
基于CBM-CFS3模型的迪庆州云杉林地上生物量碳动态研究
2018-01-05 06:07皇宝林毕艳玲温庆忠邓喜庆朱建华肖文发
林业调查规划 2017年5期
皇宝林,毕艳玲,温庆忠,邓喜庆,冯 源,朱建华,4,肖文发,4
(1.云南省林业调查规划院,云南 昆明 650051;2.中国林业科学研究院森林生态环境与保护研究所,北京 100091;3.国家林业局森林生态重点实验室,北京 100091;4.南京林业大学南方现代林业协同创新中心,江苏 南京 210037)
基于CBM-CFS3模型的迪庆州云杉林地上生物量碳动态研究
皇宝林1,毕艳玲1,温庆忠1,邓喜庆1,冯 源2,3,朱建华2,3,4,肖文发2,3,4
(1.云南省林业调查规划院,云南 昆明 650051;2.中国林业科学研究院森林生态环境与保护研究所,北京 100091;3.国家林业局森林生态重点实验室,北京 100091;4.南京林业大学南方现代林业协同创新中心,江苏 南京 210037)
为明确区域尺度森林地上生物量碳储量动态特征,采用森林资源规划设计调查数据、解析木数据和加拿大林业碳收支模型(CBM-CFS3),以云南省迪庆州云杉林为例进行研究。2005—2016年迪庆州云杉地上生物量碳储量逐渐增大,范围为9.56~10.69Tg C,碳密度平均值为65.20Mg C/hm2。树干、其他木(含树皮)碳储量逐渐增加而树叶碳储量逐渐减小,三者碳储量范围为6.81~7.81Tg C、2.12~2.28Tg C和0.60~0.63Tg C;树干、其他木和树叶碳密度平均值分别为47.08 Mg C/hm2、14.17 Mg C/hm2和3.95 Mg C/hm2。地上生物量碳储量年均固碳速率为0.10Tg C/a,表明未来迪庆州云杉地上生物量碳库仍有较大的固碳潜能。
生物量;碳收支;碳储量;碳密度;CBM-CFS3模型;迪庆州
森林是陆地生态系统的主体,其地上生物量占陆地生态系统地上部分的80%[1]。生物量是生态系统能量和物质来源,对衡量森林生产力和物质循环、评价生态系统结构与功能具有的重要意义[2]。森林地上生物量是生物量中碳储量最大、最具动态特征且最易于调查和监测的部分,是森林生态系统整体碳储量估算的基础和关键。因此,地上生物量始终是研究森林碳储量与碳收支的重点。
在研究区域尺度森林生态系统生物量及碳储量时,最常用的方法是基于森林资源清查数据的生物量转换因子法(BEF法)和相对生长法[3]。BEF法即通过生物量转换因子将森林蓄积量转换为生物量,再通过木材密度、根茎比和含碳率等参数转换为碳储量。将多期森林资源清查数据估算结果进行比较,即可得到森林生物量碳动态[4-6]。相对生长法是通过建立全林(或各组分)生物量与平均胸径和树高的关系(异速生长方程),再根据研究区域面积、林分密度以及生物量含碳率得到碳储量结果[8-11]。这2种方法优点在于技术简单、结果明确,可快速估算出区域尺度森林生物量碳储量。其缺点在于森林资源清查5年进行一次,观测周期长,因此使用多期清查数据和BEF法的估算结果无法知悉每年森林生物量碳动态;而相对生长法属于静态估算,仅能反映数据当年的森林生物量碳储量,不能显示随林龄变化的碳动态。
