
中华猕猴桃在中国潜在分布及其对气候变化响应的研究*
2018-01-04 01:43王茹琳何仕松
中国生态农业学报(中英文) 2018年1期
王茹琳, 李 庆, 何仕松, 刘 原
中华猕猴桃在中国潜在分布及其对气候变化响应的研究*
王茹琳1,2, 李 庆1**, 何仕松3, 刘 原3
(1. 四川农业大学农学院 成都 611130; 2. 四川省农村经济综合信息中心 成都 610072; 3. 四川苍溪猕猴桃研究所 苍溪 628400)
中华猕猴桃为中国特有果种, 由于其独特的口感和较高的经济价值, 近年来种植规模逐年扩大。在引种过程中, 由于缺乏合理的布局规划和适生性分析, 出现了品种单一化、易感病虫害等问题。近年来四川、陕西、贵州、重庆和湖北等猕猴桃主产省份相继开展了猕猴桃气候适宜性区划的研究, 但目前的研究多未考虑未来气候变化对猕猴桃种植分布的影响, 且伴随着气候变化的加剧, 已有的研究结果已不能完全适应实际生产的需求。本文运用生态位模型软件MaxEnt, 模拟和预测气候变化背景下大尺度范围中华猕猴桃适生区分布及其变化的可行性, 以利于科学地优化产业结构、促进产业发展。基于当前数据和IPCC AR5提出的3种气候情景以及中华猕猴桃的分布信息, 采用MaxEnt生态位模型和ArcGIS预测了中华猕猴桃的适生区及未来的变化趋势, 用受试者工作特征曲线(receiver operating characteristic curve, ROC曲线)检测模型精度、刀切法(Jackknife test)筛选主导环境变量。结果表明, 基于当前和未来情景构建的中华猕猴桃地理分布模型的AUC(area under curve)值均达到“极好”的标准, 说明模型预测结果可用于本研究。当前气候条件下, 中华猕猴桃的高适生区主要在四川、陕西、重庆、湖北、贵州、浙江、湖南、安徽、河南、江苏和甘肃等省份, 面积达1.01×106km2。中适生区则以高适生区为中心向外扩散, 包括河南、湖北、安徽、江苏和山东等地, 面积为6.79×105km2。RCP2.6和RCP4.5排放情景下, 中华猕猴桃高适生区的分布、面积及中心点位置都有所不同, 面积均呈增加趋势; RCP8.5排放情景下, 高适生区面积呈减少趋势。RCP4.5和RCP8.5排放情景下, 中华猕猴桃高适生区中心点均有向北移动趋势。MaxEnt模型对未来气候变化条件下中华猕猴桃适生区的准确模拟与预测具有潜在应用价值, 对该果树的气候适宜性区划具有重要指导意义。
中华猕猴桃; MaxEnt模型; 环境变量; 气候变化; 适生区分析; 典型浓度路径情景
生物与气候之间相互作用对物种的分布影响巨大, 气候变化不仅可以改变物种所在的生态系统, 且造成的改变往往是不可逆的。在区域尺度上, 气候是影响物种分布的主要环境因素[1]。农业生产强烈依赖气候生态条件, 气候变化将改变农作物已有的适宜性区划, 影响其生长、发育及产量[2-4]。物种分布模型 (species distribution models, SDM)是研究实际气候变化对物种分布影响的重要工具, 近年来该类模型在国内外得到广泛应用, 目前常用的有最大熵模型MaxEnt (maximum entropy model)、基于遗传算法的规则组合模型GARP(genetic algorithm for rule set produciton model)、生物气候模型BIOCLIM (biological climatic model)和气候动态模拟软件Climex (match climate and compare location model)等约14种物种分布模型[5-7]。MaxEnt模型的模拟精度要高于其他模型, 加之该软件具有运行时间短、操作简便、运行结果稳定和所需样本量小等特点在业内得到了广泛认可[8]。近年来, 国内学者利用MaxEnt模型成功研究了气候变化对水稻()[9]、玉米()[10]、小麦()[11]等粮食作物的影响, 并分析了气候因子与种植分布之间的关系, 取得了较好的模拟效果。
猕猴桃(spp.), 又称奇异果, 隶属于猕猴桃科(Actinidiaceae), 猴桃属, 为浆果类落叶藤本果树, 是20世纪人工驯化栽培野生果树最有成就的四大果树之一[12]。中国是世界猕猴桃起源中心, 猕猴桃属66个种中有62个种原产于中国[13]。近年来, 随着我国农业产业结构调整, 猕猴桃产业发展迅速, 猕猴桃的栽培面积和产量都得到长足的发展, 目前我国猕猴桃年产量已超过意大利, 成为世界第一产出国。中华猕猴桃()和美味猕猴桃()是我国栽培最广泛的两个种。中华猕猴桃为中国特有果种, 由于其口感独特、经济价值高, 近年来种植规模逐年扩大。在引种过程中, 由于缺乏合理的布局规划和适生性分析, 出现了品种单一化、易感病虫害等问题[14-15]。针对上述问题, 近年来四川[16]、陕西[17]、贵州[18]、重庆[19]和湖北[20]等省份开展了猕猴桃气候适宜性区划的研究, 并在实际生产中起到了积极作用。但目前的研究多未考虑未来气候变化对猕猴桃种植分布的影响, 且伴随着气候变化的加剧, 已有的研究结果已不能完全适应实际生产的需求, 因此有必要开展针对中华猕猴桃的气候适宜性研究, 以利于科学地优化产业结构、促进产业发展。本研究利用MaxEnt模型, 结合GIS技术, 模拟中华猕猴桃在中国的潜在分布范围, 预测、对比、分析中华猕猴桃在未来3种气候情景(RCP2.6、RCP4.5和RCP8.5)下分布范围和空间格局的变化规律, 旨在探索中华猕猴桃合理区划的有效手段, 以期为其生产布局及应对未来气候变化提供基本的数据支持。
1 材料与方法
1.1 预测软件来源
选用基于最大熵理论的MaxEnt模型软件来预测中华猕猴桃在中国的适生分布。登录普林斯顿大学Robert Schapire计算机科学研究中心的MaxEnt主页可免费下载该软件, 当前最新版本为3.3.3k版[21]。
选用ArcGIS软件将MaxEnt模型的运算结果投射到地图上显示, 预测中华猕猴桃的实际分布和潜在分布。
1.2 物种分布记录
运用生态位模型模拟物种适生区时[22-23], 首先应获取目标物种的实际分布数据, 本研究中四川境内中华猕猴桃的分布数据为实地考察获得, 采用GPS记录经纬度, 其他地区的分布数据则通过查询物种分布数据库和检索国内外公开发表的中华猕猴桃相关的期刊论文获得。本研究查询的数据库包括“国际农业与生物科学中心(CABI, http://www. cabi.org/)” 数据库、“全球物种多样性信息库(GBIF, http://www.gbif.org/)”和“教学标本资源共享平台”(http://mnh.scu.edu.cn/)。通过上述方式共获得分布点283个, 使用Google Earth软件查询分布点的经纬度信息, 按照MaxEnt软件要求, 去除重复记录、模糊记录和邻近记录的分布点。将最终确定的经纬度数据使用Excel处理, 保存格式为*.CSV[24]。
1.3 环境变量的选择
