
胶州湾海域5种粗毛猛水蚤的形态记述*
2018-01-10 09:08:00李新正
海洋科学集刊 2017年0期
马 林 李新正
胶州湾海域5种粗毛猛水蚤的形态记述*
马 林1, 2, 3李新正1, 2, 3①
(1. 中国科学院海洋研究所 海洋生物分类与系统演化实验室, 青岛 266071; 2. 中国科学院大学, 北京 100049; 3. 青岛海洋科学与技术国家实验室 海洋生物学与生物技术功能实验室, 青岛 266071)
粗毛猛水蚤科(Miraciidae Dana, 1846)是猛水蚤目(Harpacticoida Sars, 1903)种类数目居于第二的科, 分布广泛, 栖息环境多样。本文通过对2008年在胶州湾海域采集的沉积物进行离心提取, 经解剖和形态分类学方法鉴定, 鉴定粗毛猛水蚤科5属5种。其中, 田园威伦狭腹猛水蚤(Karanovic & Kim K., 2014)为我国海域的首次记录。文中还对胶州湾已发现的底栖粗毛猛水蚤制定了检索表。
粗毛猛水蚤科; 猛水蚤目; 胶州湾
粗毛猛水蚤科(Miraciidae Dana, 1846)是猛水蚤目(Harpacticoida Sars, 1903)数量居于第二的科, 主要在海洋中营底栖生活(Boxshall and Halsey, 2004), 世界上已记录55属376种。粗毛猛水蚤的栖息环境多样, 分布很广, 各大洋均有分布(Lang, 1965; Wells and Rao, 1987; Garlitska et al., 2012); 可生活在各种类型的生境中, 红树林、珊瑚礁、海草床、热液口和冷泉中均有分布; 垂直分布从潮间带到超过5000m的深海(Somerfield et al., 1998; Willen, 2003; Rose et al., 2005)。猛水蚤营养价值高, 可作为仔稚鱼和贝类等的食物, 在底栖生态系统小食物网中具有重要的作用(Lee et al., 2012); 另外, 猛水蚤繁殖周期短, 对污染非常敏感, 是监测环境污染的潜在指示种(Moreno et al., 2008; Araújo-Castro et al., 2009)。
粗毛猛水蚤科的分类学研究开始于19世纪40年代, Dana于1846年建立粗毛猛水蚤科, 但没有对种进行描述, 直到1849年报道了粗毛猛水蚤科2种, 此科的分类学研究才陆续开展。早期粗毛猛水蚤科的分类学研究主要集中在欧美国家沿岸, 我国邻近海域粗毛猛水蚤科的分类学研究开始较晚, 直到20世纪70年代日本才有粗毛猛水蚤的报道(Itô, 1972)。我国对海洋底栖粗毛猛水蚤科的分类学研究相对薄弱, 开始于21世纪, Mu和Gee(2000)首先描述了渤海的3种粗毛猛水蚤; 而对胶州湾海域底栖猛水蚤的物种形态描述研究更少, 仅有零星报道(Ma and Li, 2011)。
胶州湾位于山东半岛南岸的西部, 是一个半封闭的浅海内湾, 周围海域环境和地理位置优越, 底栖生物种类丰富。本文通过对胶州湾粗毛猛水蚤的鉴定, 为进一步明确胶州湾底栖猛水蚤的物种组成提供基础, 研究成果对于我国海洋生态学研究和环境监测具有重要意义。
1 材料与方法
刮取沉积物表层, 用10%甲醛固定, 带回实验室后, 染色, 以38µm孔径的网筛分选后用比重为1.15的胶态氧化硅溶液(Ludox-TM, 美国sigma公司)进行提取, 显微镜下挑选, 标本保存在75%的酒精中。解剖前将标本在乳酸中透明20min左右, 制片后显微镜下观察鉴定。
2 结果和讨论
(1) 羽球疑囊猛水蚤Bulbamphiascus plumosus Mu & Gee, 2000 (图1)
Mu & Gee, 2000: p. 105, figs. 1~8.
标本信息 2♀♀(MBM189103), 35°59.000′N, 120°25.500′E, 水深16.5m, 底质粗砂, 2008年5月14日李宝泉、周进、马林采自胶州湾。
形态特征 雌体体长约1mm。身体近圆柱形。额角三角形。尾节背面中部具一小的角质状的肛板。尾叉长宽几乎等长; 具7根尾毛, 第1尾毛细小。前体部无刺的分布; 后体部生殖节腹面和侧面具刺; 第4节腹面和侧面具小刺; 尾节侧面以及尾叉的侧面和腹面具刺。第1触角分8节; 第4节和末节各具一感觉毛。第2触角外肢分3节; 内肢边缘具2根刺和2根刚毛, 末端具2根羽状刺, 4根膝状弯曲的刚毛和1根羽状刚毛。大颚颚基具8齿和2根刚毛; 外肢分2节; 内肢1节。第1小颚内外肢均分1节。第2小颚底节具3个内叶, 刚毛分布为3∶3∶2; 内肢1节, 具5根刚毛。颚足内肢1节具5根刚毛和刺。第1胸足内肢第1节延伸到外肢第3节的中部。第1~4胸足各节刚毛和刺的分布如表1所示。
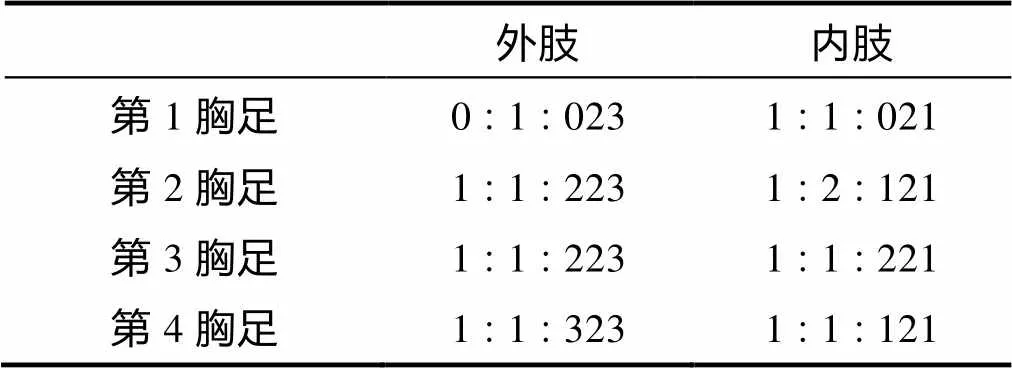
表1 羽球疑囊猛水蚤雌性第1~4胸足各节刚毛和刺的分布
第5胸足外肢长是宽的两倍, 具6根刚毛; 内肢突起末端延伸到外肢的1/2处。
讨论 Mu和Gee(2000)在对渤海湾软泥底质的沉积物进行分析时, 报道此种, 分布水深11~70m。胶州湾采集的标本个体比渤海湾采集的个体大, 身体外部形态、附肢各节刚毛和刺的分布情况与Mu和Gee(2000)描述的个体形态基本一致, 除1~3胸足底节刺的分布略有差别。
(2) 颂歌伊藤狭腹猛水蚤Itostenhelia polyhymniaKaranovic & Kim K., 2014 (图2~3)
Karanovic & Kim K., 2014: p. 54, figs. 34~42.