加拿大林业碳收支模型(CBM-CFS3)能够克服上述2种方法的缺点。该模型属于经验模型,是加拿大国家森林碳监测、计量和报告系统(NFCMARS)的核心模块[12-14]。它使用IPCC-GPG所推荐的“清单+变化法”估算森林动态,即基于森林清查数据先估算出森林每年净生长量,再模拟生态进程、人为活动及自然干扰事件对森林碳储量的影响,是IPCC推荐使用的最高层(Tier 3)的碳计量方法[13]。CBM-CFS3模型研究尺度灵活、时间步长为1 a,可估算林分到国家尺度的森林生态系统年际碳动态。本文使用CBM-CFS3模型框架,以云南省森林资源清查数据、迪庆州森林资源规划设计调查数据以及云杉解析木为主要数据源,生成输入数据并修改模型默认参数,模拟云杉面积不变、自然生长条件下迪庆州云杉地上生物量碳动态。
1 研究区域概况
云南省迪庆藏族自治州(98°35′~100°19′E,26°52′~29°16′N)位于滇、川、藏三省区交汇处的横断山脉腹地,总面积为 23 870 km2。境内地势北高南低,地形呈纵深切割之势,海拔高差悬殊。年均温4.7~16.5℃,年日照时间 1 740~2 190 h,年降雨量600~1 000 mm。迪庆州是云南省森林植被类型最丰富的区域之一,林业用地面积1.62×106hm2,森林覆盖率达73.9%,高于全省平均水平。云杉是迪庆州主要森林类型之一,根据2005年云南省迪庆州森林资源规划设计调查数据,云杉森林面积为1.55×105hm2,占迪庆州森林总面积的10.42%。迪庆州云杉分布如图1所示。
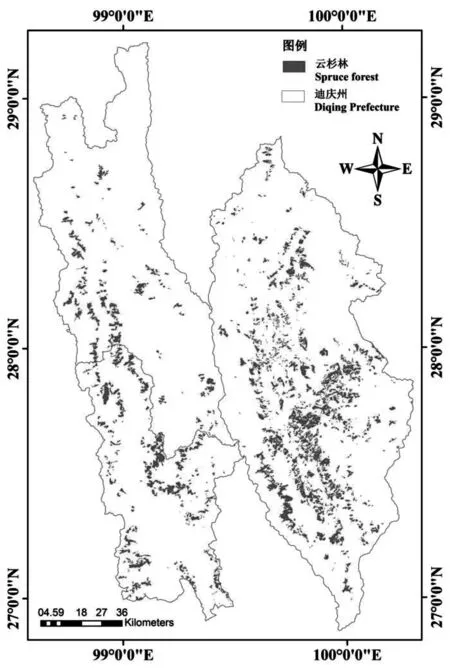
图1 2005年迪庆州云杉分布示意Fig.1 Distribution of spruce forest in Diqing at 2005
2 研究方法
CBM-CFS3是由生长曲线数据驱动的经验模型,包括生物量和死亡有机质(DOM)两大碳库系统,生物量碳库中共有树干、其他木(树枝与树皮之和)和树叶3个地上生物量碳库[13, 15]。在不考虑自然干扰和人为经营情况下,森林生物量碳储量为每年净生长量的累积。该模型输入数据来源于2005年迪庆州森林资源规划设计调查数据中云杉树种组6 130条小班记录。而模型参数主要包括区域尺度蓄积生长方程、蓄积—干材生物量转换方程和生物量组分比例参数,分别由森林资源清查数据、森林资源规划设计调查数据以及云杉解析木数据得到。生物量估算技术路线见图2,估算方程见表1。由于森林资源清查中不调查胸径5 cm以下的林木,而且不测量灌丛和草本。因此,本文以迪庆州云杉林乔木层地上生物量为研究对象,不包括胸径5cm以下的幼树、灌木和草本生物量。
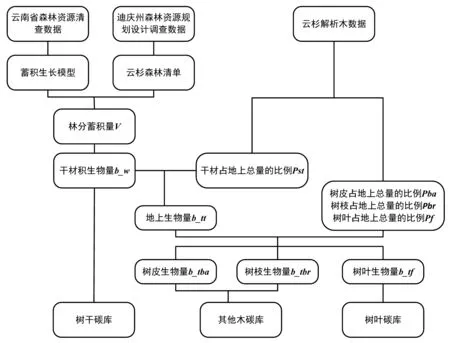
图2 研究流程Fig.2 Flow chart of study
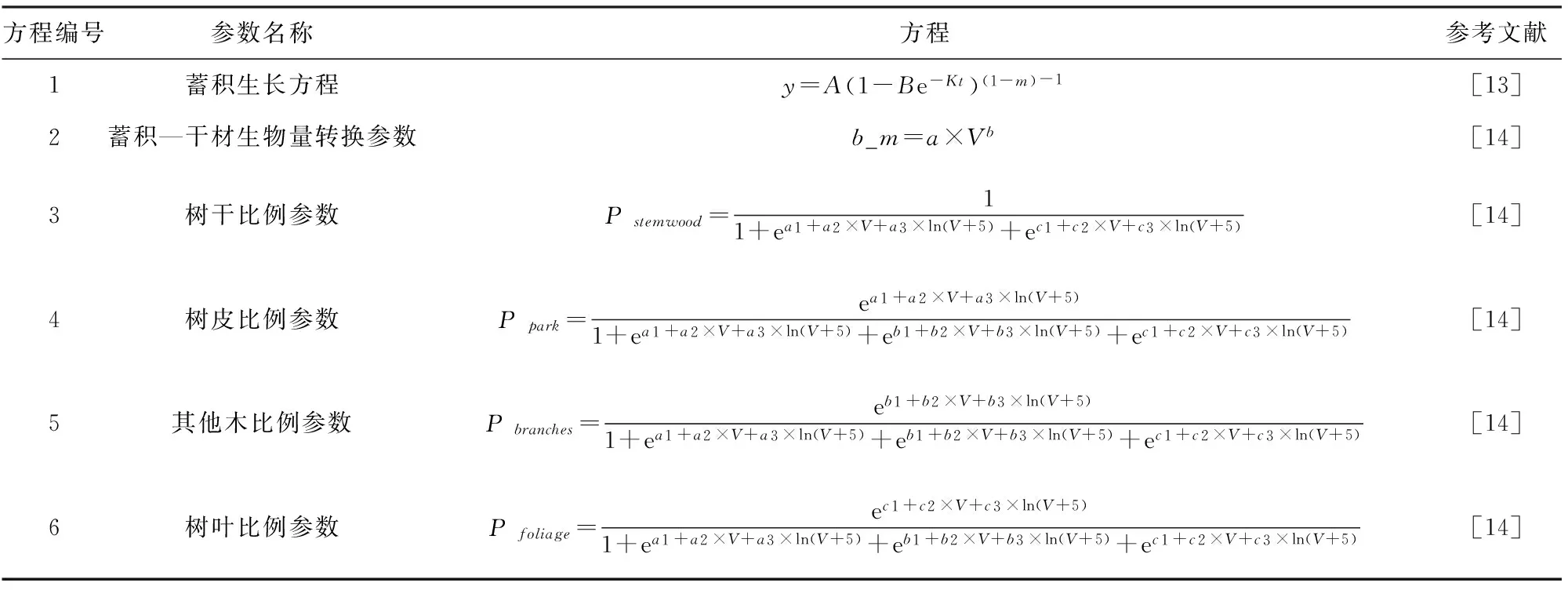