构建物种生态位模型, 还应选取合适的环境变量。本研究从worldclim网站下载了67个环境变量, 包括19个具有较强的生物学意义的生物气候变量(这些变量是以温度和雨量为基础数据, 根据不同需求计算演生而来的气候变量, 可反映温度与降水的特点及季节性变化特征)、月平均温度、月最高温度、月最低温度和月降水量。上述数据的空间分辨2.5arc-minutes(约4.5 km2)。当前情景的数据在worldclim网站下载。气候变化专门委员会(IPCC)第5次评估报告共发布的4种典型浓度路径[25](representative concentration pathways, RCP), 即RCP2.6、RCP4.5、RCP6.0 和 RCP8.5。其中RCP4.5和RCP6.0均为中等CO2排放情景, 且前者优先性大于后者, 因此本研究选择RCP2.6 (最低CO2排放情景)、RCP4.5(中等CO2排放情景)和RCP8.5(最高CO2排放情景)3种排放情景作为未来气候数据。未来时段包括2030s(2021—2040年)、2050s (2041—2060年)、2070s(2061—2080年)和2080s(2071— 2090年), 数据在国际热带农业中心(International Centre for Tropical Agriculture, CIAT)网站下载[26]。
1.4 中华猕猴桃生境模型构建与结果评价
模型的使用: 输入中华猕猴桃分布点数据和环境因子数据, 随机选取75%的中华猕猴桃分布点作为训练集(training data)建立预测模型, 剩余25%的中华猕猴桃分布点作为测试集(test data)验证模型, 选择刀切法(jackknife test)测定各变量权重, 选择创建环境变量响应曲线, 其余参数均选择模型的默认值[27]。
模型输出的数据为ASCⅡ格式, 利用ArcGIS的ArcToolbox的格式转换工具, 将该数据转为Raster格式, 使该结果可在ArcGIS中显示。MaxEnt软件模拟输出的结果值在0~1之间, 值越接近1表示物种越可能存在。本研究采用受试者工作特征曲线(ROC曲线, receiver operating characteristic curve)分析法对模拟结果进行精度评价, 曲线下面积值(area under curve, AUC)反映预测精度, 理论上AUC指标取值范围为0.5~1, AUC值越接近1表示模型预测结果精度越高。具体评价标准见表1[28-29]。
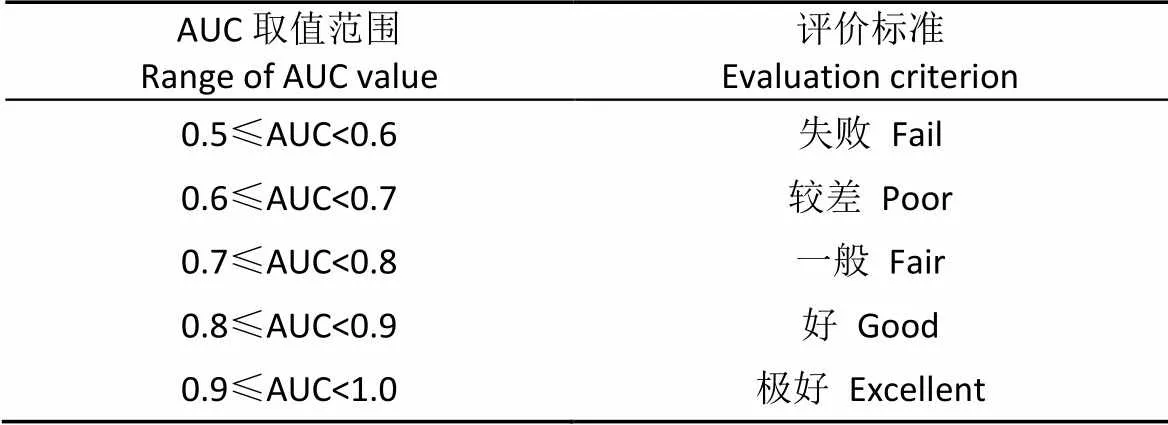
表1 曲线下面积值(AUC值)取值范围及其与MaxEnt模型准确性的关系
1.5 中华猕猴桃适生等级划分
MaxEnt模型输出结果为ASCⅡ格式文件, 首先使用ArcGIS的ArcToolbox的格式转换工具, 将该数据转为Raster格式, 使该结果可在ArcGIS中显示, 而后使用“提取分析”功能得到中华猕猴桃在研究区的存在概率分布图[24,30]。MaxEnt软件模拟输出的结果值为0~1, 值越接近1表示物种越可能存在。参考IPCC报告[31-32]关于评估可能性的划分方法, 结合中华猕猴桃的实际情况, 利用“Reclassify”功能, 划分分布值等级及相应分布范围, 并使用不同颜色表示, 划分标准为: 存在概率<0.05为不适生区; 0.05≤存在概率<0.33为低适生区; 0.33≤存在概率<0.66为中适生区; 存在概率≥0.66为高适生区。
1.6 几何中心及位移计算
参考Yue等[33]的计算方法, 统计了高适生区在未来不同时期的面积变化情况以及其中心点的位移, 具体计算公式如下:
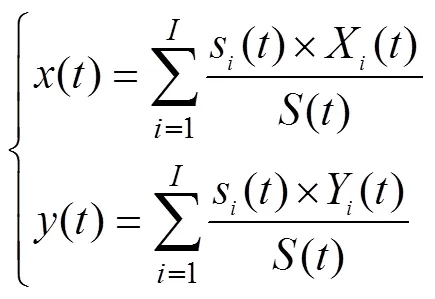
式中:表示不同的时段;为高适生区的单位栅格数目;s()为时段第(0, 1, 2, …,)个单位栅格的面积;()为时段高适生区的总面积;X()和Y()分别为时段第个单位栅格质心的经度和纬度;()、()分别为时段高适生区质心的经度和纬度。
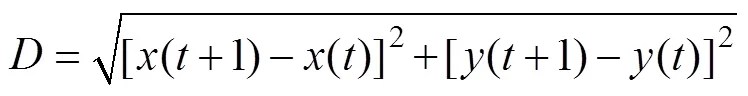
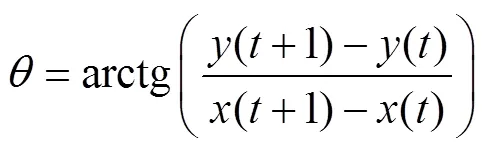
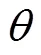
2 结果与分析
2.1 影响中华猕猴桃分布的关键环境变量的筛选及模拟评价
2.1.1 潜在环境变量的筛选
本研究选择的初始环境变量包括预测物种分布通用的19个生物气候变量和月平均气候数据, 研究表明, 这些变量之间存在不可避免的自相关及多重线性重复等问题, 这些高度相关的变量在模型预测过程中会引入冗余信息, 影响预测结果[34-35]。因此, 应首先对环境变量进行相关分析和有效筛选。本研究中, 参考Worthington等[36]方法, 筛选潜在环境变量, 具体过程为: 首先全部环境变量构建初始模型, 选择MaxEnt软件中的刀切法(Jackknife)检验来测定环境变量对模型预测的贡献大小, 以此剔除对MaxEnt模型预测结果贡献较小的环境变量。其次, 利用SPSS软件对贡献较大的关键限制因子进行Searman相关分析, 若相关系数||≥0.8, 则对比初始模型中二者的贡献率, 剔除贡献率较小的变量, 提高模型模拟的精度。经过上述过程, 共保留了22个环境变量(代码和计算单位见表2), 在此基础上重建中华猕猴桃在中国分布的最大熵模型, 并对模拟结果进行准确性评价。
2.1.2 ROC曲线和AUC值对模型准确性评价
图1是当前时段气候条件下的ROC曲线图, 图中表明基于主导环境变量构建的中华猕猴桃在中国的地理分布模型的AUC值为0.960, 依据表1的AUC值评价标准, 本次构建模型的预测准确性达到“极好”的标准; 图2为未来不同气候情境下MaxEnt模拟结果的ROC曲线图, 由图2可知所有预测结果的AUC值都达到“极好”的标准。上述结果说明模型预测可信度较高, 可用于分析气候变化对中华猕猴桃在中国分布的影响。

表2 影响中华猕猴桃分布的22个环境变量及其代码和计量单位
2.2 中华猕猴桃在中国的潜在分布
由预测结果(图3)和统计分析(表3)可知, 当前气候条件下, 中华猕猴桃在中国的适生区在25°~36°N, 101°~122°E, 由东向西呈不连续分布, 总面积达2.58×106km2, 占中国国土面积的26.92%。其中高适生区主要位于四川、陕西、重庆、湖北、贵州、浙江、湖北、安徽、河南、江苏和甘肃等省份, 面积达1.01×106km2, 占总适生面积的38.94%; 中适生区沿高适生区周围分布, 主要在中国中东部地区, 包括河南、湖北、安徽、江苏和山东等地, 面积为6.79×105km2, 占总适生面积的26.26%; 低适生区包括云南、江西、河北和辽宁等省份, 面积为8.99×105km2, 占总适生面积的34.8%。

图1 当前气候情景中华猕猴桃MaxEnt模型的ROC曲线

图2 未来不同年代3种气候变化情景下中华猕猴桃MaxEnt模型的ROC曲线
本研究选取的未来时段为2030s、2050s、2070s和2080s, 3种气候模式为RCP2.6、RCP4.5和RCP8.5, 由图4和表3可知, 上述情景下中华猕猴桃在研究区的高、中适生区仍主要集中在四川、陕西、重庆、湖北等省区, 只是在不同时段、不同气候情景下分布范围及面积有所不同。
