
2,3-丁二醇诱导下匍匐翦股颖叶片细胞结构变化及抗病相关性分析
2017-12-22 01:52刘兴菊马源马晖玲张咏梅余倩倩
草业学报 2017年12期
刘兴菊,马源,马晖玲*,张咏梅,余倩倩
(1.甘肃农业大学草业学院,甘肃 兰州 730070;2.草业生态系统教育部重点实验室,甘肃 兰州 730070;3.中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070;4.甘肃农业大学研究测试中心, 甘肃 兰州 730070;5.华南农业大学园艺学院,广东 广州 510642)
2,3-丁二醇诱导下匍匐翦股颖叶片细胞结构变化及抗病相关性分析
刘兴菊1,2,3,马源1,2,3,马晖玲1,2,3*,张咏梅4,余倩倩5
(1.甘肃农业大学草业学院,甘肃 兰州 730070;2.草业生态系统教育部重点实验室,甘肃 兰州 730070;3.中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070;4.甘肃农业大学研究测试中心, 甘肃 兰州 730070;5.华南农业大学园艺学院,广东 广州 510642)
由立枯丝核菌(Rhizoctoniasolani)引起的褐斑病是匍匐翦股颖草坪最常见的病害之一,2,3-BD(2,3-butanediol)可诱导植物产生抗病反应,提高植物的抗病性。本试验采用250 μmol/L 2,3-BD诱导匍匐翦股颖对褐斑病的抗性,以石蜡切片方法通过显微观察,测定接种立枯丝核菌后匍匐翦股颖叶片细胞显微结构的变化。结果表明,接种病原菌后匍匐翦股颖病情指数第15天达到70.45%,2,3-BD诱导后病情指数显著降低至21.63%。与未诱导接菌处理相比,诱导后匍匐翦股颖叶片细胞结构排列紧密,气腔变小,有较多乳突形成,上、下表皮细胞显著增大,第7天差异最显著,分别是其1.19和1.26倍;叶肉细胞大小于第15天差异最为显著,是其1.30倍;叶绿体数量明显增加,第1、7、15天分别是未诱导接菌处理的1.25、1.20、1.24倍;维管束和导管大小在各时间段均变化不明显,但木质化程度较高,皮层组织加厚。综上所述,250 μmol/L 的2,3-BD能诱导匍匐翦股颖对褐斑病的抗性,并使细胞结构发生变化,其中叶片上、下表皮细胞、叶肉细胞大小和叶绿体数目与抗病性呈正相关性。
2,3-BD;匍匐翦股颖;石蜡切片;细胞结构
匍匐翦股颖(Agrostisstolonifera)是禾本科多年生草本植物,翦股颖属,具匍匐茎,重要的冷季型草坪草之一[1]。一般在温暖、湿润、土壤肥沃的条件下生长良好,匍匐翦股颖的抗寒性强,绿期长,质地纤细柔软,喜光,也具一定耐阴性,能在行道树萌环境中良好生长。由于匍匐茎蔓延能力强,成坪速度快,耐频繁低修剪,适用于保龄球场、高尔夫球场、运动场等高质量高强度草坪建植,也被广泛用于常规城市、庭院、公园以及高速公路等绿化建设[2]。
由于匍匐翦股颖根系入土较浅,喜水,草坪质地密,抗病性较差,易受多种病原菌侵染。在盛夏高温潮湿季节褐斑病(Rhizoctoniasolani)、秆锈病(Pucciniagraminisf. sp.tritici)、钱斑病(Lanziamoellerodiscus)等发病率较高,这些病害通常引起草坪的质量、观赏以及绿化等功能显著退化,给草坪的生产和经营带来巨大损失。褐斑病是匍匐翦股颖最严重的病害之一。该病害主要发生在植株根部和颈部,一般病原菌先从植株根部开始侵染,然后是匍匐茎,最后是叶片。褐斑病是一种土传真菌病害,主要由病原菌立枯丝核菌(Rhizoctoniasolani)引起[3],受侵染植株叶片初期呈水浸状并颜色变暗,后期干枯萎缩,褐斑病在修剪较低的草坪上感病草呈水渍状、颜色变暗,病斑最初一般为紫绿色,感病后期颜色变成浅褐色。病斑的形状略呈圆形或不规则形。在空气湿度较大时,植株叶片感病部的外缘会有“烟圈”出现,这是由病菌的菌丝形成的,在已枯萎和新近感病的叶片上出现,当叶片干枯时“烟圈”会消失,这也是褐斑病鉴定过程中一个重要特征[4-5]。修剪过的草顶端往往是病原菌入侵的主要部位。匍匐翦股颖褐斑病的爆发会引起匍匐翦股颖草坪的大面积死亡,成为匍匐翦股颖草坪质量管理的难题之一。