方程编号参数名称方程参考文献1蓄积生长方程y=A(1-Be-Kt)(1-m)-1[13]2蓄积—干材生物量转换参数b_m=a×Vb[14]3树干比例参数Pstemwood=11+ea1+a2×V+a3×ln(V+5)+ec1+c2×V+c3×ln(V+5)[14]4树皮比例参数Ppark=ea1+a2×V+a3×ln(V+5)1+ea1+a2×V+a3×ln(V+5)+eb1+b2×V+b3×ln(V+5)+ec1+c2×V+c3×ln(V+5)[14]5其他木比例参数Pbranches=eb1+b2×V+b3×ln(V+5)1+ea1+a2×V+a3×ln(V+5)+eb1+b2×V+b3×ln(V+5)+ec1+c2×V+c3×ln(V+5)[14]6树叶比例参数Pfoliage=ec1+c2×V+c3×ln(V+5)1+ea1+a2×V+a3×ln(V+5)+eb1+b2×V+b3×ln(V+5)+ec1+c2×V+c3×ln(V+5)[14]
2.1 区域尺度云杉蓄积生长方程拟合及检验
将云南省1987—2012年森林资源清查中云杉优势树种的固定样地监测数据作为蓄积生长方程拟合样本。剔除异常数据后云杉林龄范围为0~265 a,蓄积量范围为0~664.3 m3,随机选取80%样本作为建模样本(64个),剩下20%样本为检验样本(16个)。使用DPS 13.0统计软件非线性Marquardt迭代法按方程1建立林分蓄积量随林龄变化的关系(图3a),以调整后的相关指数(R2)、均方根误差(RMSE)评价拟合效果(表2)。蓄积方程建立后将模拟值与检验样本的实测值进行对比(图3b),以回归系数b(即回归方程斜率)和R2作为方程适用性检验标准(表2)。

图3 区域尺度云杉蓄积生长方程拟合及方程适用性检验(b)Fig.3 Fitting and equation test of regional-scale yield curve of spruce
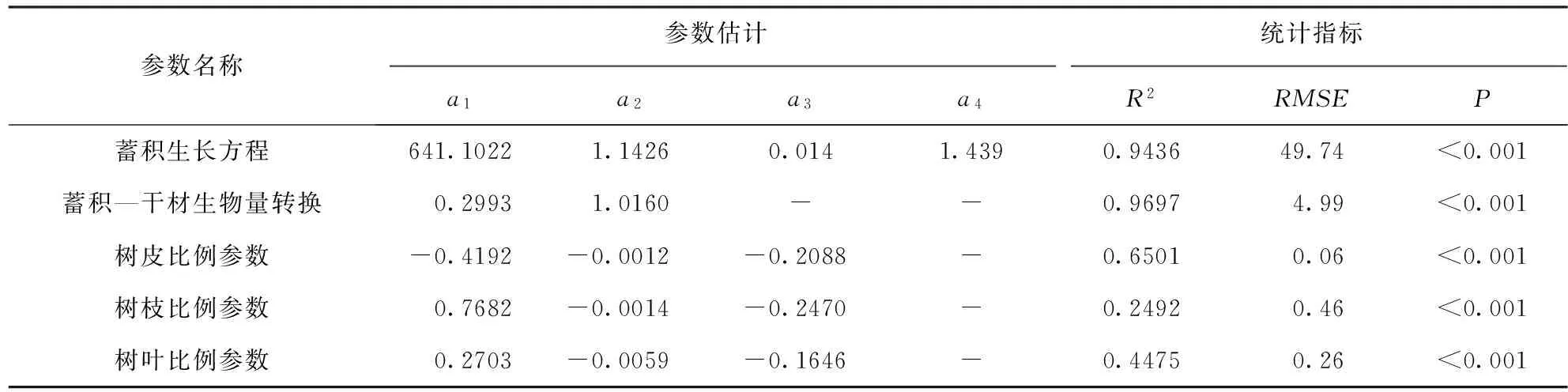
参数名称参数估计统计指标a1a2a3a4R2RMSEP蓄积生长方程641.10221.14260.0141.4390.943649.74<0.001蓄积—干材生物量转换0.29931.0160--0.96974.99<0.001树皮比例参数-0.4192-0.0012-0.2088-0.65010.06<0.001树枝比例参数0.7682-0.0014-0.2470-0.24920.46<0.001树叶比例参数0.2703-0.0059-0.1646-0.44750.26<0.001
由表2可以看出,云杉蓄积生长方程a1值为641.102 m3/hm2,即云杉林分蓄积量极限值。a2值为1.143,表示林分达到一定林龄后才有蓄积。a3和a4与同化速率有关,共同决定曲线拐点及最大生长速率。方程R2为 0.943 6,说明林龄可解释林分蓄积生长变化的94%以上,拟合效果好;RMSE为49.74,说明所建模型与样本数据契合程度较高,并且方程极显著(P<0.001)。由图3b可知,b值为1.015、R2为0.946,并且方程通过了显著性检验,满足检验标准,即所拟合的云杉蓄积生长方程具有良好的适用性。