标本信息 3♀♀(MBM189106~189108), 1♂(MBM189109), 36º08.000′N, 120º11.167′E, 水深2.8m, 底质软泥, 2008年5月14日李宝泉、周进、马林采自胶州湾。
形态特征 雌体体长约560~646µm, 雄体体长约546µm。体前体部宽于后体部, 前体部体节表面无刺的分布。尾叉长是宽的约3.5倍, 具肛板。第1触角分不明显的8节, 第4节和末节各具1感觉毛。第2触角外肢分3节; 内肢末端具7根刚毛。大颚颚基内侧具2齿; 内肢向外肢弯曲。第1小颚内外肢均分1节。第2小颚内肢具4根刚毛。颚足底节外侧顶端具钝的凸起。第1胸足内肢第1节粗短。第1~4胸足各节刚毛和刺的分布如表2所示。
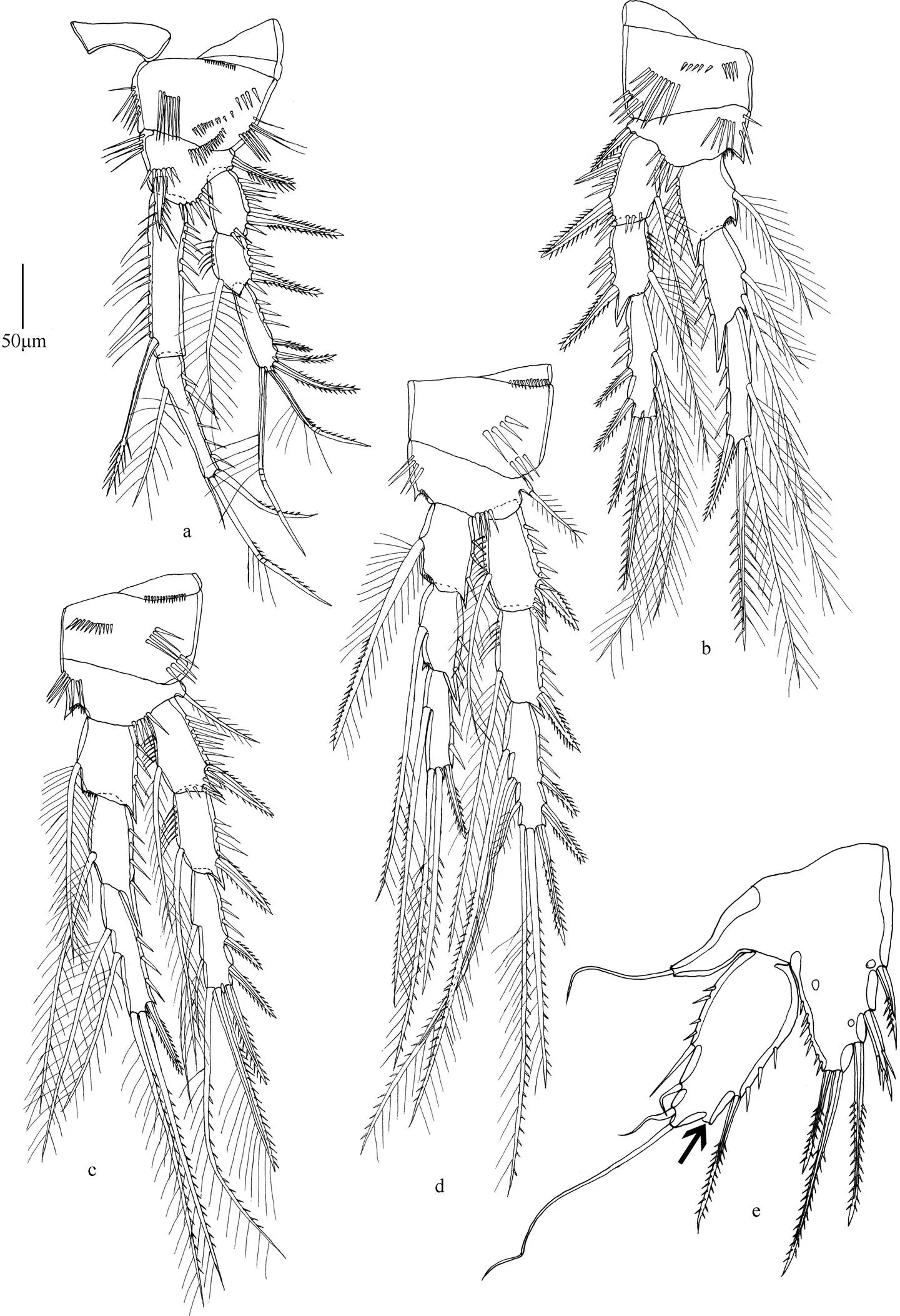
图1 羽球疑囊猛水蚤(雌性)
注: a. 第1胸足; b. 第2胸足; c. 第3胸足; d. 第4胸足; e. 第5胸足(箭头处表示缺失一根刚毛); 比例尺长度为50µm。
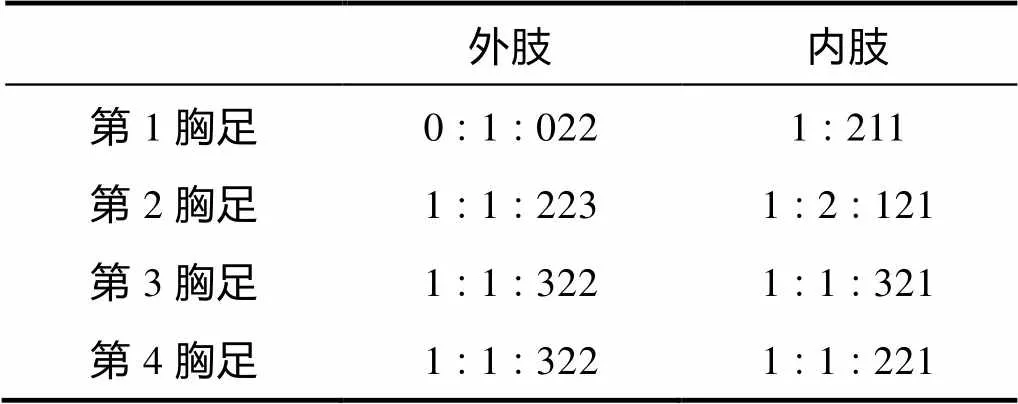
表2 颂歌伊藤狭腹猛水蚤雌性第1~4胸足各节刚毛和刺的分布
雌性第5胸足内肢突起具4根刚毛, 近外肢处具细刺; 外肢外侧具细刺, 具5根刚毛。雄性外肢具4根刚毛。