草坪病害防治中,多采用一些化学杀菌剂来防除病害[6-7],如多菌灵、扑海因、敌菌灵等被大量使用。化学杀菌剂的广泛使用已对环境造成巨大污染,同时使病原菌产生一定抗药性,防除病害效果不太理想。近年来诱导植物抗病性被大量研究,成为一种新的病害防治措施,诱导植物抗病性是通过各种化学、物理等方法诱导激发植物体自身对病原微生物的抗病潜力。诱导植物抗病性一般采用一些无毒或微毒的生物和非生物因子作为诱导因子,对人畜安全无害且抗病效果良好,有效降低化学杀菌剂的使用,可减轻对环境造成的污染,是今后实现作物绿色、安全可持续发展道路的理想选择[8]。Suzuki等[9]在匍匐翦股颖抗病诱导试验中发现病菌荧光假单胞杆菌(Pseudomonasfluorescens)株系HP72可诱发匍匐翦股颖对立枯丝核菌的抗病性。Ryu等[10]指出假单胞菌植物根际促生菌分泌的有机物(2R,3R)-butanediol及其同分异构体2,3-butanediol在诱导植物抗病性过程中有重要作用。其作用方式是以诱导系统抗性(induced systemic resistance,ISR)的抗病机制使植株产生抗病性[11]。马祥等[12-13]采用2R,3R-butanediol以ISR机制诱导处理匍匐翦股颖,能显著降低植株的病情指数,并且提高抗病相关酶活性,其抗病效果显著。
本研究利用2,3-BD诱导处理匍匐翦股颖后,接种立枯丝核菌,采用石蜡切片观察匍匐翦股颖叶片显微结构,体细胞及叶绿体数目等是否发生变化,旨在为丁二醇诱导匍匐翦股颖对褐斑病的抗病性,以便采用诱导方法防治匍匐翦股颖褐斑病的发生,减少杀菌剂对环境的污染。
1 材料与方法
1.1 试验材料
供试材料:匍匐翦股颖品种Penn-A4(A.stolonifera,Penn-A4)种子,由北京克劳沃集团提供(原产地美国);供试药剂:诱导剂2,3-BD购自Sigma;病原菌:匍匐翦股颖褐斑病病原菌为立枯丝核菌,购自中国科学院菌种保存中心。
1.2 试验设计
试验设3个处理:2,3-BD诱导并接种(2,3-butanediol treatment and with pathogenic,BD+T);清水处理并接种(water treatment and with pathogenic,W+T);清水处理不接种为对照(CK),每个处理设5个重复。
供试苗培育:于2016年5月在甘肃农业大学草业学院组培室培育。供试种子依次用无菌水浸泡5 h,70%乙醇溶液浸泡1 min,10%次氯酸钠浸泡15 min,无菌水冲洗6~7次,晾干,种植于沙土(土∶沙=2∶1)的花盆(12 cm×12 cm)中,每盆0.05 g。
诱导与接种:幼苗长出第3片真叶后,用2,3-BD(250 μmol/L) 进行土壤注射诱导处理。诱导处理7 d后接种病原菌,立枯丝核菌于PDB液体培养基中摇床上(25 ℃,100 r/min)培养4~5 d,培养好的菌种研磨成菌丝,无菌水配置成浓度为OD340=0.8的菌丝悬浮液。采用菌丝悬浮液喷雾法接种,每盆40 mL,接种后黑暗保湿16 h,再继续培育。
1.3 试验项目与方法
病情指数调查:接菌种后第7、10、15天,参考南志标[14]的方法进行病情观察,统计病叶率并计算病情指数。匍匐翦股颖褐斑病严重度的分级标准如下,0级:无症状;1级:叶片上有零星菌丝;2级:10%~40%的叶片上有菌丝;3级:40%~60%的叶片上有菌丝体生长并有坏死迹象;4级:60%~80%的叶片上有丰富的菌丝体和坏死现象;5级:全部植株普遍感病。

