
海南岛热带山地雨林演替后期幼树的叶片特征
2017-12-22 03:40刘福德安树青曹学章
植物研究 2017年6期
张 明 刘福德 安树青 曹学章
(1.环境保护部南京环境科学研究所,南京 210042; 2.南京大学生命科学学院,南京 210093; 3.天津理工大学环境科学与安全工程学院,天津 300384)
海南岛热带山地雨林演替后期幼树的叶片特征
张 明1,2刘福德3安树青2曹学章1*
(1.环境保护部南京环境科学研究所,南京 210042;2.南京大学生命科学学院,南京 210093;3.天津理工大学环境科学与安全工程学院,天津 300384)
植物的生理生态特征决定了植物在生态系统中的分布模式和物种的丰度。本文在开展样地调查的基础上,应用Li-6400光合测定系统研究了海南岛热带山地雨林原始林3个层次的6个优势种与9个伴生种幼树的光合作用参数,并测定了相应叶片的比叶面积(SLA)和氮、磷含量。15个种的SLA为38.2~143.7 cm2·g-1、单位面积最大光合速率(A-area)为2.77~7.61 μmol·m-2·s-1、单位干重最大光合速率(A-mass)为21.2~83.4 μmol·kg-1·s-1,单位面积暗呼吸速率(Rd-area)为0.02~1.15 μmol·m-2·s-1、单位干重暗呼吸速率(Rd-mass)为0.19~12.61 μmol·kg-1·s-1,光补偿点(LCP)为2.5~32.2 μmol·m-2·s-1,表观量子效率(Ф)为0.006 6~0.042 8 μmol·μmol-1photon,叶片氮(LN)含量为7.98~23.63 mg·g-1,叶片磷(LP)含量为3.98~13.40 mg·g-1。中上层种的幼树具有较高的比叶面积、单位干重最大光合速率和表观量子效率;上层种的暗呼吸速率最低;幼树叶片的氮、磷含量随物种成树所在层次升高而减少,次序为下层种>中层种>上层种,但差异均不显著(P>0.05)。优势种和伴生种的光响应参数与SLA存在相关关系。伴生种的LN与SLA正相关(P<0.01),LP与SLA负相关(P<0.05);优势种的LN、LP与SLA不相关(P>0.05)。
热带山地雨林;演替后期;优势种;伴生种;光合作用;比叶面积;叶片氮;叶片磷
热带森林具有极丰富的植物多样性。迄今为止有超过120种假说来解释为什么有如此多的物种生存在热带森林中,例如空间异质性假说[1]、竞争假说[2]、生产力假说[3]、竞争不平衡假说[4]和能量稳定面积假说[5]等。但是没有一个假说能完全解释热带森林高生物多样性的原因[6]。海南岛热带雨林在演替后期具有很高的生物多样性和复杂的群落结构[6],而林下幼树对群落结构的更新和生物多样性的维持有重要作用。
森林群落的成层结构是不同高度的植物或不同生活型的植物在空间上垂直排列的结果,森林群落的分层与光的利用有关[7]。王刚[8]等对幕阜山森林群落结构开展研究,比较了乔木层、灌木层与草本层的丰富度指数和多样性指数。张明[9]等比较研究了热带山地雨林乔木与灌木幼树的光合生理生态特征。而对于同一群落中,不同层次的乔木幼树的光合生理生态特征的研究尚未见报道。