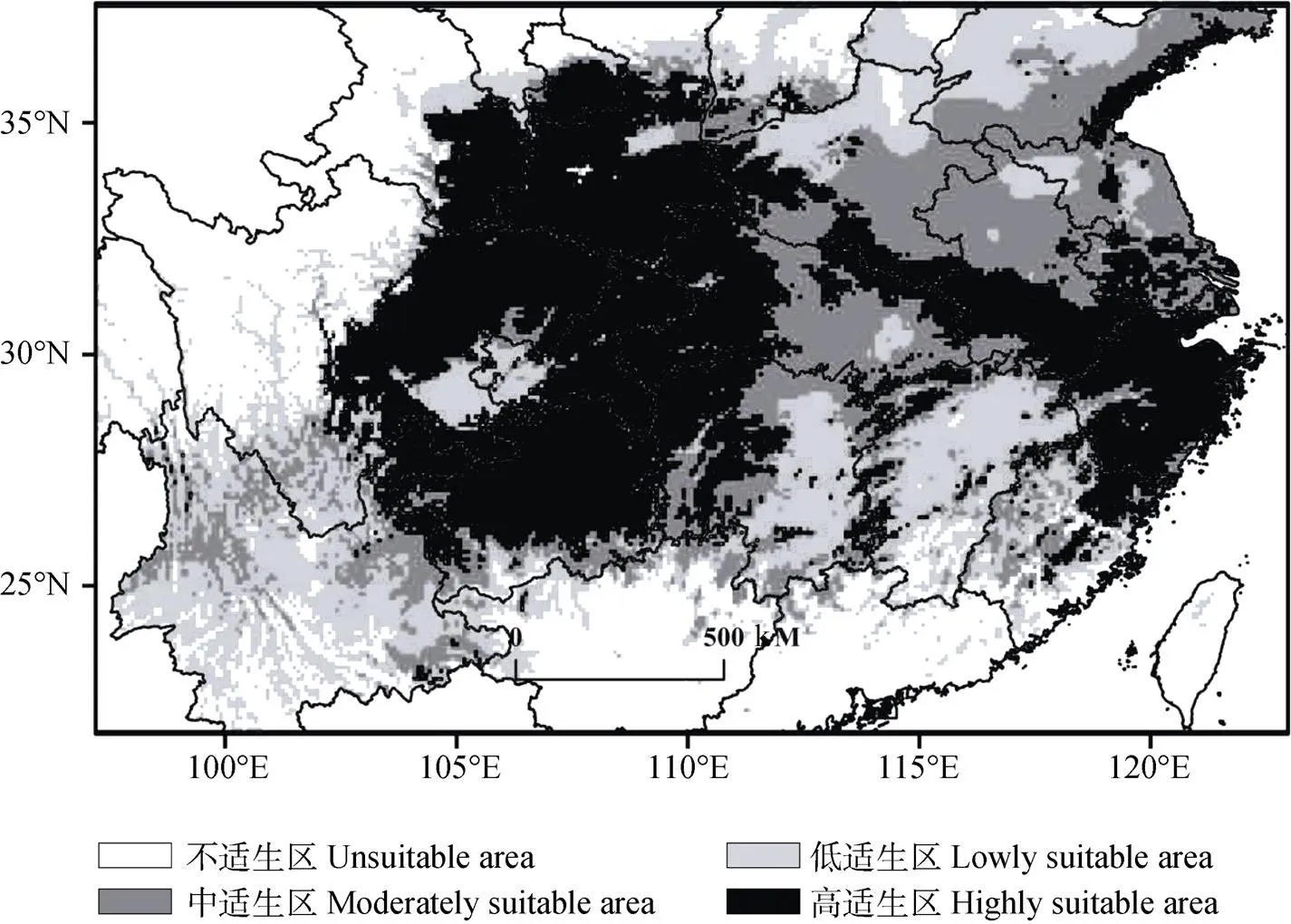
图3 基于MaxEnt模型预测的中华猕猴桃在中国的适生分析图
2.3 中华猕猴桃在中国高适生区面积变化及中心点位移
当前及RCP2.6情景下, 高适生区面积呈波动增加的变化趋势。总体上由当前的1.01×106km2增加至2080s的1.08×106km2(表3)。当前至2080s, 高适生区中心点由重庆云阳(当前)经重庆巫山(2030s)、重庆奉节(2050s和2070s)位移至湖北恩施(2080s)(图5), 总体上向东南方向移动82.39 km(表4)。
当前及RCP4.5情景下, 高适生区面积呈先减少后增加的变化趋势。总体上由当前的1.01×106km2增加至2080s的1.48×106km2(表3)。当前至2080s, 高适生区中心点由重庆云阳(当前和2030s)经湖北恩施(2050s)、重庆奉节(2070s)位移至重庆巫溪(2080s)(图5), 总体上向西北方向移动32.44 km(表4)。
当前及RCP8.5情景下, 高适生区面积呈先增加后减少变化趋势。总体上由当前的1.01×106km2增加至2080s的1.09×106km2(表3)。当前至2080s, 高适生区中心点由重庆云阳(当前)经重庆巫山(2030s)、重庆巫溪(2050s)、四川宣汉(2070s)位移至重庆巫溪(2080s)(图5), 总体上向东北方向移动40.82 km(表4)。
3 讨论与结论
MaxEnt软件基于最大熵原理, 以物种分布变量和环境变量为基础, 利用数学模型统计分析熵最大时物种的分布状态。众多研究表明[37], 在分布数据较少时MaxEnt模型的预测结果比同类预测模型更精确, 因此本研究选取MaxEnt软件来预测并分析中华猕猴桃的适生区及适生等级。目前普遍应用ROC曲线法(即AUC法)对模型精度进行评价[38], AUC的取值范围在0.5~1, 值越接近于1模型精度越高[39]。本研究结果表明, 当前及未来气候模式下中华猕猴桃的适生区预测的AUC值均大于0.9, 预测准确性达“极好”标准, 说明此次模型预测的地理分布与中华猕猴桃实际分布拟合度较高, 可以用于中华猕猴桃在中国地理分布与气候关系的研究。另外, 本研究利用地理信息系统软件ArcGIS对MaxEnt输出的栅格文件进行后期处理, 使目标物种的分布数据与环境变量数据在栅格单元上相对应, 有效降低了系统误差, 进一步提高了数据的准确性。
本研究利用ArcGIS软件统计了未来3种气候变化情景下中华猕猴桃适生区面积变化情况, 结果显示, 至2080s, 高、中适生区面积在RCP2.6、RCP4.5和RCP8.5情景下均有所增加, 只是增加的幅度不同。这说明在不同排放情景下气候变化对中华猕猴桃潜在分布的影响有一定的不确定性。环境变量的选择和作物分布点数据的多少对MaxEnt模型的模拟准确性有较大影响。张杰等[40]利用MaxEnt分析了中等温室气体排放情景下(RCP4.5)中华猕猴桃在中国潜在分布区的变化趋势, 结果表明, 至2050年, 中华猕猴桃适生区面积较当前情景略有下降, 此结论与本文研究结果并不完全吻合, 可能是选择了不同的变量和不同的分布点数据所致。
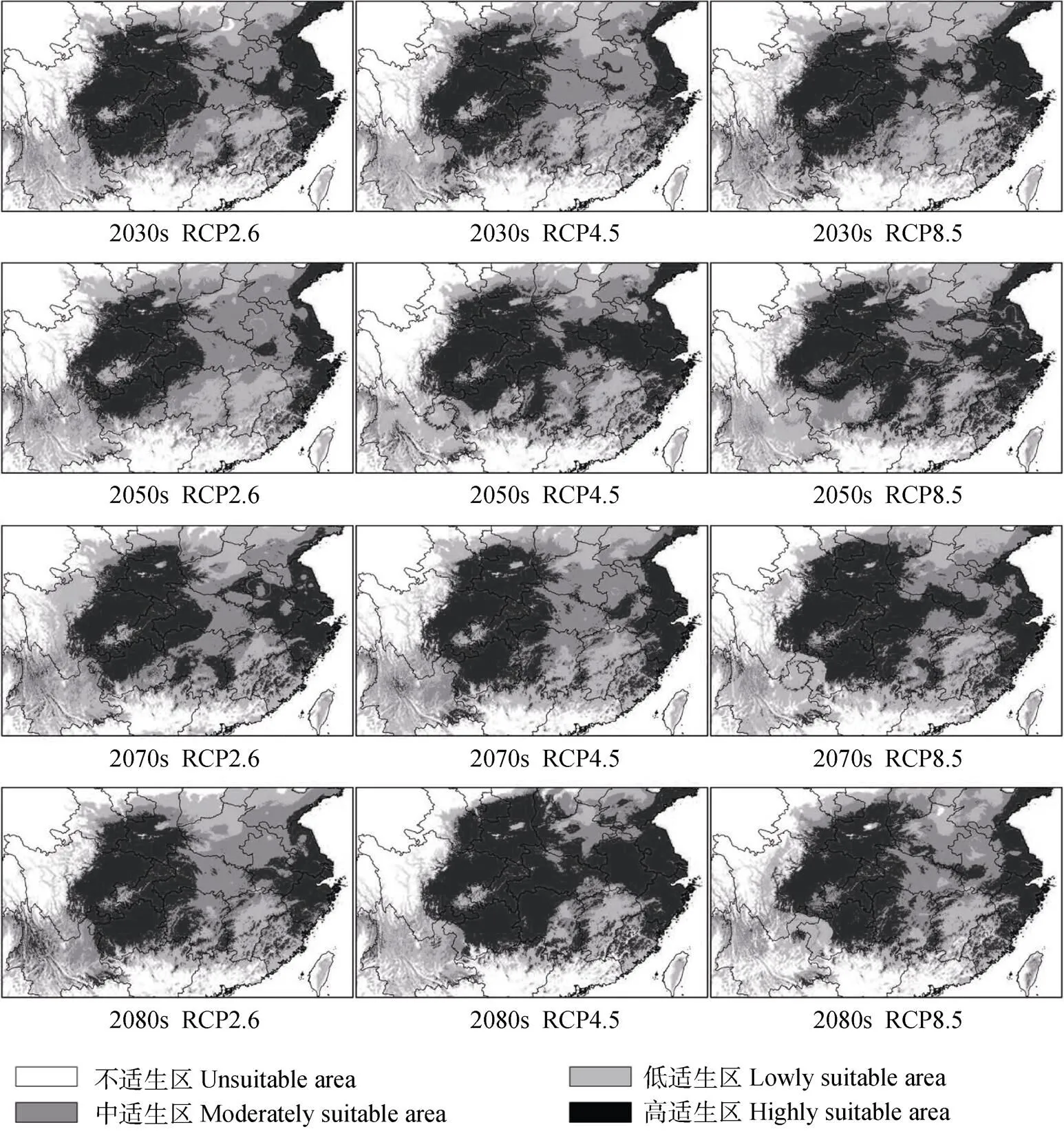
图4 未来不同年代3种气候变化背景下中华猕猴桃潜在适生区的预测结果
表3 中华猕猴桃在当前情景(1950—2000年)及未来气候条件下的适生区面积预测
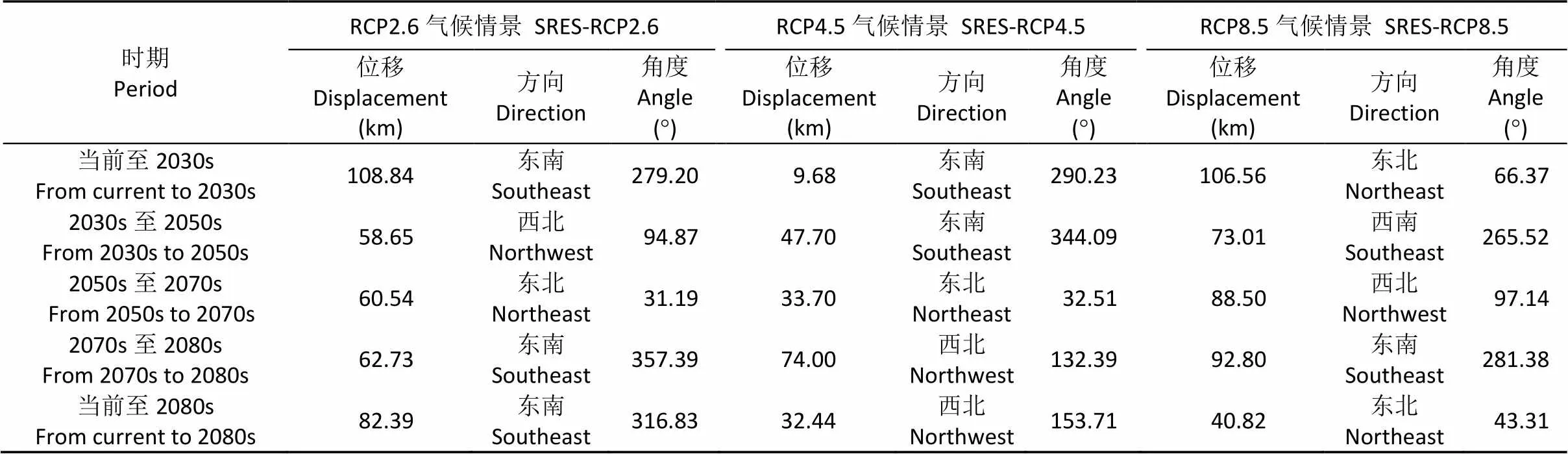
表4 未来3种气候变化情景下中华猕猴桃高适生区中心点位移距离和方向
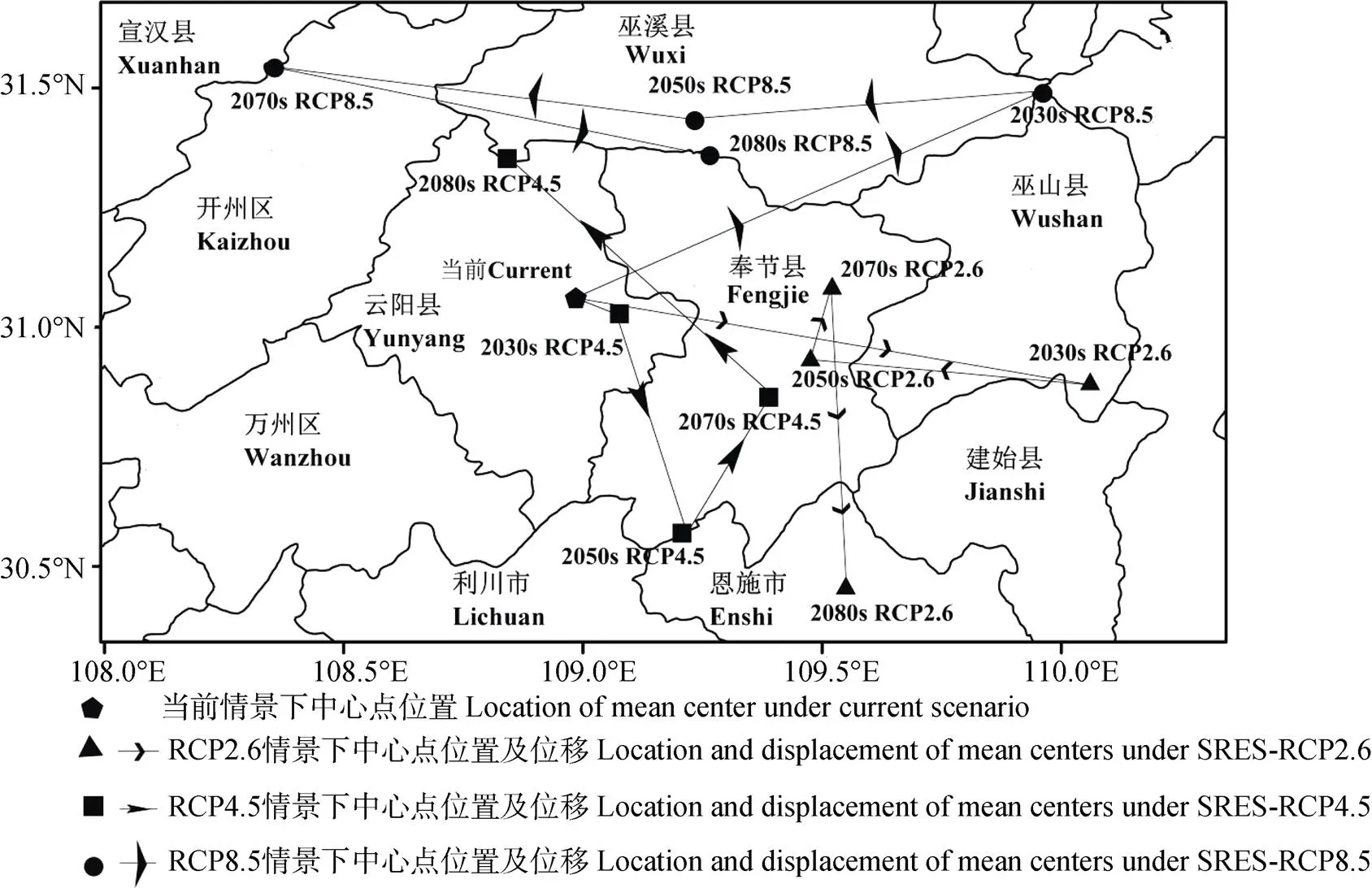
图5 未来不同年代3种气候变化情景下中华猕猴桃高适生区中心点迁移轨迹
气候是决定地球上物种分布的最主要因素, 而物种分布格局的变化则是对气候变化最明确和直接的反应。气候变暖可能实质性地改变陆地生态系统的结构和功能, 使生物栖息地范围与分布区发生变化。本文参考Yue等[33]的计算方法, 以面积作为权重, 计算了不同等级适生区中心点位置, 着重分析了适生区中心点随时间变化的位移趋势, 展示了中华猕猴桃对气候变化的响应过程, 结果显示, 到2080s, 高、中适生区的中心点均将发生不同程度的改变。其中高适生区中心点在3种情景下均向偏北方向移动, 原因可能与研究区的年平均气温与年降水量的增加有关(表5); 从结果中还可看出, 不同概率等级的中心点位没有明显的规律性, 其原因可能在于当前和未来气候变量的不连贯, 当前气候数据的起止时间为1950—2000年, 而未来气候数据的起止时间为2030—2080年, 2000—2030年的数据缺失可能造成中心点移动方向的往复。
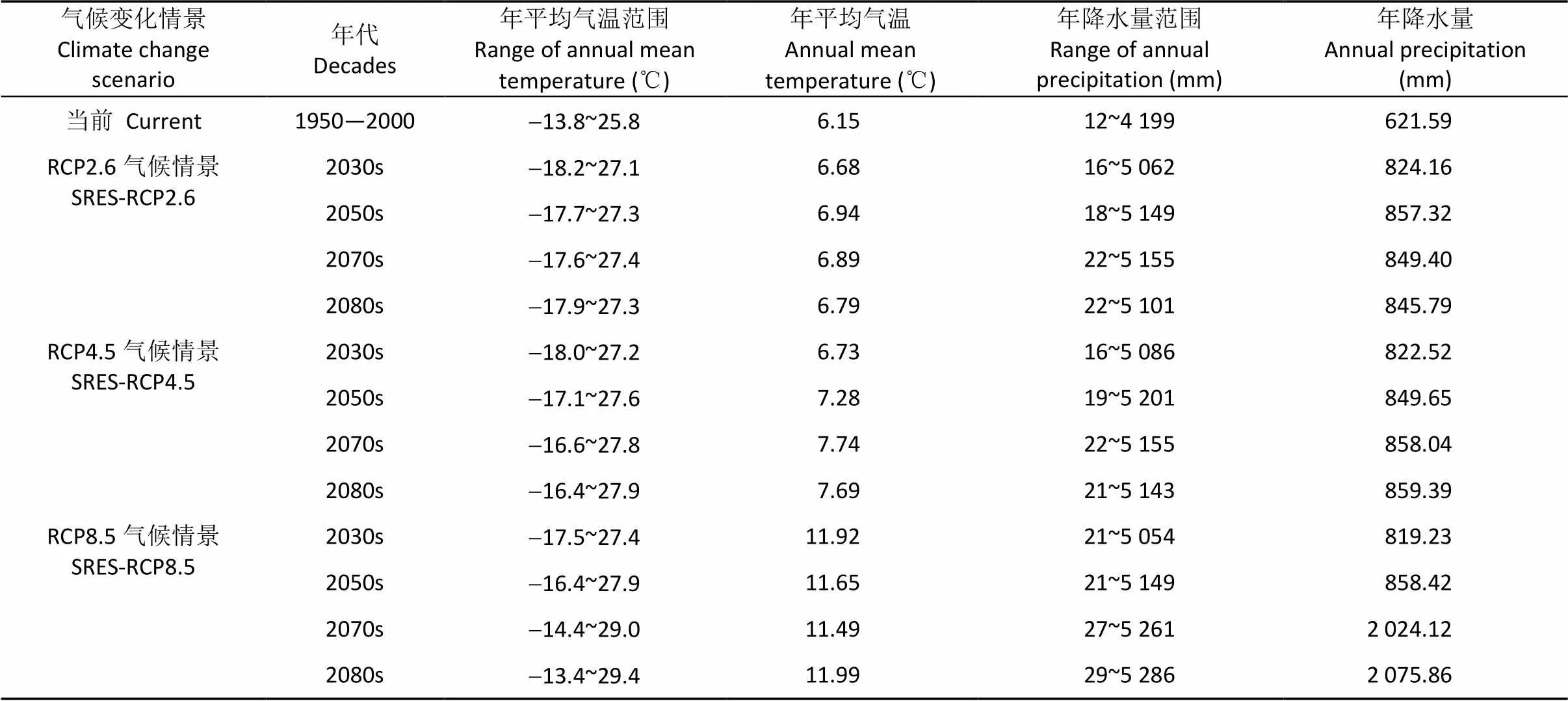
表5 气候变化情景下未来不同年代中国年平均气温与年降水量的变化
生态位模型假设物种的生态位需求是保守的, 样本大小、空间尺度和环境变量等因素会影响此类模型的预测能力和稳定性[41]。本研究中发现以下问题: 1)文中选取的19个环境变量是以温度和雨量为基础, 根据不同需求计算演生而来, 因此这些变量之间存在不可避免的自相关及多重线性重复等问题, 在模型预测过程中会引入冗余信息, 影响预测结果。因此本文首先运用刀切法对环境变量进行相关分析和有效筛选, 计算各因子对物种分布的贡献率大小, 剔除贡献率较小的环境变量, 基于主导环境变量重建模型, 提高了预测结果的准确性。2)本研究预测了未来情景下中华猕猴桃的适生区变化, 使用RCP2.6、RCP4.5和RCP8.5 3种气候模式数据。需要注意的是, 预测是以当前中华猕猴桃的发生点作为分布数据变量, 这就可能会忽略未来实际分布点的影响, 不可避免地造成系统误差。3)影响中华猕猴桃在中国种植分布的环境因子不仅仅包括气候条件、土壤类型、植被类型和地形因子、品种类型、人类活动等非生物因子, 社会经济结构、生产技术水平等因素同样会对其分布产生重要影响[42-43]。基于上述原因, 可以推测, MaxEnt模型预测的生态位比中华猕猴桃所占据的实际生态位要宽。本研究由于数据限制, 仅考虑了温度和降水这两类气候变量对适生分布的影响, 在下一步工作中, 还应注重考量各种因素相互作用的可信表达, 以改善模型的预测效果。
[1] 吴建国, 吕佳佳, 艾丽. 气候变化对生物多样性的影响: 脆弱性和适应[J]. 生态环境学报, 2009, 18(2): 693–703 Wu J G, LYU J J, Ai L. The impacts of climate change on the biodiversity: Vulnerability and adaptation[J]. Ecology and Environmental Sciences, 2009, 18(2): 693–703