切片制作:采用常规石蜡切片技术并加以改良。于接种第1、7、15天分别采取材料第3片真叶供试,取叶片中间5~7 mm,FAA(甲醇∶冰乙酸∶70%乙醇=1∶1∶18)固定液固定24 h,经酒精系列脱水,以95%伊红酒精溶液代替95%酒精溶液,以免材料的丢失,二甲苯逐级透明,54~56 ℃石蜡逐步浸蜡、包埋,Leica全自动切片机切片,切片厚度6 μm,40 ℃展片,37 ℃充分烤片,再进行番红(1 h)-固绿(10 s)二重染色法染色。Olympus BX61正置万能显微镜下观察并拍照,Image-Pro Plus 5.0软件对上、下表皮细胞、叶肉细胞、大维管束、导管细胞进行测量,并对叶绿体数目进行统计。
1.4 数据分析
采用SPSS 19.0软件进行统计分析,采用单因素ANOVA进行分析处理,Duncan’s新复极差法进行显著性方差分析, Excel 2010软件进行图表制作。
2 结果与分析
2.1 匍匐翦股颖病情指数分析
由图1所示,2,3-BD处理可有效降低匍匐翦股颖褐斑病的病情指数。随接种时间延长,两个接种处理的病情指数均呈上升趋势。接种后第7天各处理发病不显著,接种后第10天, W+T叶片可观察到少量菌丝的出现,病情指数为5.75%,显著高于BD+T,是其5.64倍。接种后第15天,未诱导接种处理中出现较多菌丝和病斑,病情指数明显升高到70.45%,是2,3-BD诱导处理的3.26倍,且差异显著(P<0.05)。
2.2 匍匐翦股颖叶片显微结构变化

图1 2,3-BD处理对匍匐翦股颖褐斑病病情指数的影响Fig.1 Effects of 2,3-BD on disease index of creeping bentgrass inoculated with R. solani CK:对照清水处理 Water treatment; W+T:清水处理并接种Water treatment and with pathogenic; BD+T:2,3-BD诱导并接种 2,3-butanediol treatment and with pathogenic; 不同小写字母表示差异显著(P<0.05) The different small letters mean the significant differences at P<0.05; 下同 The same below.
匍匐翦股颖叶片主要由三部分组成,分别是表皮、叶肉和叶脉(图2),结构比较简单。其上、下表皮各由一层细胞组成,细胞较小,并且细胞的排列紧密程度不相同,上表皮中有泡状细胞分布,水滴状,不含有叶绿体。叶肉组织比较简单,只由2~3层细胞及围绕着维管束排列的薄壁细胞组成,含有叶绿体。叶脉由一个大维管束和若干小维管束共同组成,是植物叶片的骨架部分,起支撑作用,穿插分布于叶肉细胞中,维管束鞘细胞有1~2层,但并没有与叶肉细胞组成C4植物特有的花环结构。叶片解剖结构显示匍匐翦股颖属C3植物。
2.2.1叶片上表皮细胞大小变化 如图3所示,接菌后随时间变化,匍匐翦股颖叶片上表皮细胞大小没有明显变化趋势。W+T上表皮细胞在第7天与CK相比差异不显著,第15天时减小为CK的95%,且差异性显著。BD+T上表皮细胞显著增大,与CK相比在第1、7天差异显著,分别是其1.14和1.21倍,第15天差异不显著,与W+T相比第1、7、15天均有显著差异,分别是W+T的1.19、1.19和1.08倍(P<0.05)。

图2 匍匐翦股颖叶片解剖结构Fig.2 Anatomical structure of leaf in A. stolonifera 1:叶尖Leaf apex; 2:叶片中部Middle part of leaf; 3:叶脉Leaf vein; co:皮层Cortex; ue:上表皮Upper-epidermis; de:下表皮Down-epidermis; m:叶肉细胞Mesophyll cell; bc:泡状细胞Bulliform cell; vb:维管束Vascular bundle; xy:木质部Xylem; ph:韧皮部Phloem.