优势种是对群落结构和群落环境的形成有明显控制作用的植物种群,其在群落中的地位和发展趋势在很大程度上影响着群落的稳定性和物种多样性[10]。对森林群落优势种的研究大多集中在优势种的生态位特征[11]、生物多样性[12]、空间分布[13]等方面。在草本群落中,朱慧[14~15]等比较研究了优势种与伴生种的生理生态特征。而在森林群落中,还未开展过优势种与伴生种的生理生态特征研究。
本文对海南岛热带雨林演替后期原始林不同层次优势种和伴生种的幼树在林下的光合作用光响应、暗呼吸和比叶面积进行研究,从生理生态角度探讨它们对林下光照环境的适应能力和不同层次树种幼树的生存策略。
1 研究地区自然概况
研究地位于海南岛东南部的吊罗山地区。吊罗山地区气候属于东亚季风区,海拔600 m处的雨林年均温度20.8℃,最热月均温度23.9℃,最冷月均温度16.3℃,全年大于10℃的积温7 990℃,年降雨量2 570 mm。成土母岩为花冈岩和闪长岩,土壤为山地黄壤[9]。
样地位于吊罗山原新安林场内,海拔920~980 m,植被类型属于热带山地雨林[16]。样地群落结构复杂,乔木高大,冠层可分为三层,顶层24~28 m,胸径40~60 cm,最大为100 cm;第二层15~23 m,胸径15~30 cm;第三层8~14 m,胸径5~10 cm[16]。林内平均郁闭度为0.97,其组成成分丰富;按株数计算,乔木以樟科个体最多,占10.5%;壳斗科10.0%;桃金娘科5.6%;山茶科4.9%;调查的120个种中主要优势种有阿丁枫(Altingiaobovata)、扫把青冈(Cyclobalanopsisaugustinii)、鸭脚木(Scheffleraoctophylla)等[16]。调查区内竹丛、附生植物普遍存在,板根和层间植物较少,偶有绞杀现象,有大型倒树和枯立木,林下植物贫乏,枯枝落叶层较厚[17]。优势种有阿丁枫(Altingiaobovata)、扫把青冈(Cyclobalanopsisaugustinii)、海南黄叶树(Xanthophyllumhainanense)、岭南青冈(Cyclobalanopsischampionii)、尾叶柯(Lithocarpuscaudatilimbus)、瘤果柯(Lithocarpushandelianus)、台湾枇杷(Eriobotryadeflexa)、鸡毛松(Dacrydiumimbricatus)、鸭脚木(Scheffleraoctophylla)、线枝蒲桃(Syzygiumaraiocladum)、陆均松(Dacrydiumpierrei)、海南大头茶(Gordoniahainanensis)、海岛冬青(Ilexgoshiensis)、卵叶桂(Cinnamomumrigidissimum)、五列木(Pentaphyllaxeuryoides)、海南柿(Diospyroshainanensis)、石碌苦梓(Micheliashiluensis)、多花五月茶(Antidesmamaclurei)、二叶黄杞(Engelhardiaunijuga)和米槠(Castanopsiscarlesii)。
2 研究方法
2.1 物种的选取