2.2 蓄积—干材生物量转换方程及生物量组分比例参数的拟合
蓄积—干材生物量转换方程及生物量组分比例参数拟合数据主要来源于云南省林业调查规划院为建立云杉生物量模型而采集的152株云杉解析木。即将云南全省范围内云杉按胸径2~38 cm以上划分10个径阶,在各径阶中根据树高不同均匀抽取15个样本。共采集152个样本,样本概况见表3。记录样本所在林分的林龄、林分密度及土壤、地形等环境因子,实测样本胸径、树高、枝下高、冠幅等因子。利用分层分割法分别对解析木干材、干皮、其他木及树叶进行取样,称量鲜重。将样本在85℃恒温下烘干到恒重,得到各类样本的干重率。将云杉解析木各组分鲜重与测得的干重率相乘计算出单株干材、树皮、树枝和树叶的生物量。使用平均木法将样本单株及各组分生物量转换为林分水平生物量。
根据Boudewyn等[16]提供的方程形式(表1方程2~6),使用DPS 13.0软件拟合出蓄积—干材生物量转换参数以及各组分占地上生物量的比例参数,以均方根误差(RMSE)评价模拟效果。将拟合出的蓄积—干材生物量转换参数和各组分生物量参数替换模型默认参数,拟合曲线见图4。CBM-CFS3模型计算出各组分生物量之后,将树枝和树皮归入同一个碳库中,即其他木碳库(Other wood pool)[13],因此这个碳库对应2种组分生物量。
由表2可知,蓄积—干材生物量转换方程R2高达 0.969 7,反映出蓄积与生物量间的极强相关性,拟合效果较好。不同生物量组分中,树皮R2值最高为0.650 1,拟合效果最好;其次是其他木和树叶,其他木R2最小,为0.249 2。所有组分拟合方程均达到极显著水平,使用这些参数可较好地模拟出迪庆州云杉生物量。

表3 云杉解析木样本概况
3 结果分析
3.1 迪庆州云杉地上生物量碳动态
地上生物量包含树干、其他木和树叶3个子碳库。2005—2016年地上生物量碳库中树干碳储量最大,其他木碳储量其次,而树叶碳储量最小。树干碳储量逐年增大,范围为6.81~7.81Tg C,年均固碳速率为0.09Tg C/a;其他木碳库的碳储量也逐年增大,范围为2.12~2.28Tg C,年均固碳速率为0.02Tg C/a;而树叶碳储量逐渐减小,范围为0.60~0.63Tg C,平均值为0.61Tg C。树干和其他木碳库的碳储量增加大于树叶碳储量的减小,因而地上生物量碳储量表现为逐年增加,地上生物量碳储量范围为9.56~10.69Tg C,年均固碳速率为0.10Tg C/a(表4)。
树干碳密度最大,树叶碳密度最小。2005—2016年间树干碳密度逐渐增大,范围为43.86~50.28 Mg C/hm2,平均值为47.08Mg C/hm2,年均固碳速率为0.58 Mg C/hm2;其他木碳密度逐渐增大,范围为13.62~14.70 Mg C/hm2,平均值为14.17 Mg C/hm2,年均固碳速率为0.10 Mg C/hm2;树叶碳密度逐年减小,范围为3.86~4.04 Mg C/hm2,平均值为3.95 Mg C/hm2;地上生物量碳密度逐渐增大,范围为61.53~68.84Mg C/hm2,平均值为65.20 Mg C/hm2,年均固碳速率为0.66 Mg C/hm2(图5)。