讨论 Karanovic和Kim(2014)对韩国海域采集的小个体标本和大个体标本进行过详细的描述。胶州湾采集的标本个体属于大个体标本, 与韩国报道的个体身体外部形态、附肢刚毛和刺的分布情况基本一致, 除雌性第5胸足内肢近外肢处的刺更密集。目前此种仅在韩国海域和胶州湾有记录。

图2 颂歌伊藤狭腹猛水蚤(雌性)整体图
(3) 双刺爪狭腹猛水蚤Onychostenhelia bispinosa Huys & Mu, 2008 (图4~5)
Huys & Mu, 2008: p. 53, figs. 1~7; Kim et al., 2011, p. 56, figs. 1~6; Lee et al., 2012, p. 118, figs. 78~83; Song et al., 2012, p. 1403.
标本信息 5♀♀, 2♂♂(MBM189116), 35º59′N, 120º25.5′E, 水深16.5m, 底质粗砂, 2008年5月14日李宝泉、周进、马林采自胶州湾。
形态特征 雌体体长530~650µm。雄体体长450~580µm。头胸甲比其他体节宽。额角三角形, 侧面凹陷, 顶端两半。尾叉圆柱形, 长约为基部宽的8倍, 具尾毛7根。第1触角7节, 第1节和第2节具凸起, 第4节和末节均具感觉毛。第2触角内肢侧缘具2根刺和2根刚毛, 末端具6个附属物; 外肢分3节。大颚外肢内侧具3根刚毛, 末端具3根刚毛; 内肢内侧具2根刚毛, 末端具6根刚毛。第1小颚内外肢基部愈合。第2小颚内肢具5根刚毛。颚足内肢分2节。第1胸足底节长大于宽, 基节短。第1~4胸足刚毛和刺的分布如表3所示。