根据样地调查与分析,选择5个优势种(dominant species,DS)与10个伴生种(companion species,CS)。5个优势种为:阿丁枫(Altingiaobovata)、尾叶柯(Lithocarpuscaudatilimbus)、绒毛石栎(Quercusgomeziana)、海南黄叶树(Xanthophyllumhainanense)、鸭脚木(Scheffleraoctophylla)、多花五月茶(Antidesmamaclurei)。10个伴生种为:吊罗栎(Quercustiaoloshanica)、木荷(Schimasuperba)、竹叶栎(Quercusbambusifolia)、粗脉樟(Cinnamomumvalidinerve)、鱼骨木(Canthiumdicoccum)、四角蒲桃(Syzygiumtetragonum)、谷木(Memecylonligustrifolium)、子楝树(Decaspermumgracilentum)、山香圆(Turpiniamontana)。其中,阿丁枫、尾叶柯、绒毛石栎、吊罗栎和木荷为上层树种(upper layer species,US);海南黄叶树、鸭脚木、竹叶栎和粗脉樟为中层树种(middle layer species,MS);多花五月茶、鱼骨木、四角蒲桃、谷木、子楝树和山香圆为下层树种(lower layer species,LS)。每个物种3~5个个体。
2.2 光响应曲线的测定
于2005年3~4月,对上述物种进行光合的测定,选择完全舒展并且成熟的未受损叶片,每株3片;用LI-6400便携式光合作用系统(LI-COR,Inc,USA)测定叶片的光响应曲线。使用开放气路,空气流速为0.5 L·min-1,大气温度26±1℃,空气中相对湿度50%~70%,CO2浓度380±10 μmol·mol-1。为了测量的准确和便于比较,测量都在天气晴朗的上午8:00~11:30进行,人工光源使用6400-02B内置式红蓝光源。林下不同幼苗幼树的微环境略有差异,因此在测量前使用人工光源进行5~10 min的光合诱导。光照强度梯度设置为2 500、2 000、1 500、1 000、500、200、100、50、20、0 μmol·m-2·s-1[18]。
2.3 比叶面积的测定
从测定光合的每株幼树上采集10~15片完全展开并且健康的叶片,用光电叶面积仪测定叶片的面积,然后带回实验室用烘箱烘干(105℃下杀青30 min,然后80℃连续烘干48 h),称量烘干重。比叶面积(SLA)定义为叶面积与烘干重的比值(cm-2·g-1)[19~20]。
2.4 叶片氮、磷含量的测定
所采集的叶片在烘箱内在105℃下杀青30 min,然后温度调至80℃连续烘48 h[20],烘干的样品研磨并过2 mm筛,用于叶片氮片(LN)、磷(LP)的测定,测定方法按照实验室常规方法测定[21]。
2.5 数据分析
得到的光强-光合数据根据Prioul & Chartier[22]提供的模型进行非线性回归拟合。具体模型为:
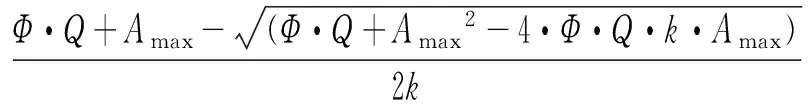
式中:A为净光合速率;Amax为最大净光合速率;Ф表观量子效率;K为曲角;Rday为光下呼吸速率;Q为光照强度。