2.2.2叶片下表皮细胞大小变化 接种病原菌后随时间变化,匍匐翦股颖叶片下表皮细胞大小无明显变化趋势(图4)。W+T处理与CK相比,各时间段均无显著性差异。2,3-BD对匍匐翦股颖叶片下表皮的影响显著,在第1、7、15天BD+T处理叶片细胞均明显大于CK和W+T处理,差异性显著,分别是其CK的1.14、1.24、1.23倍,W+T的1.18、1.26、1.23倍(P<0.05)。
2.2.3叶片叶肉细胞大小变化 接菌后随时间延长(图5),CK匍匐翦股颖叶片叶肉细胞大小逐渐增大,W+T处理叶肉细胞在第7天时显著增大,第15天迅速减小,均与CK有显著差异,分别是CK的1.15倍和80%。BD+T处理叶肉细胞大小在各时间段无明显变化趋势,第1、7天显著大于CK,分别是CK的1.16和1.12倍,第1、15天显著大于W+T处理,分别是W+T的1.16和1.30倍(P<0.05)。
2.2.4叶片大维管束大小变化 接菌后随时间延长,各处理匐匍翦股颖叶片维管束大小在各时间段变化均不显著(图6)。W+T处理在第15天时减小,是CK的90%。2,3-BD诱导对维管束大小的影响不明显,BD+T处理各时间段与CK均无显著差异,第15天显著大于W+T处理,是其1.10倍(P<0.05)。
2.2.5叶片大维管束导管大小变化 如图7所示,接种病原菌后随时间延长,匍匐翦股颖叶片大维管束导管大小有缓慢减小趋势。W+T处理,在接种病原菌后第15天明显减小,是CK的86%且差异性显著。各时间段BD+T处理导管大小均无显著变化,与CK均无显著差异,第15天显著大于W+T处理,是其1.22倍(P<0.05)。
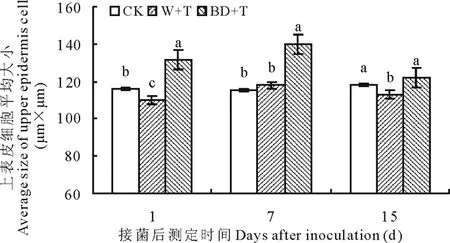
图3 匍匐翦股颖叶片上表皮细胞变化Fig.3 The changes in upper epidermis cell of leaf in A. stolonifera

图4 匍匐翦股颖叶片下表皮细胞变化Fig.4 The changes in down epidermis cell of leaf in A. stolonifera

图5 匍匐翦股颖叶片叶肉细胞变化Fig.5 The changes in mesophyll cell of leaf in A. stolonifera
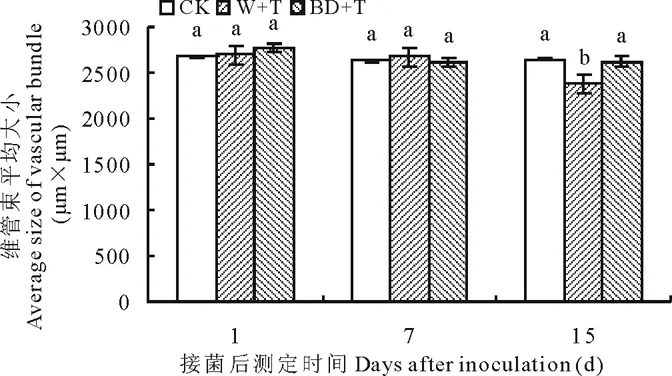
图6 匍匐翦股颖叶片大维管束变化Fig.6 The changes in main vascular bundle of leaf in A. stolonifera

图7 匍匐翦股颖叶片大维管束导管变化Fig.7 The changes in vessels of main vascular bundle in leaf to A. stolonifera
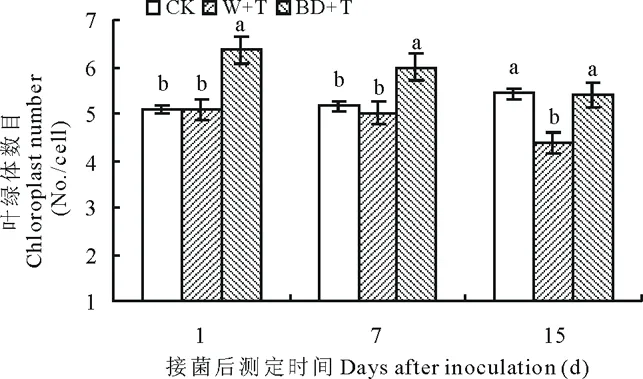
图8 匍匐翦股颖叶片叶绿体变化Fig.8 The changes in chloroplast of leaf in A. stolonifera
2.2.6叶片叶绿体数目变化 如图8所示,接菌后随时间延长,CK匍匐翦股颖叶片叶绿体数目缓慢增加,BD+T和W+T处理呈明显下降趋势。W+T处理第1、7天与CK无显著差异,第15天下降显著,是CK的80%。2,3-BD对匍匐翦股颖叶绿体数目影响较为显著,BD+T处理接菌后第1、7天均显著多于CK,分别是其1.25和1.16倍,第15天时减少,与CK相比差异性不显著;第1、7、15天叶绿体数目均显著多于W+T,分别是其1.25、1.20、1.24倍(P<0.05)。

图9 接种病原菌后匍匐翦股颖叶片解剖结构变化Fig.9 The changes in anatomical structure of leaf in A. stolonifera after inoculation with pathogenic A:接种1 d叶脉Main vein at 1 day after inoculation; B:接种7 d叶脉Main vein at 7 day after inoculation; C:接种15 d叶脉Main vein at 15 day after inoculation; 1:对照清水处理(CK) Water treatment; 2:2,3-BD处理+病原菌2,3-BD treatment+pathogenic; 3:清水处理+病原菌Water treatment+pathogenic; co:皮层Cortex; ue:上表皮Upper-epidermis; de:下表皮Down-epidermis; m:叶肉细胞Mesophyll cell; bc:泡状细胞Bulliform sell; vb:维管束Vascular bundle; xy:木质部Xylem; ph:韧皮部Phloem.