[2] 李勇, 杨晓光, 王文峰, 等. 全球气候变暖对中国种植制度可能影响Ⅴ. 气候变暖对中国热带作物种植北界和寒害风险的影响分析[J]. 中国农业科学, 2010, 43(12): 2477–2484 Li Y, Yang X G, Wang W F, et al. The possible effects of global warming on cropping systems in China Ⅴ. The possible effects of climate warming on geographical shift in safe northern limit of tropical crops and the risk analysis of cold damage in China[J]. Scientia Agricultura Sinica, 2010, 43(12): 2477–2484
[3] 邓振镛, 张强, 宁惠芳, 等. 西北地区气候暖干化对作物气候生态适应性的影响[J]. 中国沙漠, 2010, 30(3): 633–639 Deng Z Y, Zhang Q, Ning H F, et al. Influence of climate warming and drying on crop eco-climate adaptability in Northwestern China[J]. Journal of Desert Research, 2010, 30(3): 633–639
[4] 王晓煜, 杨晓光, 孙爽, 等. 气候变化背景下东北三省主要粮食作物产量潜力及资源利用效率比较[J]. 应用生态学报, 2015, 26(10): 3091–3102 Wang X Y, Yang X G, Sun S, et al. Comparison of potential yield and resource utilization efficiency of main food crops in three provinces of Northeast China under climate change[J]. Chinese Journal of Applied Ecology, 2015, 26(10): 3091–3102
[5] Anderson R P, Raza A. The effect of the extent of the study region on GIS models of species geographic distributions and estimates of niche evolution: Preliminary tests with montane rodents (genus) in Venezuela[J]. Journal of Biogeography, 2010, 37(7): 1378–1393
[6] 朱耿平, 刘国卿, 卜文俊, 等. 生态位模型的基本原理及其在生物多样性保护中的应用[J]. 生物多样性, 2013, 21(1): 90–98 Zhu G P, Liu G Q, Bu W J, et al. Ecological niche modeling and its applications in biodiversity conservation[J]. Biodiversity Science, 2013, 21(1): 90–98
[7] 许仲林, 彭焕华, 彭守璋. 物种分布模型的发展及评价方法[J]. 生态学报, 2015, 35(2): 557–567 Xu Z L, Peng H H, Peng S Z. The development and evaluation of species distribution models[J]. Acta Ecologica Sinica, 2015, 35(2): 557–567
[8] 李国庆, 刘长成, 刘玉国, 等. 物种分布模型理论研究进展[J]. 生态学报, 2013, 33(16): 4827–4835 Li G Q, Liu C C, Liu Y G, et al. Advances in theoretical issues of species distribution models[J]. Acta Ecologica Sinica, 2013, 33(16): 4827–4835
[9] 段居琦, 周广胜. 中国水稻潜在分布及其气候特征[J]. 生态学报, 2011, 31(22): 6659–6668 Duan J Q, Zhou G S. Potential distribution of rice in china and its climate characteristics[J]. Acta Ecologica Sinica, 2011, 31(22): 6659–6668
[10] 何奇瑾, 周广胜. 我国春玉米潜在种植分布区的气候适宜性[J]. 生态学报, 2012, 32(12): 3931–3939 He Q J, Zhou G S. Climatic suitability of potential spring maize cultivation distribution in China[J]. Acta Ecologica Sinica, 2012, 32(12): 3931–3939
[11] 孙敬松, 周广胜. 利用最大熵法(MaxEnt)模拟中国冬小麦分布区的年代际动态变化[J]. 中国农业气象, 2012, 33(4): 481–487 Sun J S, Zhou G S. Inter-decadal variability of winter wheat planting zone in China during 1961 to 2010 simulated by maximum entropy (MaxEnt)[J]. Chinese Journal of Agrometeorology, 2012, 33(4): 481–487
[12] 黄宏文, 龚俊杰, 王圣梅, 等. 猕猴桃属()植物的遗传多样性[J]. 生物多样性, 2000, 8(1): 1–12 Huang H W, Gong J J, Wang S M, et al. Genetic diversity in the genus[J]. Chinese Biodiversity, 2000, 8(1): 1–12
[13] 徐小彪, 张秋明. 中国猕猴桃种质资源的研究与利用[J]. 植物学通报, 2003, 20(6): 648–655 Xu X B, Zhang Q M. Researches and utilizations of germplasm resource of kiwifruit in China[J]. Chinese Bulletin of Botany, 2003, 20(6): 648–655
[14] 刘瑶, 朱天辉, 樊芳冰, 等. 四川猕猴桃溃疡病的发生与病原研究[J]. 湖北农业科学, 2013, 52(20): 4937–4942 Liu Y, Zhu T H, Fan F B, et al. Occurrence and pathogen identification of kiwifruit bacterial canker in Sichuan[J]. Hubei Agricultural Sciences, 2013, 52(20): 4937–4942