根据拟合结果,计算出最大光合速率(maximum photosynthesis rate,Amax)、光补偿点(light compensation point,LCP)(表观光合速率为0时的光照强度)和光饱和点(light saturating point,LSP)(表观光合速率开始达到最大值时的光照强度),表观量子效率Ф,曲角K和光下呼吸速率Rday。
试验所得数据采用SPSS13.0软件进行分析。采用最小显著差法(LSD)对植物进行种间多重比较并进行相关分析和T检验。
3 结果与分析
3.1 叶片结构、养分和光合特征
幼树叶片比叶面积(SLA)以中层伴生种鱼骨木最高,中层伴生种粗脉樟最低,仅为鱼骨木的26.6%。幼树叶片氮含量(LN)最高的是中层优势种海南黄叶树,最低的是中层伴生种粗脉樟;磷含量最高的是下层优势种多花五月茶,最低的是上层优势种绒毛石栎。
单位面积最大光合速率(A-area)以上层伴生种木荷最高,中层优势种海南黄叶树次之,下层优势种多花五月茶最低。单位干重最大光合速率(A-mass)仍然以上层伴生种木荷最高,中层伴生种鱼骨木次之;以中层伴生种粗脉樟最低。暗呼吸速率(单位面积Rd-area或单位干重Rd-mass)以中层优势种海南黄叶树最高;以上层伴生种吊罗栎最低。光补偿点(LCP)以上层伴生种木荷和下层伴生种山香圆最高,以中层伴生种竹叶栎和上层优势种阿丁枫最低。表观量子效率(Ф)最高为上层伴生种木荷,最低为下层伴生种子楝树(表1)。
3.2 不同层次树种间光合特征比较
比叶面积(SLA)和表观量子效率(Ф)的次序为中层种>上层种>下层种,但差异不显著(P>0.05);单位面积最大光合速率(A-area)、单位干重暗呼吸速率(Rd-mass)和光补偿点(LCP)的次序为中层种>下层种>上层种,差异也不显著(P>0.05)。中层种的单位重量最大光合速率(A-mass)大于下层种,且差异显著(P<0.05);也大于上层种,但差异不显著(P>0.05)。从下层种到上层种,单位面积暗呼吸速率(Rd-area)、叶片氮含量(LN)和叶片磷含量(LP)逐渐减少,次序为下层种>中层种>上层种,但差异不显著(P>0.05)(表2)。
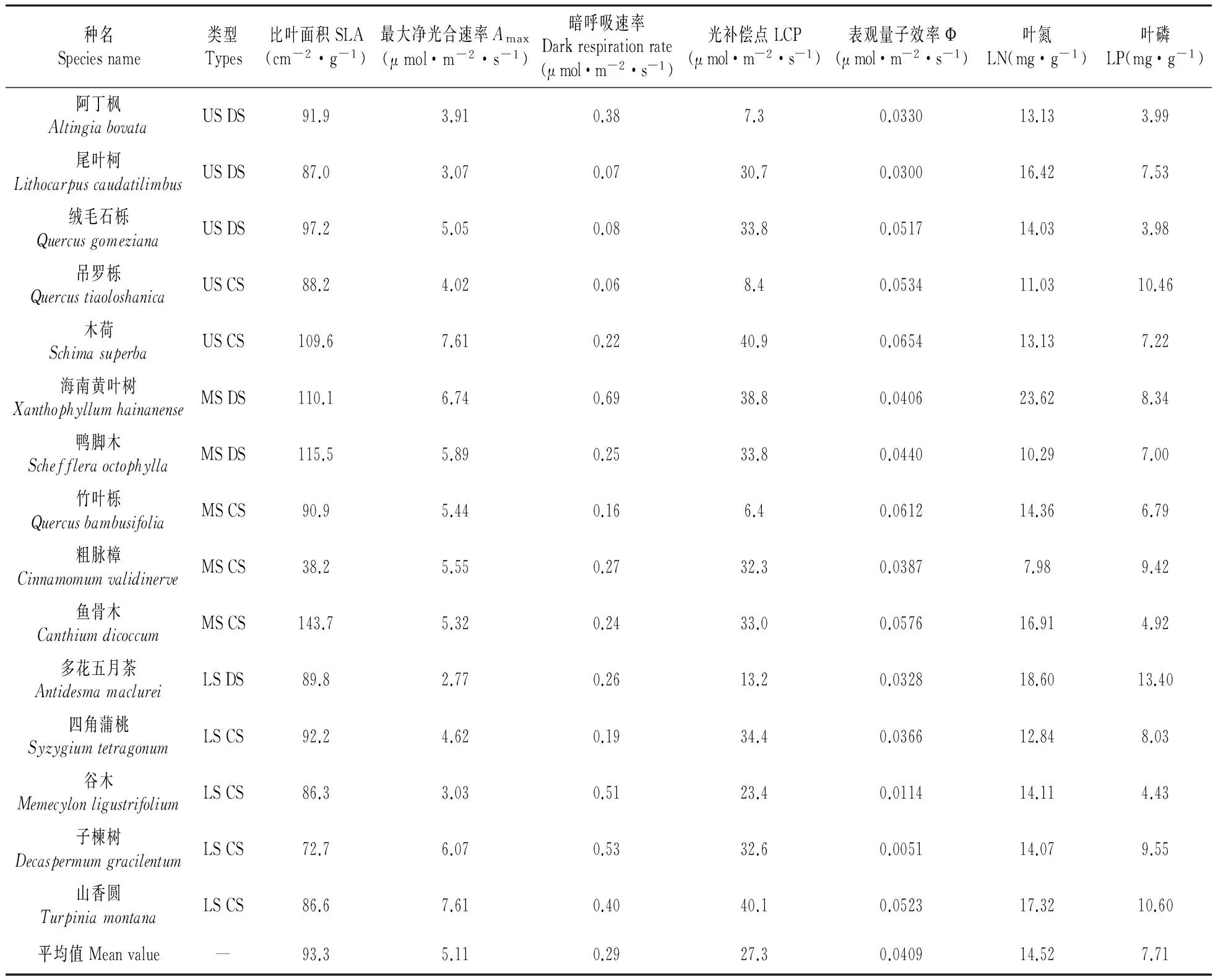
表1 不同层次优势种与伴生种的比叶面积和光合作用光响应参数及叶片养分含量
注:LCP.光饱和速率;Ф.表观量子效率;LN.叶氮;LP.叶磷 下同。
Note:LCP.Light compensation poin;Ф.Apparcnt quantum yield;LN.Leaf nitrogen;LP.Leaf phosphorus The same as below.
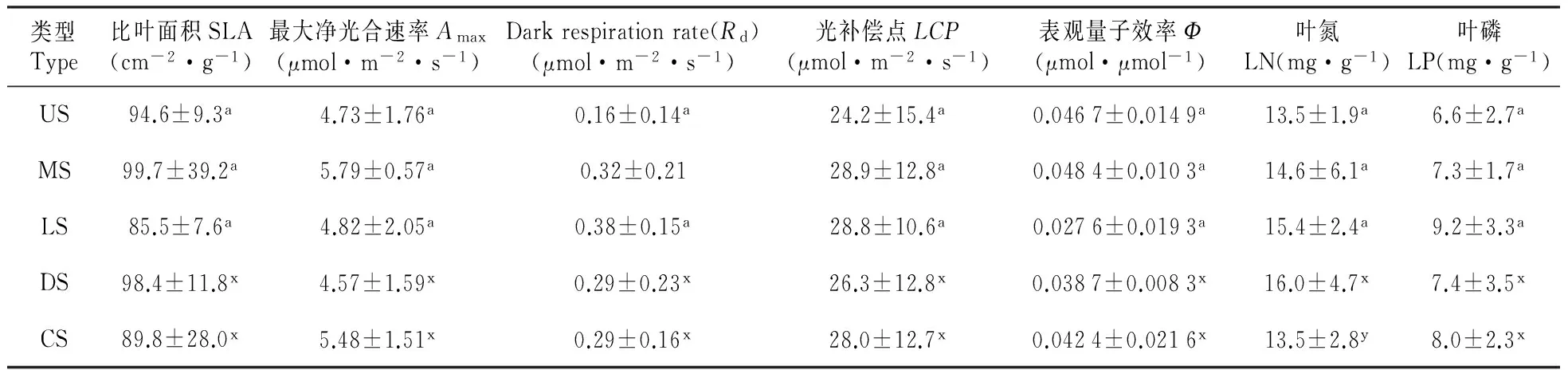
表2 不同类型树种的比叶面积、光合作用光响应参数和叶片养分含量比较
注:数据为平均值±标准误(n=5-9);标记相同字母的值差异不显著
Note:The data were the means±standard errors of separated experiments(n=5-9);Values followed by the same letter are not significantly different.