第5胸足外肢末端具一刺状突起, 外侧具2根窄刺, 内侧具3根长的刚毛。内肢与基节愈合, 具5个附属物, 外侧第2根刚毛稍呈羽状, 中间的刚毛一侧具细刺, 一侧具小羽片, 内侧第2根刚毛与中间刚毛等长。雄性第5胸足不连续, 背面中部向外弯曲, 内侧刺靠近顶端处更粗壮。
讨论 本种与Huys和Mu(2008)的描述的个体身体外部形态和附肢上刚毛和刺的分布情况基本一致, 此种在中国渤海和黄海以及韩国海域均有分布。
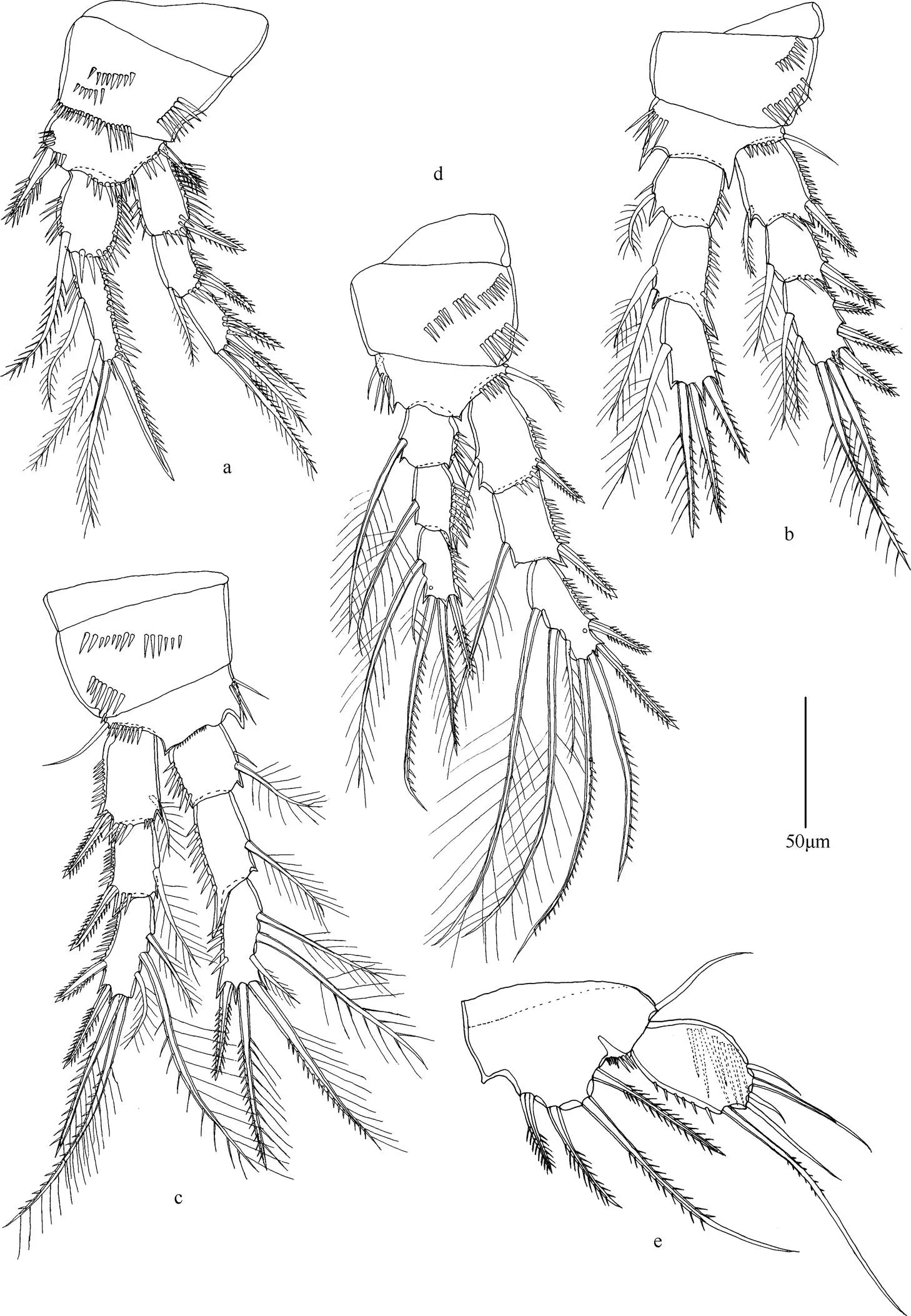
图3 颂歌伊藤狭腹猛水蚤(雌性)
注: a. 第1胸足; b. 第2胸足; c. 第3胸足; d. 第4胸足; e. 第5胸足; 比例尺长度为50µm。

表3 双刺爪狭腹猛水蚤雌性第1~4胸足各节刚毛和刺的分布
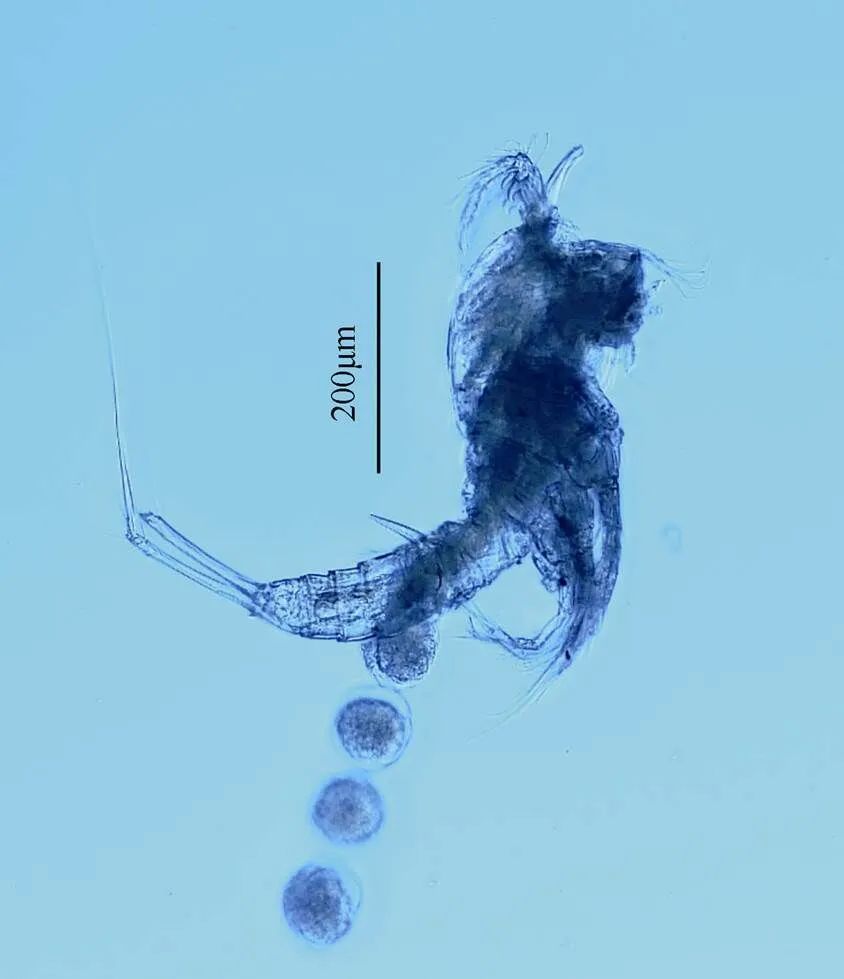
图4 双刺爪狭腹猛水蚤(雌性)整体图
(4) 显著华疑囊猛水蚤Sinamphiascus dominatus Mu & Gee, 2000 (图6)
Mu & Gee, 2000: p. 119, figs. 15~21; Nam & Lee, 2012, p. 46, figs. 1~8; Lee et al., 2012, p. 126, figs. 84~91; Song et al., 2012, p. 1403.