2.3 叶片解剖结构的变化
接种病原菌后随时间的变化,匍匐翦股颖叶片其他显微结构也发生相应的变化(图9)。接种病原菌第1天,CK和W+T处理相比变化不明显,BD+T处理比CK和W+T处理的细胞排列更紧密;叶绿体数量较多且清晰可数,紧靠细胞边缘排列,气腔明显变小,且有较多乳突形成。接种病原菌后第7天,CK、BD+T和W+T处理的叶绿体都未发生较大变化;W+T处理与CK相比气腔变小,BD+T处理气腔也明显较W+T处理与CK小,同时,BD+T和W+T两个处理的皮层组织相比CK有明显加厚现象。接种病原菌后第15天,匍匐翦股颖叶片结构发生较大变化。W+T处理叶绿体变得模糊不清,且数量减少明显,BD+T处理叶绿体仍然清晰可数、排列有序,数量变化不太明显,与CK和W+T处理相比,维管束增大且木质化程度明显升高,皮层组织也有明显增厚现象。
3 讨论与结论
植物抗病性诱导的机理包括分子结构、组织细胞和生理功能等的一系列复杂变化。病原物侵入寄主后,植物的细胞壁、内皮层、厚壁组织、维管束组织、表皮细胞及许多内含物均能抑制病菌的定殖和扩展。已有大量研究表明,诱导剂能使植株细胞结构发生变化来提高植株的抗病作用。2,3-BD在诱导植物抗病方面,近几年研究中也取得了有效成果[10-13]。本试验结果表明,250 μmol/L的2,3-BD可显著降低匍匐翦股颖褐斑病的病情指数,降低病原菌对植株的侵染和伤害,显著提高匍匐翦股颖幼苗期对褐斑病的抗病能力。
病原菌入侵阶段,植物叶片厚度、上、下表皮细胞和皮层等均与抗病性相关。李淼[15]指出,叶片越厚,表皮越厚,表皮上覆盖物越多,则植株对溃疡病(Pseudomonassyringaepv.actinidiae)的抗病效果就越好,上、下表皮细胞与抗病性也有一定关系。司龙亭等[16]试验指出,植物叶片厚度、下表皮厚度、栅栏组织厚度以及叶片结构的紧密程度等均与植物的抗白粉病(Erysiphegraminisf.sp.tritici)能力呈正相关性,其品种抗病品系各指标明显高于感病品系。采用BTH诱导甜瓜对抗抗病性的研究[17-18]中发现,苯并噻二唑(benzothiadiazole,BTH)诱导能使果皮的细胞间隙变小,细胞排列更为紧密且细胞壁加厚以有效抵抗病原菌的入侵。王丽芳[19]同样认为叶片厚度和上、下表皮厚度以及细胞排列紧密度与植物的抗病性为正相关性。蔚丽珍[20]试验指出,病原微生物侵染植株后,叶片叶肉细胞变大,细胞数目减少,叶绿体数量也减少。乳突的形成能使表皮细胞面积增大,排列更为紧密,能有效地阻止病原菌的入侵[21]。本试验结果表明,2,3-BD在匍匐翦股颖结构抗性方面有一定的影响,经2,3-BD诱导处理的匍匐翦股颖叶片细胞排列紧密,气腔变小,上、下表皮细胞有所增大且有较多的乳突形成,接种病原菌后,叶肉细胞也有所增大,随时间变化,叶肉细胞大小的变化趋势较未经2,3-BD诱导的稳定。与上述试验结果一致。匍匐翦股颖叶片结构简单,只有单层表皮细胞,上、下表皮细胞及叶肉细胞的增大都能使叶片的厚度增加,细胞内含物的量也会有所增加,这能使植物在结构抗性方面得到提高,起到抵抗病原菌的作用。
一般而言,维管束越大、数目越多,对植物的养分供应就越好。刘进等[22]试验指出,植物维管束较短或有一定的特殊结构,病害的抗扩展能力就较强,维管束少且木质部导管短这一结构能有效延缓病原菌在植物体的繁殖和进一步扩展,木质化程度高和皮层组织的增厚能有效阻止病原菌的入侵。本试验结果表明,经2,3-BD诱导处理后,匍匐翦股颖叶片大维管束有所增大且木质化程度高,皮层组织加厚。木质部导管的大小变化较小,与未经诱导处理的相比没有差异性,在接种病原菌后,2,3-BD诱导后维管束及导管的大小均没有呈现出明显的变化,而未诱导处理出现缓慢的下降趋势,这说明2,3-BD诱导处理后,能够较长时间地维持匍匐翦股颖叶片维管束的运输功能,使叶片的正常功能得到维持,在抵御病害入侵时起到一定的作用。
叶绿素是植物进行光合作用的基础,光合作用是衡量植物合成功能的重要生理指标。一般植物叶绿素的含量越高,叶片的光合作用越强,植物所积累的有机物质也就越多,能为植株积极抵御病原菌入侵提供的能量就越多,抗性也随之增强。病原物侵染植物后,往往能与叶绿体发生相互作用,导致叶绿体功能减退甚至解体,发病严重的导致叶绿素合成受阻,出现叶片褪绿、黄化或花叶等症状。本试验结果表明,2,3-BD能使匍匐翦股颖叶片叶绿体含量显著增加,接种病原菌后随时间的变化,叶绿体含量逐渐减少,但始终多于未经诱导的处理以及对照组且变化较稳定。试验结果与陈臻等[23]相符,他认为病害能使植物叶绿体含量减少,但抗病品种叶绿素含量下降幅度低于感病品种。王春明等[24]也指出,不同葡萄品种叶片叶绿素含量降幅与品种间的抗病能力呈显著负相关。徐秉良等[25]也发现,叶片内叶绿素含量的高低与植株抗性强弱呈正相关性。这都表明叶绿素含量的变化与植物的抗病性有关。所以,本试验结果说明2,3-BD能使匍匐翦股颖叶片叶绿体含量增加,增强植株对褐斑病侵染的抗性。