3.2 模拟结果对比验证
使用生物量转换因子法(BEF法)估算2005年迪庆州云杉地上生物量碳储量,即基于迪庆州资源规划设计调查数据和中国温室气体清单研究[17]提供的云杉不同龄组生物量扩展因子(BEF)、基本木材密度(SVD)与含碳率(CF)来进行估算,并将结果与CBM模拟结果进行对比(图6)。用BEF法估算2005年迪庆州云杉地上生物量碳储量为9.25 Tg C,CBM-CFS3估算结果为9.56Tg C,可见2种方法对云杉地上生物量估算值极为接近,仅相差0.31 Tg C,说明本地化后的模型参数数值合理。
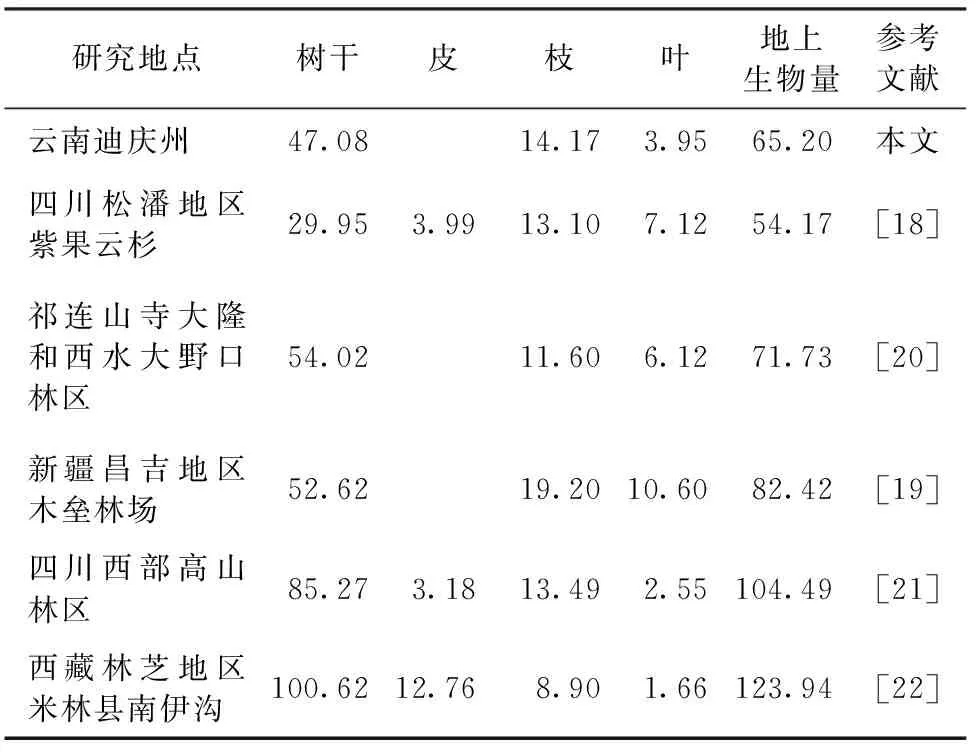
将本研究结果与已发表的云杉生物量研究结果进行对比(表5),迪庆州云杉地上生物量碳密度平均值(65.20Mg C/hm2)大于江洪[18]研究结果(54.17 Mg C/hm2),但小于王燕[19]、王金叶[20]、马明东[21]和方江平[22]得出的碳密度值。生物量碳密度与树种生长特性、平均林龄和立地环境有密切关系;估算结果也因数据来源、研究方法不同而产生较大差异。本研究结果在前人结论范围内,即CBM-CFS3对区域尺度云杉地上生物量碳动态的估算合理可信。
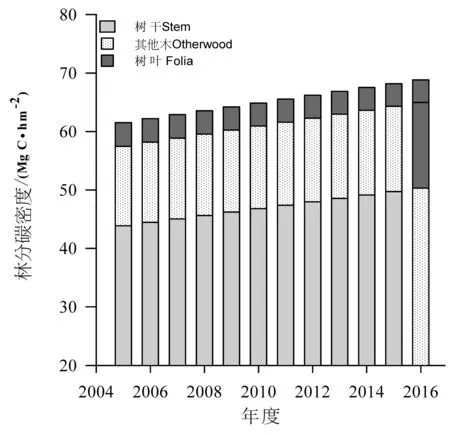
图5 迪庆州云杉地上生物量碳密度动态Fig.5 Dynamic of above-ground biomass carbon density of spruce forest in Diqing
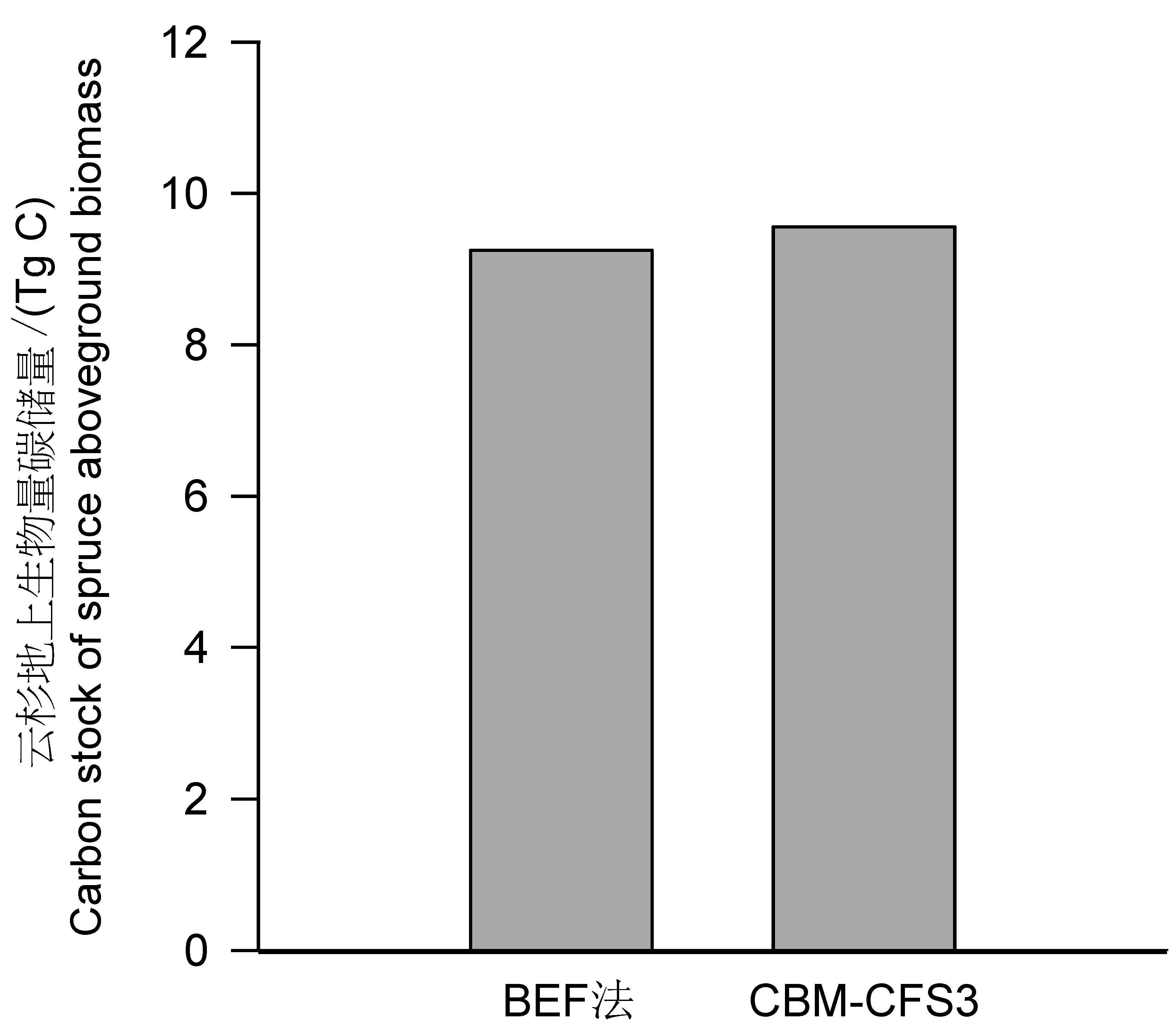
图6 云杉地上生物量碳储量的2种估算方法结果对比Fig.6 Comparison of above-ground biomass carbon stock estimated by two methods
/(Mg C·hm-2)
注:若文献结果为生物量(t/hm2),按含碳率为0.5将其转化为生物量碳密度(Mg C/hm2)
4 讨论
区域尺度云杉蓄积生长方程是按Richard方程形式将云南全省云杉固定样地作为样本拟合得到方程,具有良好的生物学意义[23-24],相关指数高达0.943 6,并且通过显著性检验和适用性检验,模拟效果好。该方程适于估算云南区域尺度云杉林分蓄积生长,并且可为森林资源更新提供数据支持。
CBM-CFS3模型中生物量组分参数拟合方程与唐守正等人提出的相容性生物量方程[25-28]相似,主要区别在于方程形式不同以及生物量组分的标准设置不同,而多项对数方程拟合过程简单,能够有效估算地上生物量及其组分碳动态。
将BEF法估算结果与参数修改后的CBM-CFS3模拟结果对比,二者估算值极为接近,同时云杉地上生物量碳密度也在前人文献结果范围内,表明云杉碳储量和碳密度估算结果合理并且CBM-CFS3模型适于估算迪庆地区森林生物量碳动态。相较于收获法以及IPCC第二层次生物量转换因子法,CBM-CFS3是更为系统的碳计量工具,可估算多种尺度森林碳储量动态变化。