标本信息 34♀♀, 7♂♂(MBM189119), 35º59′N, 120º25.5′E, 水深16.5m, 底质粗砂; 23♀♀, 5♂♂(MBM189120), 36º09.000′N, 120º15.000′E, 水深5.0m, 底质软泥; 2008年5月14日李宝泉、周进、马林采自胶州湾。
形态特征 雌性个体最长达900µm, 雄性个体最长达825µm。近圆柱形。前体部各体节无刺的分布。后体部第4体节背面和腹面具刺, 尾节中部具半圆形的肛板。尾叉近方形。额角长, 呈三角形。第1触角分8节, 第4节和末节具感觉毛。第2触角外肢分3节; 内肢分2节, 末节侧面具2根刺和1根刚毛, 末端具1根刺和6根刚毛。大颚外肢分不明显的2节; 内肢1节, 具8根刚毛。第1小颚内外肢均分1节。第2小颚内肢1节, 具6根刚毛。颚足内肢1节, 具1爪和3根刚毛。第1~4胸足各节刚毛和刺的分布如表4所示。

表4 显著华疑囊猛水蚤雌性第1~4胸足各节刚毛和刺的分布
第5胸足具5根羽状刺, 几乎等长。外肢长约为宽的两倍, 内侧具1根刺, 末端具2根刚毛, 外侧具3根羽状刺。第5胸足外肢长是宽的约1.5倍, 具6根刚毛和刺; 内肢具2根羽状刚毛。
讨论 本文记录的显著华疑囊猛水蚤与Mu和Gee(2000)描述的个体身体外部形态、附肢刚毛和刺的分布情况基本相同, 除雌性后体部倒数第2体节后缘刺的数量更多, 分布更连续。这种变化可能是生境变化造成的。
(5) 田园威伦狭腹猛水蚤 Willenstenhelia thalia Karanovic & Kim K., 2014 (图7~8)
Karanovic & Kim K., 2014: p. 74, figs 48~54.
标本信息 3♀♀, 36º08.000′N, 120º11.167′E, MBM189240, 水深2.8m, 底质软泥, 2008年5月15日李宝泉、周进、马林采自胶州湾。
形态特征 雌体体长约380~400µm。体前体部宽于后体部, 前体部体节表面无刺的分布。尾叉长是宽的约4.6倍, 具肛板。第1触角分不明显的8节, 第4节具1感觉毛。第2触角外肢分3节; 内肢末端具7根刚毛。大颚颚基表面不具刚毛; 内肢向外肢弯曲。第1小颚内外肢均分1节。第2小颚内肢具4根刚毛, 外肢具2根刚毛。颚足底节外侧顶端具钝的凸起。第1~4胸足各节刚毛和刺的分布如表5所示。
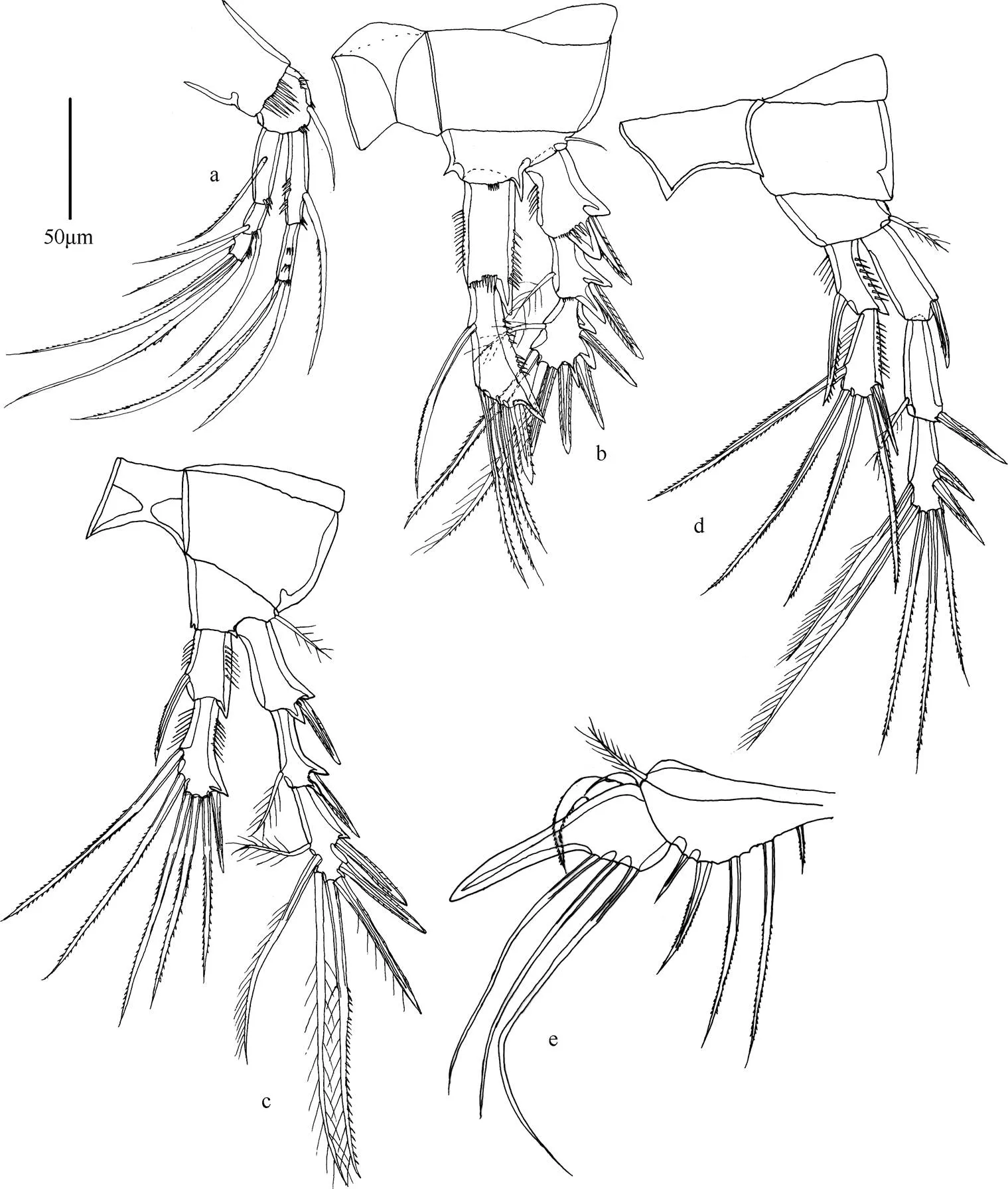
图5 双刺爪狭腹猛水蚤(雌性)
注: a. 第1胸足; b. 第2胸足; c. 第3胸足; d. 第4胸足; e. 第5胸足; 比例尺为50µm。
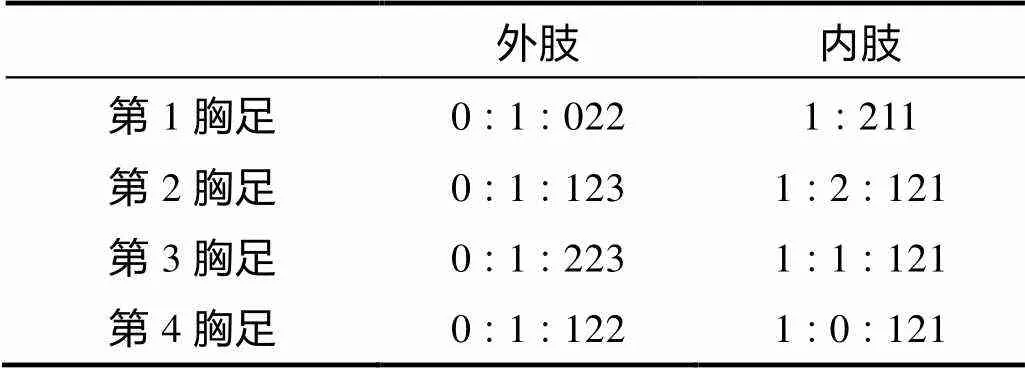
表5 田园威伦狭腹猛水蚤雌性第1~4胸足各节刚毛和刺的分布
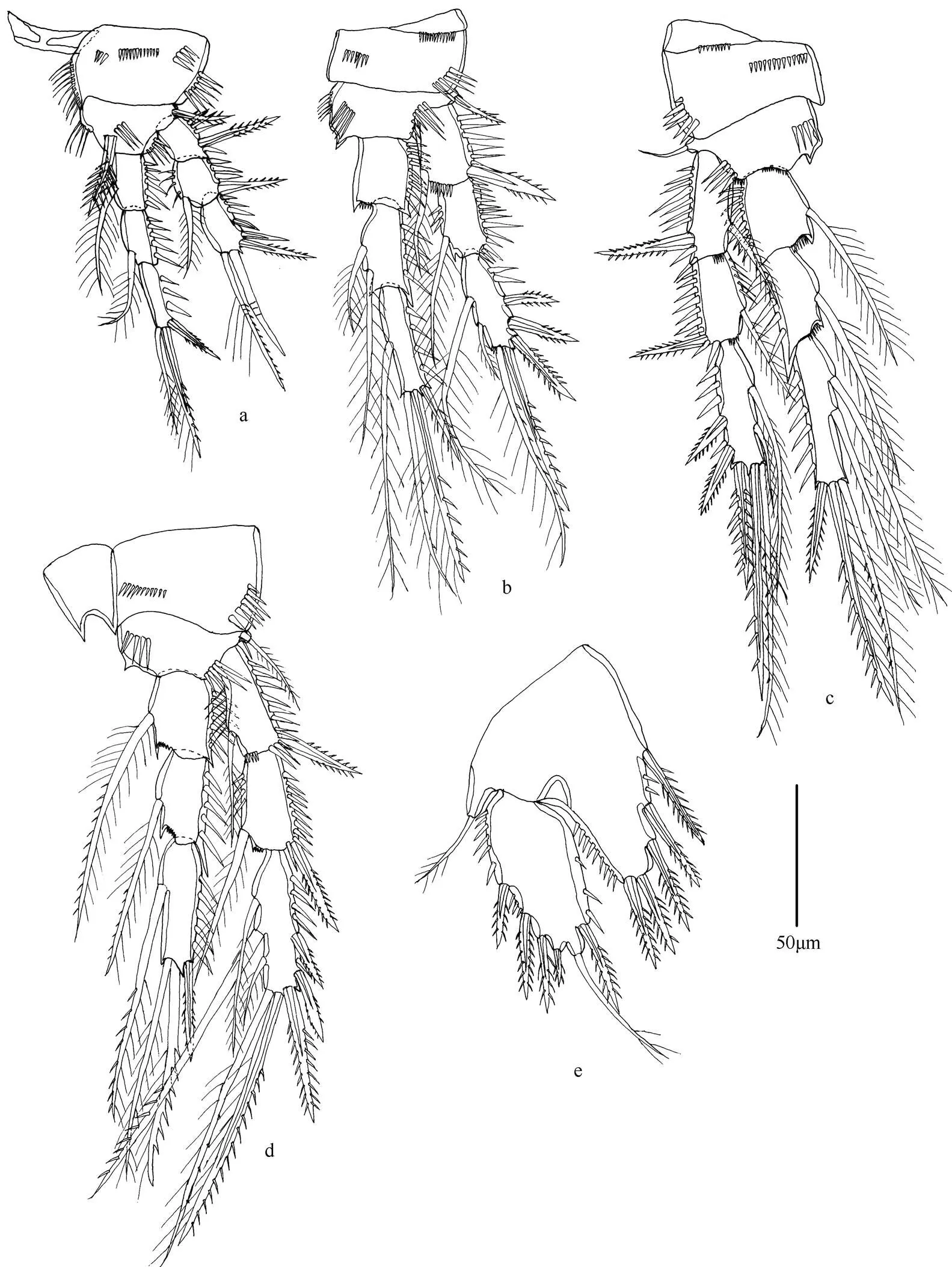
图6 显著华疑囊猛水蚤(雌性)
注: a. 第1胸足; b. 第2胸足; c. 第3胸足; d. 第4胸足; e. 第5胸足; 比例尺为50µm。

图7 田园威伦狭腹猛水蚤(雌性)整体图
雌性第5胸足内外肢愈合, 基节具3根刚毛, 内侧两根刚毛与外侧刚毛之间具有较大的空隙; 外肢具5根刚毛。
讨论 本种为中国海域首次记录, 身体外部形态、附肢刚毛和刺的分布情况与Karanovic 和Kim(2014)对韩国海域标本的描述一致, 无明显变化。目前此种仅在韩国海域和胶州湾有记录。
3 结论
本文通过对胶州湾采集的沉积物进行分析, 鉴定粗毛猛水蚤科5属5种,发现1新记录种田园威伦狭腹猛水蚤(Karanovic & Kim K., 2014)。目前, 胶州湾海域已鉴定到种水平的粗毛猛水蚤有6种。本文通过对已知种的形态特征分析, 制定了胶州湾粗毛猛水蚤科的种的检索表(见文末附表), 为以后的鉴定工作提供了便利条件。胶州湾海域的猛水蚤物种组成研究还不充分, 一些种因为样品量不足, 无法准确鉴定, 还需要进一步补充采集。
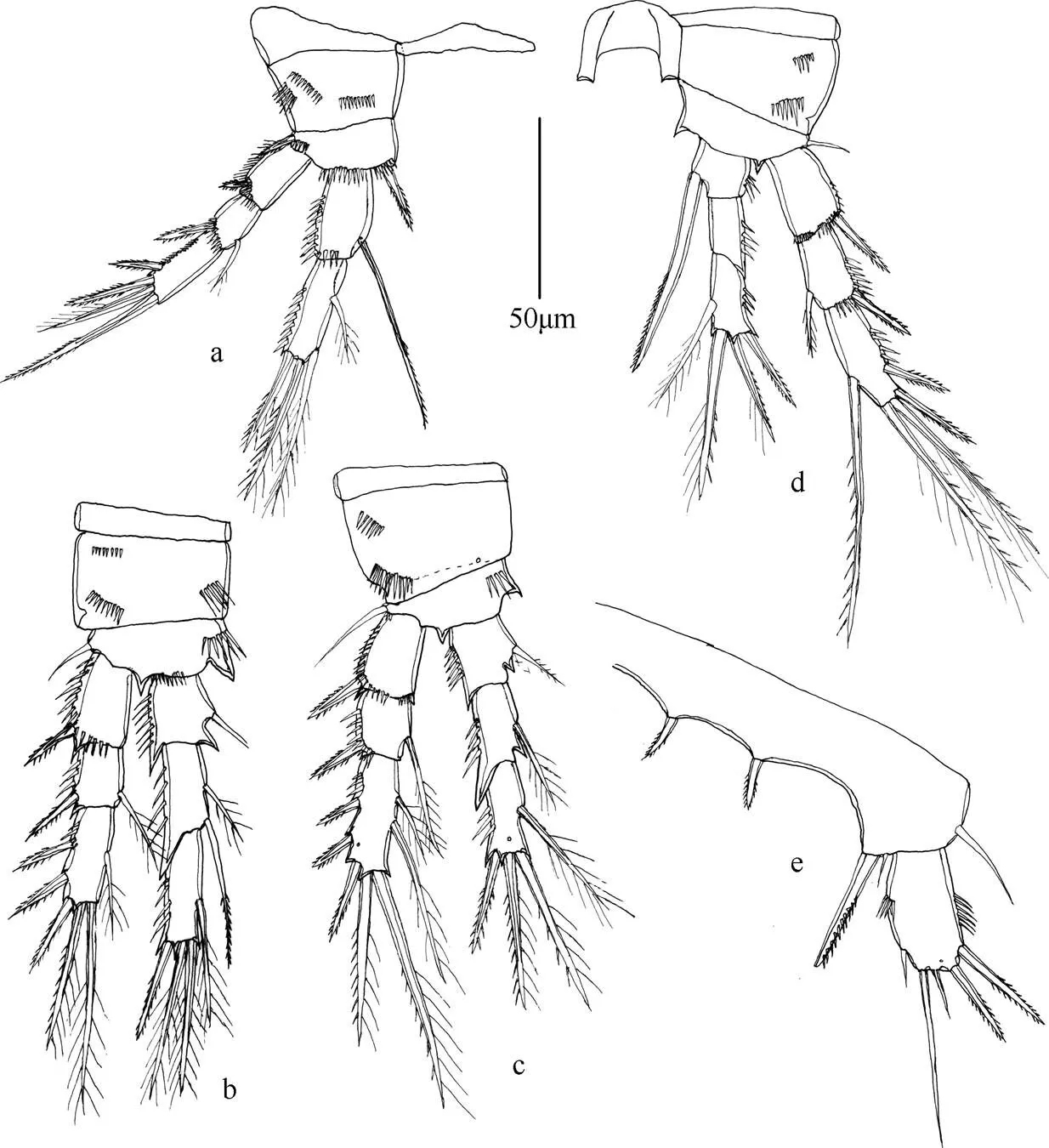
图8 田园威伦狭腹猛水蚤(雌性)
注: a. 第1胸足; b. 第3胸足; c. 第2胸足; d. 第4胸足; e. 第5胸足; 比例尺为50µm。
致谢 感谢课题组的各位同事在标本采集中提供的帮助。