References:
[1] Xu Z. The World Gramineae Grass. Beijing: Agricultural Science Publishing House, 1999.
徐柱. 世界禾草属志. 北京: 农业科学出版社, 1999.
[2] Guo W J, Chen P, Liu A. Turf establishment and maintenance of creeping bentgrass in South Subtropical China. Agriculture and Technology, 2004, (2): 111-114.
郭伟经, 陈平, 刘艾. 南亚热带匍匐翦股颖草坪建植与管理技术. 农业与技术, 2004, (2): 111-114.
[3] Piper C V, Coe H S. Rhizoctonia in lawns and pastures. Phytopathology, 1919, 9(1): 89-92.
[4] Smith J D, Jackson N, Woolhouse A R. Fungal Diseases of Amenity Turf Grasses. E & FN Spon Ltd., 1989.
[5] Zhou C Q, Yuan H J. The research on occurrence regularity and control technology toRhizoctoniasolanispeckle diseases of turfgrasses. Agricultural Technology Extension, 2013, (7): 100.
周翠琴, 袁辉杰. 草坪立枯丝核菌褐斑病的发生规律及防治技术研究. 农技推广, 2013, (7): 100.
[6] Chen H B. Effect of several fungicides onPythiumdiseases of turfgrasses. Grassland and Turf, 2009, (2): 17-20.
陈海波. 几种杀菌剂对草坪草腐霉枯萎病的防治试验. 草原与草坪, 2009, (2): 17-20.
[7] Mu H T R, Wang R, Liu R T,etal. Use of carbendazim to controlPythiumdiseases of turfgrasses. Grassland and Turf, 2000, (2): 22-25.
穆合塔尔, 王瑞, 刘荣堂, 等. 多菌灵防治草坪草腐霉枯萎病的研究. 草原与草坪, 2000, (2): 22-25.
[8] Luo L Y. The research progress and application prospect of plant induced disease resistance. Agricultural Technical Services, 2012, 29(2): 175-177.
罗来银. 植物诱导抗病性研究进展及其应用前景. 农技服务, 2012, 29(2): 175-177.
[9] Suzuki S, He Y, Oyaizu H. Indole-3-acetic acid production inPseudomonasfluorescensHP72 and its association with suppression of creeping bentgrass brown patch. Current Microbiology, 2003, 47: 138-143.
[10] Ryu C, Farag M, Hu C,etal. Bacterial volatiles inducesystemic resistance inArabidopsis. Plant Physiology, 2004, 134: 1017-1026.