2005—2016年,迪庆州云杉地上生物量总体、树干和其他木碳库储量均逐渐增加,而树叶碳库逐渐减小,这与生长过程中云杉地上生物量年增量逐渐减小、树叶所占总生物量的比例逐渐减少有关,反映出云杉地上生物量的碳分配规律。林下幼树、灌草是森林生物量的重要组分,文献结果表明,幼树、灌草生物量之和占乔木层地上生物量比例的6.65%~17.83%[18,20-22];方江平[22]指出,幼树及灌木、草本生物量之和高达16.74Mg C/hm2。由于我国森林调查仅针对胸径大于5 cm的乔木层林木,较少关注幼树、灌木及草本,因此以森林调查数据驱动的CBM-CFS3模型不能估算出林下层生物量,将低估区域尺度生态系统碳储量。研究表明,增加林下灌木、草本甚至苔藓的生物量估算,能有效降低区域尺度上森林碳储量估算的不确定性[22, 29]。未来需要加强对林下层生物量研究,明确其碳储量及变化规律,可使用扩展因子与乔木层生物量相乘的方式进行估算。
本研究所得结果是在假设2005—2016年迪庆州云杉面积不增减且无重大自然干扰和人为影响的情况下得出,然而实际上由于造林或毁林等措施,使得迪庆州云杉林面积不可避免地发生变化,采伐也会导致地上生物量的直接损失和转移,因此本文结果是一种理想状态的估算,未来需要量化土地利用变化和人为干扰对迪庆州云杉森林碳储量的影响,更加精确地估算森林碳动态。
[1] MALHI Y, BALDOCCHI D D, JARVIS P G. The carbon budget of tropical, temperate and boreal forests [J]. Plant, Cell & Environment, 1999, 22(6): 715-740.
[2] STEPHENSON N L, DAS A J, CONDIT R, et al. Rate of tree carbon accumulation increases continuously with tree size[J]. Nature, 2014, 507(7490): 90-93.
[3] 方精云,刘国华,徐篙龄. 我国森林植被的生物量和净生产量[J].生态学报,1996,16(5):497-508.
[4] 赵敏,周广胜. 中国森林生态系统的植物碳贮量及其影响因子分析[J]. 地理科学,2004,24(1): 50-54.
[5] LEHTONEN A, M?KIP?? R, HEIKKINEN J, et al. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests[J]. Forest Ecology and Management, 2004, 188(1): 211-224.
[6] 李海奎, 赵鹏祥, 雷渊才, 等. 基于森林清查资料的乔木林生物量估算方法的比较[J]. 林业科学, 2012, 48(5): 44-52.
[7] 郭兆迪, 胡会峰, 李品, 等. 1977—2008 年中国森林生物量碳汇的时空变化[J]. 中国科学·生命科学, 2013, 43(5): 421-431.