Araújo-Castro C M V, Souza-Santos L P, Torreiro A G et al., 2009. Sensitivity of the marine benthic copepod(Copepoda, Harpacticoida) to potassium dichromate and sediment particle size., 57(1): 33—41
Boxshall G A, Halsey S H, 2004. An Introduction to Copepod Diversity. London: Ray Society, 966
Garlitska L, Neretina T, Schepetov D et al., 2012. Cryptic diversity of the ‘cosmopolitan’ harpacticoid copepod: genetic and morphological evidence. Molecular Ecology, 21(21): 5336—5347
Huys R, Mu F H, 2008. Description of a new species ofItô (Copepoda, Harpacticoida, Miraciidae) from the Bohai Sea, China. Zootaxa, 1706: 51—68
Itô T, 1972. Descriptions and records of marine harpacticoid copepods from Hokkaido, IV., 18(2): 305—336
Karanovic T, Kim K, 2014. New insights into polyphyly of the harpacticoid genus(Crustacea, Copepoda) through morphological and molecular study of an unprecedented diversity of sympatric species in a small South Korean bay. Zootaxa, 3783(1): 1—96
Kim K, Park E, Lee W, 2011. First record of(Copepoda: Harpacticoida: Miraciidae) from Korea. Bull. Nat. Inst. Biol. Resour., 2(2): 55—65
Lang K, 1965. Copepoda Harpacticoidea from the Californian pacific coastKungl. Svenska Vetenskapsakademiens Handlingar, 10(2): 1—566
Lee W, Park E, Song S J, 2012. Invertebrate Fauna of Korea, 21 (11), Marine Harpacticoida. South Korea: National Institute of Biological Resources, Ministry of Environment, 276
Ma L, Li X Z, 2011.sp. nov. (Harpacticoida, Miraciidae), a new copepod species from Jiaozhou Bay, Yellow Sea. Crustaceana, 84(9): 1085—1097
Moreno M, Vezzulli L, Marin Vet al.,2008. The use of meiofauna diversity as an indicator of pollution in harbours., 65: 1428—1435
Mu F H, Gee J M, 2000. Two new species of(Copepoda: Harpacticoida: Diosaccidae) and a related new genus, from the Bohai Sea, China. Cahiers de Biologie Marine, 41(2): 103—135