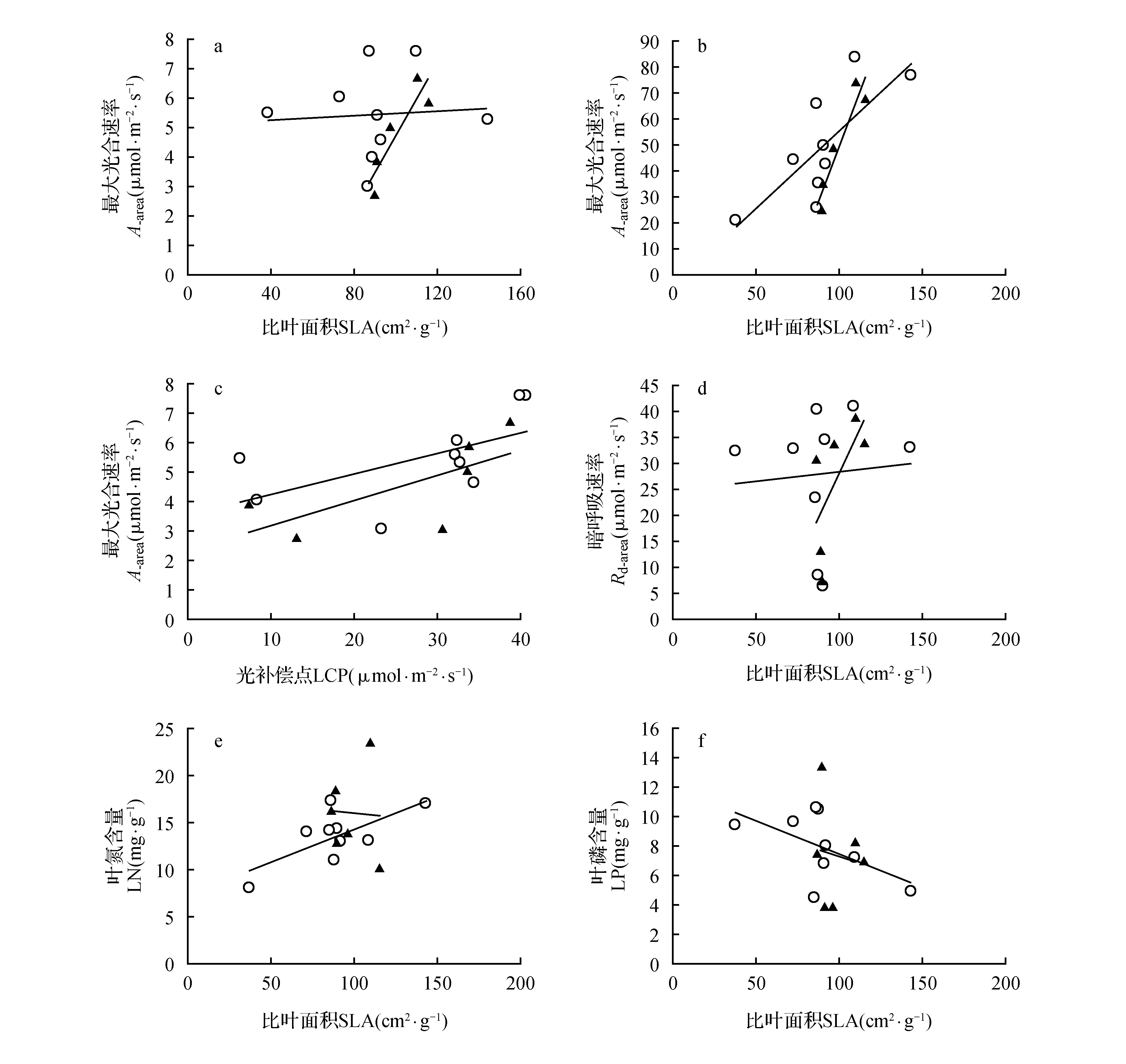
图1 海南岛热带雨林原始林优势种(▲)与伴生种(○)的最大光合速率、暗呼吸速率、叶磷含量、叶氮含量与比叶面积以及单位面积最大光合速率与光补偿点的关系Fig.1 The relationship of A-area,A-mass,Rd-area,leaf N(LN),leaf P(LP) vs. SLA and A-area vs. light compensation point(LCP) for dominant(▲) and companion(○) Hainan Island tropical forest tree species
3.3 优势种与伴生种间光合特征比较
优势种的比叶面积和单位干重暗呼吸速率大于伴生种,且不显著(P>0.05);前者的最大光合速率(A-area和A-mass)、单位面积暗呼吸速率、光补偿点、表观量子效率、叶片磷含量小于后者,也不显著(P>0.05)。优势种的叶片氮含量显著大于伴生种(P<0.05)(表2)。
经线性回归分析,优势种的最大光合速率(A-area和A-mass)与SLA显著正相关(P<0.01);伴生种的A-mass与SLA显著正相关(P<0.01)(图1)。优势种和伴生种的A-area和A-mass均与LCP显著正相关(P<0.01,P<0.05)。优势种的Rd-mass与SLA显著正相关(P<0.05)。伴生种的LN与SLA显著正相关(P<0.01);LP与SLA显著负相关(P<0.05),回归方程如表3所示。
4 讨论
植物群落组成不仅取决于其生长环境因素的作用,而且也受群落内物种间的相互作用的影响。在演替过程中,经过局部环境筛选的植物,都能适应其周围小生境的环境,从而进行繁殖与群落的更新。在此过程的作用下,功能相似的物种则处于相同的生态位,使得群落内各物种的特征趋于相同[23]。本研究中,各树种间SLA变化幅度不大,最高者鱼骨木的SLA约为粗脉樟的约3.7倍,与郑征等[24]和Santiago[25]等研究的热带雨林林下树种结果类似。结果表明,不同层次物种以及优势种与伴生种在适应林下低光环境时,在叶片性状上选择了相似的适应策略。