[11] Cortes-Barco A M, Hsiang T, Goodwin P H. Induced systemic resistance against three foliar diseases ofAgrostisstoloniferaby (2R, 3R)-Butanediol or an isoparaffin mixture. Annals of Applied Biology Plant Pathology, 2010, 157: 179-189.
[12] Ma X, Ma H L, Yao T,etal. Butanediol induced disease resistant against brown spot in creeping bentgrass. Journal of Gansu Agricultural University, 2011, 46(6): 77-80.
马祥, 马晖玲, 姚拓, 等. 新型诱导剂丁二醇对匍匐翦股颖抗病性诱导的研究. 甘肃农业大学学报, 2011, 46(6): 77-80.
[13] Ma X, Ma H L, An H H,etal. Effects on defense-related enzyme activity in creeping bentgrass with inducer butanediol. Grassland and Turf, 2012, 32(3): 37-42, 48.
马祥, 马晖玲, 安惠惠, 等. 诱导剂丁二醇对匍匐翦股颖抗病相关的防卫酶活性的影响. 草原与草坪, 2012, 32(3): 37-42, 48.
[14] Nan Z B. Grass diseases investigation and evaluation//Ren J Z. The Methods of Grassland Science Rerearch. Beijing: China Agriculture Press, 1998: 214-236.
南志标. 牧草病害的调查与评定//任继周.草业科学研究方法. 北京: 中国农业版社, 1998: 214-236.
[15] Li M. Study on the leaf tissue structure of kiwifruit cultivars in relation to bacterial canker disease resistance. Journal of Anhui Agricultural Sciences, 2002, 30(5): 740-742.
李淼. 猕猴桃品种叶片组织结构与抗溃疡病的关系. 安徽农业科学, 2002, 30(5): 740-742.
[16] Si L T, Tian L B. Relationship between the leaf structure of bitter melon and resistance to powdery mildew. Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(10): 2010-2015.
司龙亭, 田丽波. 苦瓜叶片结构与白粉病抗性的关系. 西北植物学报,2013, 33(10): 2010-2015.
[17] Zhang Z K. Preharvest BTH Treatment Induces Resistance in Fruit of Muskmelon. Lanzhou: Gansu Agricultural University, 2006.
张正科. 采用BTH处理对厚皮甜瓜的抗病性诱导. 兰州: 甘肃农业大学, 2006.
[18] Li M. Effect of BTH or Harpin Treatment on Latent Infection in Muskmelons (CucumismelonL.). Lanzhou: Gansu Agricultural University, 2005.
李梅. 采前 BTH 或 Harpin 处理对“银帝”甜瓜潜伏侵染的影响. 兰州: 甘肃农业大学, 2005.
[19] Wang L F. Physiology Mechanism and the Ultrastructure Studies on the Resistance to Sunflower Rust. Hohhot: Inner Mongolia Agricultural University, 2008.
王丽芳. 向日葵抗锈病生理机制及超微结构的研究. 呼和浩特: 内蒙古农业大学, 2008.
[20] Wei L Z. Effects of Antioxidant Enzyme System and Mesophyll Cell Structure on Seeding Leaf of Chinese Cabbage During the Infection of Black Rot. Yangling: North West Agriculture and Forestry University, 2009.
蔚丽珍. 黑腐病侵染对大白菜幼苗叶片抗氧化酶系统和叶肉细胞结构的影响. 杨凌: 西北农林科技大学, 2009.
[21] Bi J X, Yu D D. The research of wheat powdery mildew resistance. Journal of Anhui Agriculture Science, 2008, 36(28): 12320-12321, 12384.
毕建秀, 于冬冬. 小麦白粉病抗性研究. 安徽农业科学, 2008, 36(28): 12320-12321, 12384.
[22] Liu J, Liu J F, Zeng Y,etal. The morphological structure of host plant disease resistance. Journal of Sichuan Forestry Science and Technology, 2001, 22(3): 54-56.
刘进, 刘建锋, 曾义, 等. 寄主植物的形态结构抗病性. 四川林业科技, 2001, 22(3): 54-56.
[23] Chen Z, Xu B L. Relationship between chlorophyll and resistance to leaf blight of turfgrass. Grassland and Turf, 2005, 109(2): 59-61.
陈臻, 徐秉良. 叶绿素含量与草坪草对叶枯病抗病性的关系. 草原与草坪, 2005, 109(2): 59-61.
[24] Wang C M, Du H. Effects of leaf chlorophyll mass fraction on grape cultivar resistance toPlasmoparaviticola. Acta Agriculturae Boreali-Occidentalis Sinica, 2016, 25(3): 458-464.
