病毒,传染类似鼠疫等的传染性疾病。关于褐家鼠的起源及扩散问题,学术界目前仍存在争议。大量的历史及研究基本确定褐家鼠是起源于亚洲,但是具体起源区域及迁徙路线尚无定论。近期,研究者利用高通量大规模测序平台对全球多地区110个褐家鼠样本进行测序,解析和阐明了褐家鼠的东亚南部起源及迁移问题。该研究从群体遗传学角度和计算生物学角度揭示了褐家鼠的东亚南部起源,并描绘了其从起源地往东亚北部以及欧洲/非洲/中东的两条主要扩散路线。对其迁出时间进行的推断显示:褐家鼠在~173,000年前从东亚南部迁往东亚北部;在~3,100年前,由东亚南部往中东迁徙,紧接着在~2,000年前往非洲迁徙,~1,800年前迁往欧洲。这推翻了人们之前认为褐家鼠起源于东亚北部的这一假说。同时,在褐家鼠的迁徙过程中,很多与免疫系统相关的基因受到了快速进化。(MolecularBiologyandEvolution2017,https://doi.org/10.1093/molbev/msx276)

人类肤色变异出现于90万年前
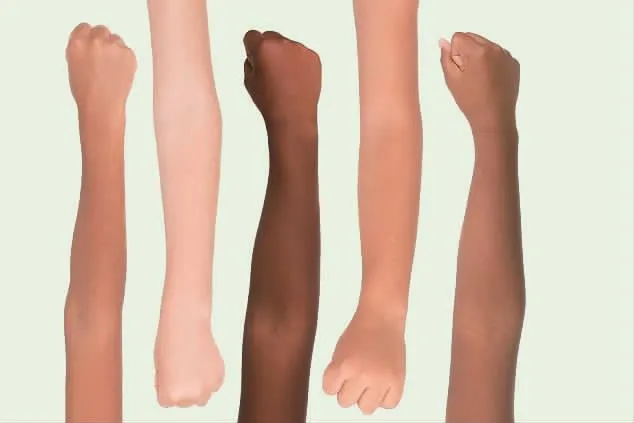
人类具有多种皮肤颜色。浅色皮肤和深色皮肤都会提供益处:比如,深色皮肤被认为有助于阻止紫外线照射带来的一些负面影响,而在较低紫外线照射的区域,浅色皮肤能够更好地促进维生素D合成。但是迄今为止,仅少量基因已被证实促进皮肤颜色发生正常的变化,而且这主要是通过研究欧洲人群发现的。近期,研究者在一项针对不同非洲群体的研究中鉴定出与皮肤色素沉着相关的新的基因变异。当人们谈及非洲人的肤色时,大多数人想到肤色较深的皮肤,但新研究显示在非洲,存在比价大的肤色变化:从一些非洲人具有较浅的肤色到全球水平上最深的肤色,以及介于这两者之间的肤色。研究者还鉴定出影响这些性状的基因变异,并且证实影响浅色皮肤和深色皮肤的突变在现代人类起源之前就已存在较长时间了。研究者综合各种数据,找到人类基因组中8个同皮肤色素沉着水平存在密切关联的位点。对于每个位点来说,均存在1个同较白皮肤相关的遗传变异以及1个同较黑皮肤存在关联的变异。在同较白皮肤相关的变异中,有7个至少在27万年前出现,其中有4个在90多万年前出现。最新的观点认为智人在约30万年前出现于非洲。但本研究的发现意味着相对较白的肤色变异早于人类的出现,并且自此以后被保留在非洲部分地区。同时,一些特别黑的肤色由控制较浅肤色的基因变异演化而来的时间相对较近,从而表明拥有较白皮肤的人群"更加高级"这一种族主义概念存在严重缺陷。(Science2017,10.1126/science.aan8433)
鸟类丢失牙齿的真正原因
喙是在包括鸟类在内的许多兽脚类恐龙中演化出的重要器官。早期的鸟类具有牙齿,而进步的鸟类及所有现生鸟类拥有角质喙。以往对现生鸟类喙的发育过程有比较深入的研究,但是对鸟类牙齿丢失的原因及其机制一直众说纷纭。传统观点认为牙齿的丢失有助于减轻鸟类体重,增强鸟类的飞行能力,也有观点认为是单纯的基因突变导致鸟类牙齿丢失。研究者曾在一种兽脚类恐龙——难逃泥潭龙中发现的牙齿在个体发育过程中逐渐丢失的现象,这一现象为阐明鸟类牙齿丢失的机制带来了新的曙光。在之后的研究中,研究者发现牙齿在个体发育过程中逐渐丢失的现象在窃蛋龙类和基干鸟类中同样存在,并且在恐龙向鸟类演化的过程中牙齿丢失的时间不断提前——这些现象说明牙齿的异时发育退化才是导致鸟类牙齿丢失直接原因。
牙齿的丢失必然伴随着取食器官的改变。研究者通过基于大数据的特征相关性分析发现,牙齿的丢失与角质喙的发育存在着必然的联系。以往的研究表明,骨形态发生蛋白BMP不仅控制着牙齿发育,也参与角质喙的发生过程。研究小组通过发育生物学实验证实,这一信号通路表达的上调在引起牙齿发育中断的同时还可以促进角质喙的生长——这一结果第一次阐明了牙齿异时发育退化可能的机制。新的研究表明,基因突变并不是导致鸟类牙齿丢失的真正原因。之前相关的分子遗传学研究主要是基于现生鸟类展开的,而鸟类的喙出现后自然选择对牙齿发生相关基因的作用逐渐减小,最终导致这些基因发生突变和丢失。因此,鸟类牙齿发育基因的突变应当是在鸟类演化后期发生的事件,并不是导致鸟类牙齿丢失的直接原因。这一现象不仅存在于恐龙-鸟类的支系中,而在几十个脊椎动物的支系中普遍存在。这说明牙齿的异时发育丢失是脊椎动物'丢牙'常用的'手段',它不仅颠覆了我们对鸟类喙演化历史已有的认识,对于理解脊椎动物牙齿演化的模式也具有深远的意义 "。(Pnas 2017,114:10930-10935)


牙齿的异时发育丢失在不同门类的脊椎动物中普遍存在
猜你喜欢
青少年科技博览(中学版)(2022年10期)2023-01-07
作文周刊·小学二年级版(2022年20期)2022-05-05
疯狂英语·新悦读(2020年4期)2020-06-18
好孩子画报(2020年3期)2020-05-14
小天使·四年级语数英综合(2019年9期)2019-11-09
疯狂英语·新阅版(2019年4期)2019-09-10
意林·全彩Color(2019年6期)2019-07-24
小学生作文(中高年级适用)(2017年4期)2017-07-10
知识就是力量(2017年1期)2017-01-06
家禽科学(2016年4期)2016-05-04