表3最大光合速率、暗呼吸速率、叶磷含量、叶氮含量与比叶面积以及单位面积最大光合速率与光补偿点的回归分析
Table3RegressionbetweenA-area,A-mass,Rd-area,leafN(LN),leafP(LP)andSLAandbetweenA-areaandlightcompensationpoint(LCP)fordominant(DS)andcompanion(CS)HainanIslandtropicalforesttreespecies
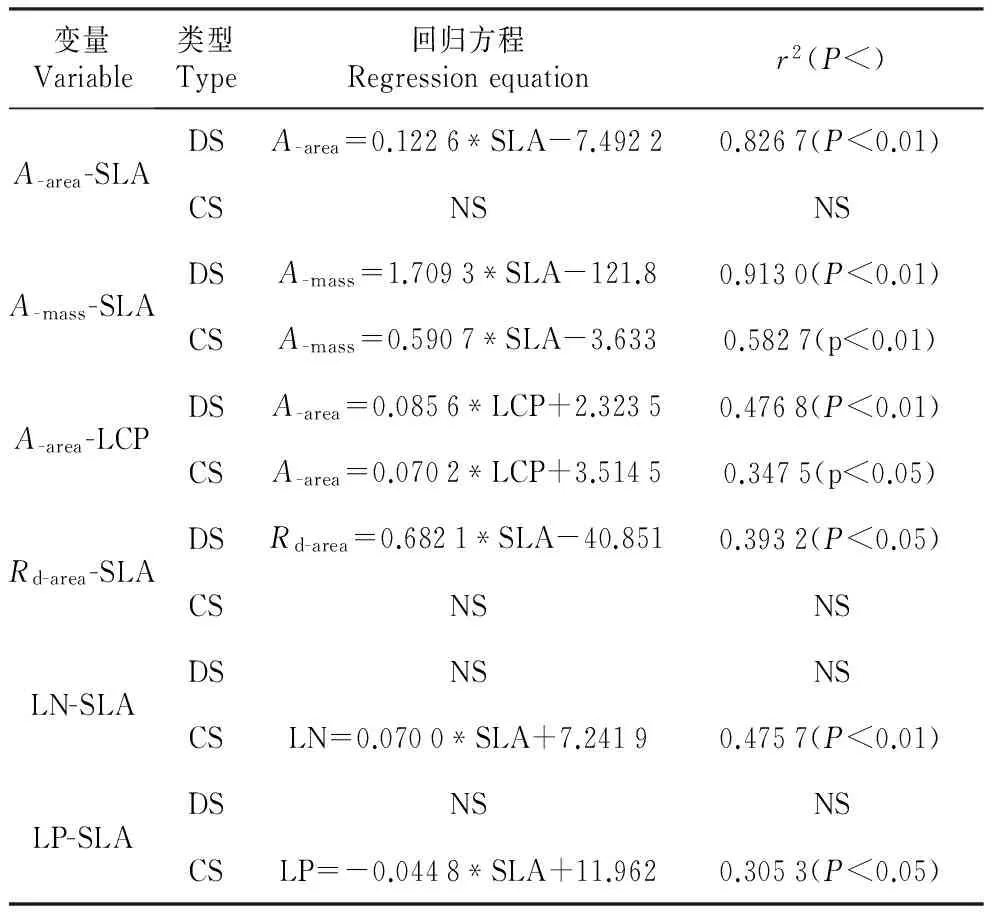
变量Variable类型Type回归方程Regressionequationr2(P<)A-area-SLADSA-area=0.1226*SLA-7.49220.8267(P<0.01)CSNSNSA-mass-SLADSA-mass=1.7093*SLA-121.80.9130(P<0.01)CSA-mass=0.5907*SLA-3.6330.5827(p<0.01)A-area-LCPDSA-area=0.0856*LCP+2.32350.4768(P<0.01)CSA-area=0.0702*LCP+3.51450.3475(p<0.05)Rd-area-SLADSRd-area=0.6821*SLA-40.8510.3932(P<0.05)CSNSNSLN-SLADSNSNSCSLN=0.0700*SLA+7.24190.4757(P<0.01)LP-SLADSNSNSCSLP=-0.0448*SLA+11.9620.3053(P<0.05)
注:NS表示相关关系不显著(P>0.05),因此未列出回归参数。
Note:NS indicates a non-significant relationship(P>0.05),and therefore no regression parameters are shown.