Rose A,Seifried S, Willen E et al., 2005. A method for comparing within-core alpha diversity values from repeated multicorer samplings, shown for abyssal Harpacticoida (Crustacea: Copepoda) from the Angola Basin. Organisms, Diversity & Evolution, 5(S1): 3—17
Somerfield P J, Gee J M, Aryuthaka C, 1998. Meiofaunal communities in a Malaysian mangrove forest. Journal of the Marine Biological Association of the United Kingdom, 78(3): 717—732
Song S J, Park J, Kwon B O et al., 2012. Ecological checklist of the marine and brackish-water harpacticoid copepod fauna in korean waters. Zoological Studies, 51(8): 1397—1410
Wells J B J, Rao G C, 1987. Littoral Harpacticoida (Crustacea: Copepoda) from Andaman and Nicobar islands. Memoirs of the Zoological Survey of India, 16(4): 1—385
Willen E, 2003. A new species of(Copepoda, Harpacticoida) from a hydrothermal, active, submarine volcano in the New Ireland Fore-Arc system (Papua New Guinea) with notes on deep sea colonisation within the Stenheliinae. Journal of Natural History, 37(14): 1691—1711
附:
种的检索表
1. 第2~4胸足内肢分2节···························· 双刺爪狭腹猛水蚤Huys & Mu, 2008
第2~4胸足内肢分3节·········································································································· 2
2. 大颚内肢长于外肢, 并向外肢弯曲, 具1根长刚毛, 长度至少大于外肢长度的四倍; 雌性具2个卵囊········ 3
大颚与上述特征不同·············································································································· 5
3. 第4胸足外肢末端具8根刚毛和刺············ 青岛韦氏狭腹猛水蚤(Ma & Li, 2011)
第4胸足外肢末端不多于7根刚毛和刺······················································································ 4
4. 第4胸足外肢末端具7根刚毛和刺·····························································································
·················································· 颂歌伊藤狭腹猛水蚤Karanovic & Kim K., 2014