[15] 涂美艳, 江国良, 陈栋, 等. 四川省猕猴桃产业发展现状及对策[J]. 湖北农业科学, 2012, 51(10): 1945–1949 Tu M Y, Jiang G L, Chen D, et al. Development status and countermeasures of kiwifruit industry in Sichuan Province[J]. Hubei Agricultural Sciences, 2012, 51(10): 1945–1949
[16] 于成, 叶丽君, 刘泽全, 等. 都江堰市海沃特猕猴桃种植的气候适应性区划[J]. 安徽农业科学, 2010, 38(11): 5741–5743 Yu C, Ye L J, Liu Z Q, et al. Climate adaptability division ofcv. Hayward in Dujiangyan[J]. Journal of Anhui Agricultural Sciences, 2010, 38(11): 5741–5743
[17] 贺文丽, 李星敏, 朱琳, 等. 基于GIS的关中猕猴桃气候生态适宜性区划[J]. 中国农学通报, 2011, 27(22): 202–207 He W L, Li X M, Zhu L, et al. Climate ecological applicability regionalization for kiwifruit based on GIS in Guanzhong of Shaanxi Province[J]. Chinese Agricultural Science Bulletin, 2011, 27(22): 202–207
[18] 莫建国, 池再香, 汤苾, 等. 贵州山区红心猕猴桃种植气候区划[J]. 中国农业气象, 2016, 37(1): 36–42 Mo J G, Chi Z X, Tang B, et al. Climate suitability for red cartridge kiwifruit planting in Guizhou mountainous area[J]. Chinese Journal of Agrometeorology, 2016, 37(1): 36–42
[19] 曾永美, 高阳华, 杨世琦. 基于GIS的重庆市万盛区猕猴桃气候区划分析[J]. 重庆师范大学学报: 自然科学版, 2012, 29(2): 89–93 Zeng Y M, Gao Y H, Yang S Q. The kiwifruit climate division analysis of Chongqing Wansheng District that based on geographic information system[J]. Journal of Chongqing Normal University: Natural Science Edition, 2012, 29(2): 89–93
[20] 刘敏, 刘云鹏, 刘宗芳, 等. 夷陵区野生猕猴桃生态气候适应性及其区划[J]. 湖北气象, 2003, (2): 18–20 Liu M, Liu Y P, Liu Z F, et al. Wild Chinese gooseberry ecologic climatization in Yi-Ling County and its region division[J]. Meteorology Journal of Hubei, 2003, (2): 18–20
[21] Ha H, Heumann B W, Liesch M, et al. Modelling potential conservation easement locations using physical and socio-economic factors: A case-study from south-east Michigan[J]. Applied Geography, 2016, 75: 104–115
[22] 郭杰, 刘小平, 张琴, 等. 基于MaxEnt模型的党参全球潜在分布区预测[J]. 应用生态学报, 2017, 28(3): 992–1000 Guo J, Liu X P, Zhang Q, et al. Prediction for the potential distribution area ofat global scale based on MaxEnt model[J]. Chinese Journal of Applied Ecology, 2017, 28(3): 992–1000
[23] 张天蛟, 刘刚. 提高生态位模型时间转移能力的方法研究[J]. 中国农业大学学报, 2017, 22(2): 98–105 Zhang T J, Liu G. Study of methods to improve the temporal transferability of niche model[J]. Journal of China Agricultural University, 2017, 22(2): 98–105
[24] 曹学仁, 陈林, 周益林, 等. 基于MaxEnt的麦瘟病在全球及中国的潜在分布区预测[J]. 植物保护, 2011, 37(3): 80–83 Cao X R, Chen L, Zhou Y L, et al. Potential distribution ofin China and the world, predicted by MaxEnt[J]. Plant Protection, 2011, 37(3): 80–83
[25] 闫冠华, 李巧萍, 邢超. 不同温室气体排放情景下未来中国地面气温变化特征[J]. 南京信息工程大学学报: 自然科学版, 2011, 3(1): 36–46 Yan G H, Li Q P, Xing C. Future surface air temperature changes in China under different greenhouse gas emission scenarios[J]. Journal of Nanjing University of Information Science and Technology: Natural Science Edition, 2011, 3(1): 36–46