[8] 陈庆, 郑征, 冯志立, 等. 云南普洱地区思茅松林生物量及碳储量研究[J]. 云南大学学报(自然科学版), 2014, 36(3): 439-445.
[9] 曾鸣, 聂祥永, 曾伟生. 中国杉木相容性立木材积和地上生物量方程[J]. 林业科学, 2013, 49(10): 74-79.
[10] 胡海清, 罗碧珍, 魏书精, 等. 小兴安岭 7 种典型林型林分生物量碳密度与固碳能力[J]. 植物生态学报, 2015, 39(2): 140-158.
[11] 姜鹏, 张绍轩, 任佳佳, 等. 木兰围场典型落叶松—杨桦混交林生物量及固碳能力[J]. 生态学报, 2015, 35(9): 2937-2945.
[12] KURZ W A,APPS M J. Developing Canada′s national forest carbon monitoring, accounting and reporting system to meet the reporting requirements of the Kyoto Protocol[J].Mitigation and Adaptation Strategies for Global Change,2006,11(1):33-43.
[13] KURZ W A,DYMOND C C,WHITE T M, et al.CBM-CFS3: a model of carbon-dynamics in forestry and land-use change implementing IPCC standards[J].Ecological Modelling,2009,220(4):480-504.
[14] KULL S J, RAMPLEY G J, MORKEN S, et al. Operational-scale carbon budget model of the Canadian forest sector (CBM-CFS3) version 1.2: user′s guide[M]. Edmonton, Natural Resources Canada, 2014:1-17.
[15] STINSON G,KURZ W A, SMYTH C E, et al. An inventory-based analysis of Canada′s managed forest carbon dynamics, 1990 to 2008[J].Global Change Biology,2011,17(6):2227-2244.
[16] BOUDEWYN P, SONG X, MAGNUSSEN S, et al. Model-based, volume-to-biomass conversion for forested and vegetated land in Canada[M].Victroia: Pacific Forestry Centre, 2007: 2-12.