第4胸足外肢末端具5根刚毛和刺·····························································································
····················································· 田园威伦狭腹猛水蚤Karanovic & Kim K., 2014
5. 第1胸足执握状, 内肢第1节长于第2节和第3节长度之和·····························································
······························································· 羽球疑囊猛水蚤Mu & Gee, 2000
第1胸足非执握状, 内肢第1节与第2节等长···············································································
····························································· 显著华疑囊猛水蚤Mu & Gee, 2000
Five Species of the Family Miraciidae (Copepoda) from the Jiaozhou Bay
MA Lin1, 2, 3LI Xin-Zheng1, 2, 3*
(1. Department of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Laboratory of Marine Biology and Biotechnology, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China)
Miraciidae Dana, 1846 is the second largest family in the Order Harpacticoida. Miraciids are common inhabitants of the marine benthos, and can be found in different types of sediments. Five species belonging to five genera were recognized morphologically,based on specimens obtained from the Jiaozhou Bay in the year 2008.Karanovic & Kim K., 2014 was first reported in China. A key to species of the family Miraciidae in Jiaozhou Bay was also given in the paper.
Miraciidae; Harpacticoida; Jiaozhou Bay
Q959
10.12036/hykxjk20170822003
国家自然科学基金(41206148); 中国科学院海洋研究所“一三五”专项资助项目(2012IO060104; 青岛海洋科学与技术国家实验室鳌山科技创新计划项目(No. 2015ASKJ01))。马 林, 助理研究员, E-mail: malin@qdio.ac.cn
李新正, 博士生导师, 研究员, E-mail: lixzh@qdio.ac.cn
2017-08-22,
2017-09-04
猜你喜欢
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
新农民(2020年15期)2020-06-22 07:38:58
东方企业家(2018年8期)2018-08-22 05:44:08
中国环境科学(2015年7期)2015-08-30 00:18:12
湖南农业(2015年7期)2015-03-20 01:48:37
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:20:57
华南农业大学学报(2012年2期)2012-11-10 08:21:16
台州学院学报(2011年3期)2011-01-13 03:45:52