[26] 张晓华, 高云, 祁悦, 等. IPCC第五次评估报告第一工作组主要结论对《联合国气候变化框架公约》进程的影响分析[J]. 气候变化研究进展, 2014, 10(1): 14–19 Zhang X H, Gao Y, Qi Y, et al. Implications of the findings from the working group Ⅰ contribution to the IPCC fifth assessment report on the UNFCCC process[J]. Progressus Inquisitiones de Mutatione Climatis, 2014, 10(1): 14–19
[27] 王雷宏, 杨俊仙, 徐小牛. 基于MaxEnt分析金钱松适生的生物气候特征[J]. 林业科学, 2015, 51(1): 127–131 Wang L H, Yang J X, Xu X N. Analysis of suitable bioclimatic characteristics ofby using MaxEnt model[J]. Scientia Silvae Sinicae, 2015, 51(1): 127–131
[28] 王运生, 谢丙炎, 万方浩. ROC曲线分析在评价入侵物种分布模型中的应用[J]. 生物多样性, 2007, 15(4): 365–372 Wang Y S, Xie B Y, Wan F H. Application of ROC curve analysis in evaluating the performance of alien species’ potential distribution models[J]. Biodiversity Science, 2007, 15(4): 365–372
[29] 赵力, 朱耿平, 李敏, 等. 入侵害虫西部喙缘蝽和红肩美姬缘蝽在中国的潜在分布[J]. 天津师范大学学报: 自然科学版, 2015, 35(1): 75–78 Zhao L, Zhu G P, Li M, et al. Potential distribution ofandin China[J]. Journal of Tianjin Normal University: Natural Science Edition, 2015, 35(1): 75–78
[30] 赵文娟, 陈林, 丁克坚, 等. 利用MaxEnt预测玉米霜霉病在中国的适生区[J]. 植物保护, 2009, 35(2): 32–38 Zhao W J, Chen L, Ding K J, et al. Prediction of potential geographic distribution areas of the maize downy mildew in China by using MaxEnt[J]. Plant Protection, 2009, 35(2): 32–38
[31] 韩乐琼, 韩哲, 李双林. 不同代表性浓度路径(RCPs)下21世纪长江中下游强降水预估[J]. 大气科学学报, 2014, 37(5): 529–540 Han L Q, Han Z, Li S L. Projection of heavy rainfall events in the middle and lower reaches of the Yangtze River valley in the 21st century under different representative concentration pathways[J]. Transactions of Atmospheric Sciences, 2014, 37(5): 529–540
[32] 赵泽芳, 卫海燕, 郭彦龙, 等. 人参潜在地理分布以及气候变化对其影响预测[J]. 应用生态学报, 2016, 27(11): 3607–3615 Zhao Z F, Wei H Y, Guo Y L, et al. Potential distribution ofand its predicted responses to climate change[J]. Chinese Journal of Applied Ecology, 2016, 27(11): 3607–3615
[33] Yue T X, Fan Z M, Chen C F, et al. Surface modelling of global terrestrial ecosystems under three climate change scenarios[J]. Ecological Modelling, 2011, 222(14): 2342– 2361
[34] 陈新美, 雷渊才, 张雄清, 等. 样本量对MaxEnt模型预测物种分布精度和稳定性的影响[J]. 林业科学, 2012, 48(1): 53–59 Chen X M, Lei Y C, Zhang X Q, et al. Effects of sample sizes on accuracy and stability of maximum entropy model in predicting species distribution[J]. Scientia Silvae Sinicae, 2012, 48(1): 53–59
[35] 朱耿平, 刘强, 高玉葆. 提高生态位模型转移能力来模拟入侵物种的潜在分布[J]. 生物多样性, 2014, 22(2): 223–230Zhu G P, Liu Q, Gao Y B. Improving ecological niche model transferability to predict the potential distribution of invasive exotic species[J]. Biodiversity Science, 2014, 22(2): 223–230
[36] Worthington T A, Zhang T J, Logue D R, et al. Landscape and flow metrics affecting the distribution of a federally-threatened fish: Improving management, model fit, and model transferability[J]. Ecological Modelling, 2016, 342: 1–18
[37] 蔡静芸, 张明明, 粟海军, 等. 生态位模型在物种生境选择中的应用研究[J]. 经济动物学报, 2014, 18(1): 47–52 Cai J Y, Zhang M M, Su H J, et al. Application of ecological niche models for selection of species habitat[J]. Journal of Economic Animal, 2014, 18(1): 47–52