不同层次的物种在叶片光合特征上存在一定差异。从上层种到下层种,叶片氮、磷含量逐渐升高,下层种更趋于将养分分配于叶片中,以适应林下光环境[26]。叶片中氮、磷含量高,又使得下层种有较高的暗呼吸速率和光补偿点,因此其单位干重最大光合速率低;而下层种SLA低,叶片偏厚实,从而使其单位面积最大光合速率有所升高,略高于上层种。下层种的表观量子效率最低,对光的利用效率最低,与郑征[24]等研究的下层树种相似。中上层种对光的利用效率高,具有较高的光合能力[27~28],这也是其快速生长所必须的[29],因此中上层种更趋于将养分用于生长,从而降低了其叶片养分含量。
优势种与伴生种在叶片光合特征上也存在一定差异。优势种的氮含量显著高于伴生种,其比叶面积也高于伴生种。优势种将更多的氮含量分布于叶片,并增加其比叶面积,有助于在林下捕获光[30~31],是对林下光环境的一种适应;但叶片高氮使得呼吸变大,从而使最大光合速率降低。优势种的叶片氮、磷含量与SLA无相关关系,表现出其对林下环境的一种更灵活的适应。
尽管植物物种在生活型、叶片大小、叶片形状等方面都存在极大的多样性,但是在一定范围的物种之间存在着某种特定的联系,这种相似的相关关系是植物物种趋同进化的一种证明[32]。在本研究中,A-mass与SLA之间的相关关系在优势种和伴生种中都达到了显著水平,与Reich[33]等研究结果相似,强有力地支持了植物趋同进化的思想。
不同树种在遗传上的不同,决定了其所能达到分布层次的不同,因此幼树在生理上也表现不同的光合特征,对林下的低光环境有不同的适应策略。中上层种幼树在光合能力和对光的利用效率上显示优越性,但不同的中上层种的光合侧重点不同,养分利用上也不同。上层种幼树以低暗呼吸速率适应,中层种以高光合能力适应,而下层种以叶片高养分含量适应。优势种幼树在光合能力上并无优势,却更显示出更灵活的适应策略。不同次层树种在叶片生理特征上的分异,有利于形成复杂的群落结构和高生物多样性的维持。本文初步对海南岛热带雨林演替后期原始林不同层次优势种和伴生种的幼树在林下的光合作用参数和叶片特征进行比较研究,将为今后深入研究奠定基础。
5 结论
(1)中上层种的幼树具有较高的比叶面积、单位干重最大光合速率和表观量子效率;上层种的暗呼吸速率最低。
(2)幼树叶片的氮、磷含量随物种成树所在层次升高而减少,次序为下层种>中层种>上层种,但差异均不显著(P>0.05)。
(3)优势种和伴生种的光响应参数与SLA存在相关关系。伴生种的LN与SLA正相关(P<0.01),LP与SLA负相关(P<0.05);优势种的LN、LP与SLA不相关(P>0.05)。
1.Pielou E C.Species-diversity and pattern-diversity in the study of ecological succession[J].Journal of Theoretical Biology,1966,10(2):370-383.
2.Pianka E R.Evolutionary ecology[M].New York:Harper and Row,1974.
3.Connell J H.On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees[M].//Den Boer P J,Gradwell D R.Dynamics of populations.The Netherlands,Wageningen:Center for Agricultural Publishing and Documentation,1971:298-310.
4.Huston M.A general hypothesis of species diversity[J].The American Naturalist,1979,113(1):81-101.
5.Wilson E O.The diversity of life[M].Cambridge:The Belknap Press of Harvard University Press,1992.
6.安树青,王峥峰,曾繁敬,等.海南吊罗山热带山地雨林植物种类多样性研究[J].中山大学学报:自然科学版,1999,38(6):78-83.
An S Q,Wang Z F,Zeng F J,et al.Biodiversity of tropical mountane rain forest on Diaoluo Mountain,Hainan[J].Acta Scientiarum Naturalium Universitatis Sunyatseni,1999,38(6):78-83.
7.田大伦.高级生态学[M].北京:科学出版社,2008.
Tian D L.Advanced ecology[M].Beijing:Science Press,2008.
8.王刚,周本智,李正才,等.幕阜山森林群落结构与物种多样性研究[J].生态科学,2010,29(4):332-338.
Wang G,Zhou B Z,Li Z C,et al.Community structure and species diversity of forest communities in Mufu Mountain[J].Ecological Science,2010,29(4):332-338.
9.张明,王文进,刘福德,等.海南热带山地雨林幼苗幼树的光合能力与水分利用效率[J].应用生态学报,2007,18(10):2160-2166.
Zhang M,Wang W J,Liu F D,et al.Photosynthetic capacity and water use efficiency of tropical montane rainforest seedlings or saplings in Hainan Island[J].Chinese Journal of Applied Ecology,2007,18(10):2160-2166.
10.龙成.热带常绿季雨矮林优势种群和主要伴生种群结构、动态、空间分布格局及种间联结性研究[D].海口:海南大学,2013.
Long C.Research on structure,dynamic,spatial distribution pattern and interspecific association of dominant and associated populations in tropical evergreen monsoon elfin forest[D].Haikou:Hainan University,2013.
11.卢炜丽,张洪江,陈奇伯,等.重庆四面山杉木林群落乔木层优势种群生态位特征研究[J].西北林学院学报,2016,31(2):60-65.
Lu W L,Zhang H J,Chen Q B,et al.Niche characteristics of the tree-layer dominant populations ofCunninghamialanceolataforest in Chongqing Simian Mountains[J].Journal of Northwest Forestry University,2016,31(2):60-65.