王春明, 杜蕙. 葡萄叶片叶绿素质量分数与其霜霉病抗性的关系. 西北农业学报, 2016, 25(3): 458-464.
[25] Xu B L, Yan H X. Effects of chlorophyll content and stoma density on pumpkin resistance to Powdery mildew. Plant Protection, 2009, 35(1): 79-81.
徐秉良, 颜惠霞. 南瓜品种对白粉病的抗病性与叶绿素含量和气孔密度的相关性. 植物保护, 2009, 35(1): 79-81.
Thecorrelationbetweencellmicrostructureinducedby2,3-butanediolandfungalresistanceofcreepingbentgrass
LIU Xing-Ju1,2,3, MA Yuan1,2,3, MA Hui-Ling1,2,3*, ZHANG Yong-Mei4, YU Qian-Qian5
1.PrataculturalCollege,GansuAgriculturalUniversity,Lanzhou730070,China; 2.KeyLaboratoryofGrasslandEcosystem,Lanzhou730070,China; 3.SinoU.S.CentersforGrazinglandEcosystemSustainability,Lanzhou730070,China; 4.InstrumentalResearch&AnalysisCenter,GansuAgriculturalUniversity,Lanzhou730070,China; 5.CollegeofHorticulture,SouthChinaAgriculturalUniversity,Guangzhou510642,China
The brown spot caused byRhizoctoniasolaniis one of the most common diseases of creeping bentgrass (Agrostisstolonifera) lawns. Application of 2,3-butanediol (2,3-BD ) can induce plant disease response and improve disease resistance. In the present study, creeping bentgrass was inoculated withR.solaniafter treatment with 2,3-BD (250 μmol/L) and then the cell microstructure of leaves was observed using a paraffin sectioning method. It was found that the disease index of creeping bentgrass without 2,3-BD reached 70.45% after 15 days from inoculation. However, the disease index of creeping bentgrass treated with 2,3-BD was significantly lower, at 21.63% (P<0.05). Compared to leaves of pathogen-infected plants untreated with 2,3-BD, creeping bentgrass leaves with both pathogen inoculation and 2,3-BD treatment had a closely arranged cell structure, smaller cavities, more papillae, and increased upper and lower epidermal cell size. The size increase of upper and lower epidermal cells of 2,3-BD induced plants, compared to uninduced plants, was a factor of 1.19 and 1.26 times, respectively, and was most significant on day 7 after inoculation. The mesophyll cell size increase was the most significant on the 15th day (×1.30). The chloroplast density 1, 7, and 15 days after inoculation was, respectively, 1.25, 1.20, and 1.24 times that of uninduced plants (P<0.05). The size of vascular bundles and ducts did not change significantly at any time, however, the degree of lignification was higher and the cortical tissue became thicker in induced plants. In conclusion, 2, 3-BD can induce resistance of creeping bentgrass to brown spot disease through change in the cell structure. There was a positive correlation between the upper and lower epidermal cell size, the mesophyll cell size, and the number of chloroplasts per cell in creeping bentgrass leaves, and the disease resistance.
2,3-butanediol; creeping bentgrass; paraffin section; the cell structure
10.11686/cyxb2017065http//cyxb.lzu.edu.cn
刘兴菊, 马源, 马晖玲, 张咏梅, 余倩倩. 2,3-丁二醇诱导下匍匐翦股颖叶片细胞结构变化及抗病相关性分析. 草业学报, 2017, 26(12): 170-178.
LIU Xing-Ju, MA Yuan, MA Hui-Ling, ZHANG Yong-Mei, YU Qian-Qian. The correlation between cell microstructure induced by 2,3-butanediol and fungal resistance of creeping bentgrass. Acta Prataculturae Sinica, 2017, 26(12): 170-178.
2017-02-24;改回日期:2017-03-29
国家自然科学
基金项目(31360583)资助。
刘兴菊(1990-),女,甘肃白银人,在读硕士。 E-mail:1437415960@qq.com*通信作者Corresponding author. E-mail: mahl@gsau.edu.cn
猜你喜欢
今日农业(2022年14期)2022-09-15
蔬菜(2022年12期)2022-01-01
星星·散文诗(2021年34期)2021-12-21
河北农业大学学报(2021年5期)2021-11-10
今日农业(2020年13期)2020-12-15
新疆农业科学(2020年9期)2020-10-13
四川蚕业(2020年4期)2020-02-10
河南农业科学(2019年9期)2019-09-24
农村百事通(2019年1期)2019-01-30
烟台果树(2019年1期)2019-01-28