[38] 韩阳阳, 王焱, 项杨, 等. 基于MaxEnt生态位模型的松材线虫在中国的适生区预测分析[J]. 南京林业大学学报: 自然科学版, 2015, 39(1): 6–10Han Y Y, Wang Y, Xiang Y, et al. Prediction of potential distribution ofin China based on MaxEnt ecological niche model[J]. Journal of Nanjing Forestry University: Natural Sciences Edition, 2015, 39(1): 6–10
[39] 康传志, 周涛, 郭兰萍, 等. 全国栽培太子参生态适宜性区划分析[J]. 生态学报, 2016, 36(10): 2934–2944Kang C Z, Zhou T, Guo L P, et al. Ecological suitability and regionalization of(Miq.) Pax ex Pax et Hoffm. in China[J]. Acta Ecologica Sinica, 2016, 36(10): 2934–2944
[40] 张杰, 敖子强, 吴永明, 等. 中华猕猴桃()在中国的适生性及其潜在地理分布模拟预测[J]. 热带地理, 2017, 37(2): 218–225 Zhang J, Ao Z Q, Wu Y M, et al. Prediction of potential geographic distribution ofin China based on maximum entropy niche model and ArcGIS[J]. Tropical Geography, 2017, 37(2): 218–225
[41] 乔慧捷, 胡军华, 黄继红. 生态位模型的理论基础、发展方向与挑战[J]. 中国科学: 生命科学, 2013, 43(11): 915–927Qiao H J, Hu J H, Huang J H. Theoretical basis, future directions, and challenges for ecological niche models[J]. Scientia Sinica Vitae, 2013, 43(11): 915–927
[42] 杨妙贤, 肖德兴, 梁红, 等. 中华猕猴桃性别分化的细胞形态学观察[J]. 园艺学报, 2011, 38(2): 257–264Yang M X, Xiao D X, Liang H, et al. Cytomorphological observation on sex differentiation of[J]. Acta Horticulturae Sinica, 2011, 38(2): 257–264
[43] 任丹, 漆雁斌, 于伟咏, 等. 农户机械使用程度及其影响因素研究——基于四川省205户猕猴桃种植户的调查[J]. 四川农业大学学报, 2016, 34(4): 528–534Ren D, Qi Y B, Yu W Y, et al. Research on the degree of use and the influencing factors of farmers’ machinery — based on the survey of 205 households of kiwifruit farmers in Sichuan Province[J]. Journal of Sichuan Agricultural University, 2016, 34(4): 528–534
Potential distribution ofin China and its predicted response to climate change*
WANG Rulin1,2, LI Qing1**, HE Shisong3, LIU Yuan3
(1. College of Agronomy, Sichuan Agricultural University, Chengdu 611130, China; 2. Sichuan Provincial Rural Economic Information Center, Chengdu 610072, China; 3. Kiwifruit Institute of Cangxi County, Cangxi 628400, China)
Kiwifruit (spp.), belonged to Actinidiaceae, is a type of perennial deciduous woody liana and an important class of berry fruit. With rich sugar, protein, amino acids, vitamins and especially high vitamin C content, the kiwifruit is known as “the king of the fruit” and has a good market prospect.is a species endemic in China with a fast-expanding planting area due to its unique subtle flavor and high economic value. Optimization of planting scale and distribution of the crop has been the major concern for regional planning. The objective of this study was to test and determine the possibility of using the MaxEnt (the maximum entropy) model to simulate and predict future large-scale distribution ofBased on current environmental factors, three future climate scenarios suggested in the IPCC fifth report and current distribution sites of, we used the MaxEnt model in combination with ArcGIS to predict the potential geographic distribution and trend of change ofin China. The dominant factors were chosen by using the Jackknife test and the Receiver Operating Characteristic (ROC) curve was used to evaluate the simulation. The results showed that high value of area under curve (AUC) denoted good