[17] 国家发展和改革委员会应对气候变化司. 2005中国温室气体清单研究[M]. 北京:中国环境出版社,2014.
[18] 江洪. 紫果云杉天然中龄林分生物量和生产力的研究[J]. 植物生态学与地植物学丛刊, 1986(2): 1-8.
[19] 王燕, 赵士洞. 天山云杉林生物量和生产力的研究[J]. 应用生态学报, 1999, 10(4): 389-391.
[20] 王金叶, 车克钧. 祁连山青海云杉林碳平衡研究[J]. 西北林学院学报, 2000, 15(1): 9-14.
[21] 马明东, 江洪, 罗承德, 等. 四川西北部亚高山云杉天然林生态系统碳密度、净生产量和碳贮量的初步研究[J]. 植物生态学报, 2007, 31(2): 305-312.
[22] 方江平. 西藏南伊沟林芝云杉林生物量与生产力研究[J]. 林业科学研究, 2012, 25(5): 582-589.
[23] 刘建敏. 基于小班数据库的忻城县森林优势树种组生长模型研究[J]. 广西科学, 2011, 18(3):294-297
[24] CABELLA B C T, RIBEIRO F, MARTINEZ A S. Effective carrying capacity and analytical solution of a particular case of the Richards-like two-species population dynamics model[J]. Physica A: Statistical Mechanics and its Applications, 2012, 391(4): 1281-1286.
[25] 张会儒,赵有贤. 应用线性联立方程组方法建立相容性生物量模型研究[J]. 林业资源管理,1999 (6): 63-67.
[26] 唐守正,张会儒,胥辉. 相容性生物量模型的建立及其估计方法研究[J]. 林业科学, 2000, 36(1):19-27
[27] 胥辉,刘伟平. 相容性生物量模型研究[J]. 福建林学院学报,2001,21(1):18-23.
[28] 符利勇,雷渊才,孙伟, 等. 不同林分起源的相容性生物量模型构建[J]. 生态学报,2014,34(6):1461-1470.
[29] BONA K A, FYLES J W, SHAW C, et al. Are Mosses Required to Accurately Predict Upland Black Spruce Forest Soil Carbon in National-Scale Forest C Accounting Models[J]. Ecosystems, 2013, 16(6): 1071-1086.
Carbon Dynamic of Above-ground Biomass of Spruce Forest in Diqing Tibetan Autonomous Prefecture Based on CBM-CFS3 Model
HUANG Baolin1;BI Yanling1; WEN Qingzhong1;DENG Xiqing1;FENG Yuan2,3;ZHU Jianhua2,3,4;XIAO Wenfa2,3,4
(1.Yunnan Forest Inventory and Planning Institute, Kunming 650051,China; 2.Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry , Beijing 100091,China; 3. Key Laboratory of Forest Ecology and Environment of State Forestry Administration , Beijing 100091, China; 4. Co-Invovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China)
This study utilized Carbon Budget Model of the Canadian Forest Sector (CBM-CFS3) to quantify the regional-scale forest biomass carbon dynamic by the case of the spruce forest in Diqing Tibetan Autonomous Prefecture of Yunnan, with the data source of Forest Inventory for Planning and Designing and analytical trees. From 2005 to 2016, above-ground biomass carbon stock of spruce forest in Diqing increased gradually from 9.56 to 10.69Tg C, with the average carbon density of 65.20 Mg C ha-1. The carbon stock of stem and branch (including bark) increased but the carbon stock of folia declined annually, ranges of which were 6.81-7.81Tg C, 2.12-2.28Tg C and 0.60-0.63Tg C respectively, and average carbon density of which were 47.08Mg C ha-1, 14.17Mg C ha-1and 3.95 Mg C ha-1respectively. The annual carbon sequestration rate (0.10Tg C a-1) suggested that spruce forest in Diqing had the potential ability to sequestrate enormous carbon in the future..
biomass; carbon budget; carbon stock; carbon density; CBM-CFS3 model; Diqing Tibetan Autonomous Prefecture
2017-03-13.
皇宝林(1976-),男,云南保山人,工程师.从事林业调查规划及软件开发工作.
10.3969/j.issn.1671-3168.2017.05.001
S791.18;S718.556;S711
A
1671-3168(2017)05-0001-08
猜你喜欢
香格里拉(2022年2期)2022-08-16
云南画报(2020年12期)2021-01-18
现代农村科技(2020年7期)2020-12-19
矿产综合利用(2020年1期)2020-07-24
花卉(2020年6期)2020-01-07
乡村科技(2019年14期)2019-02-21
启迪与智慧·教育版(2018年12期)2018-02-26
中成药(2017年7期)2017-11-22
中国民族医药杂志(2016年8期)2016-05-09
中国洗涤用品工业(2015年2期)2015-02-28