results which significantly differed from random predictions. Based on the evaluation criterion, the accuracies of the predictions ofpotential distribution in the current and future periods were excellent.The predicted result of the MaxEnt model was imported into ArcGIS10.0 for further analysis and showed that under present climatic conditions, the total suitable area was 26.92% of the total land area in China. The potential distribution was highly consistent with the locations of specimen records and field surveys. The highly suitable areas were in Sichuan, Shaanxi, Chongqing, Hubei, Guizhou, Zhejiang, Hunan, Anhui, Henan, Jiangsu and Gansu Provinces. The areas of highly suitable habitat in the main producing provinces were analyzed statistically. The results showed that under the current conditions, the most suitable area forcultivation was 1.01×106km2, accounting for 38.94% of the total suitable areas. The moderately suitable areas were in Henan, Hubei, Anhui and Shandong Provinces, with the area of 6.79×105km2,accounting for 26.26% of the total suitable areas. Comparison of future suitable areas with current suitable areas showed that areas of high suitability increased under scenarios RCP2.6 and RCP4.5, but decreased under scenario RCP8.5. Under scenarios RCP4.5 and RCP8.5, the mean center of highly suitable area ofmoved northward. The result showed that the MaxEnt model was highly reliable in determining not only the range of geographic distribution ofbut also in identifying dominant environmental factors driving the geographic distribution. Whereas climate was a decisive factor in species distribution, change in distribution pattern of species was the most direct effect of climate change. The results provided a critical reference base forplantation pattern and countermeasures to cope with climate change in China.
; MaxEnt model; Environmental variables; Climate change; Suitable area analysis; Representative concentration pathway scenario
Corresponding author, E-mail: liq8633@163.com
Jun. 16, 2017; accepted Jul. 12, 2017
10.13930/j.cnki.cjea.170557
S601.9
A
1671-3990(2018)01-0027-11
通信作者:李庆, 主要研究方向为病虫害生物防治。E-mail: liq8633@163.com 王茹琳, 主要研究方向为气候变化与植物、病虫害关系。E-mail: wrl_1986_1@163.com
2017-06-16
2017-07-12
*This study was supported by the Sichuan Fruit Innovation Team Project of Modern Agricultural Industry Technology System of China (2013-2018) and the Project of Meteorological Service for Agriculture, Rural areas and Farmers in Sichuan.
*国家现代农业产业体系四川水果创新团队项目(2013-2018)和四川省三农气象服务专项资助
王茹琳, 李庆, 何仕松, 刘原. 中华猕猴桃在中国潜在分布及其对气候变化响应的研究[J]. 中国生态农业学报, 2018, 26(1): 27-37
WANG R L, LI Q, HE S S, LIU Y. Potential distribution ofin China and its predicted responses to climate change[J]. Chinese Journal of Eco-Agriculture, 2018, 26(1): 27-37
猜你喜欢
西部林业科学(2022年6期)2023-01-03
中国中医药信息杂志(2022年7期)2022-07-17
中国野生植物资源(2022年3期)2022-04-19
快乐语文(2021年36期)2022-01-18
今日农业(2021年11期)2021-11-27
动漫星空(兴趣百科)(2020年12期)2020-12-12
电脑爱好者(2020年10期)2020-07-28
植物资源与环境学报(2020年2期)2020-04-20
电脑爱好者(2019年16期)2019-10-30
作文小学中年级(2019年9期)2019-10-14