12.申文辉,谭长强,何琴飞,等.桂西南蚬木优势群落物种组成及多样性特征[J].生态学杂志,2016,35(5):1204-1211.
Shen W H,Tan C Q,He Q F,et al.Species composition and diversity characteristics ofExcentrodendronhsienmu-dominated communities in southwestern Guangxi,China[J].Chinese Journal of Ecology,2016,35(5):1204-1211.
13.康瑶瑶,李瑞生,高永龙.王朗自然保护区岷江冷杉林优势种与主要伴生种的空间格局[J].西部林业科学,2015,44(2):48-53.
Kang Y Y,Li R S,Gao Y L.Spatial pattern of dominant and main companion tree species in anAbiesfaxonianaforest in Wanglang Nature Reserve[J].Journal of West China Forestry Science,2015,44(2):48-53.
14.朱慧,马瑞君.入侵植物马缨丹(Lantanacamara)及其伴生种的光合特性[J].生态学报,2009,29(5):2701-2709.
Zhu H,Ma R J.Photosynthetic characteristics comparison between an invasive plant,LantanacamaraL.,and associated species[J].Acta Ecologica Sinica,2009,29(5):2701-2709.
15.朱慧,马瑞君.入侵植物假臭草及其伴生种的光合特性[J].福建林学院学报,2010,30(2):145-149.
Zhu H,Ma R J.Comparison of photosynthetic characteristics between an invasive plant,Eupatoriumcatarium,and assoiated species[J].Journal of Fujian College of Forestry,2010,30(2):145-149.
16.王文进,张明,刘福德,等.海南岛吊罗山热带山地雨林两个演替阶段的种间联结性[J].生物多样性,2007,15(3):257-263.
Wang W J,Zhang M,Liu F D,et al.Species association in tropical montane rain forest at two successional stages in Diaoluo Mountain of Hainan Island[J].Biodiversity Science,2007,15(3):257-263.
17.王峥锋,安树青,Campell D G,等.海南岛吊罗山山地雨林物种多样性[J].生态学报,1999,19(1):61-67.
Wang Z F,An S Q,Campell C G,et al.Biodiversity of the montane rain forest in Diaoluo Mountain,Hainan[J].Acta Ecologica Sinica,1999,19(1):61-67.
18.张明,刘福德,王中生,等.热带山地雨林演替早期先锋树种与非先锋树种叶片特征的差异[J].南京林业大学学报:自然科学版,2008,32(4):28-32.
Zhang M,Liu F D,Wang Z S,et al.Differences of leaf traits between pioneer and non-pioneer tree species in early succession stage of tropical montane rain forest[J].Journal of Nanjing Forestry University:Natural Sciences Edition,2008,32(4):28-32.
19.Shipley B,Almeida-cortez J.Interspecific consistency and intraspecific variability of specific leaf area with respect to irradiance and nutrient availability[J].Écoscience,2003,10(1):74-79.
20.陈飞宇,罗天祥,张林,等.江西九连山常绿阔叶林主要树种叶建成消耗的比较[J].生态学报,2006,26(8):2485-2493.
Chen F Y,Luo T X,Zhang L,et al.Comparison of leaf construction cost in dominant tree species of the evergreen broadleaved forest in Jiulian Mountain,Jiangxi province[J].Acta Ecologica Sinica,2006,26(8):2485-2493.
21.鲍士旦.土壤农化分析:3版[M].北京:中国农业出版社,2000.
Bao S D.Soil and agricultural chemistry analysis:3rd ed[M].Beijing:China Agriculture Press,2000.
22.Prioul J L,Chartier P.Partitioning of transfer and carboxylation components of intracellular resistance to photosynthetic CO2fixation:a critical analysis of the methods used[J].Annals of Botany,1977,41(4):789-800.
23.Lavorel S,Garnier E.Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J].Functional Ecology,2002,16(5):545-556.
24.郑征,曹敏.西双版纳热带季节雨林6种树种幼树在林下的光合作用[J].植物生态学报,1998,22(6):499-504.
Zheng Z,Cao M.Photosynthetic response to light in understory for six species young trees of a tropical seasonal rain forest in Xishuangbanna[J].Acta Phytoecologica Sinica,1998,22(6):499-504.
25.Santiago L S,Wright S J.Leaf functional traits of tropical forest plants in relation to growth form[J].Functional Ecology,2007,21(1):19-27.
26.Warren C R,Adams M A.Distribution of N,rubisco and photosynthesis inPinuspinasterand acclimation to light[J].Plant,Cell & Environment,2001,24(6):597-609.
27.Thomas S C,Bazzaz F A.Asymptotic height as a predictor of photosynthetic characteristics in Malaysian rain forest trees[J].Ecology,1999,80(5):1607-1622.
28.Davies S J.Photosynthesis of nine pioneerMacarangaspecies from Borneo in relation to life history[J].Ecology,1998,79(7):2292-2308.
29.Thomas S C.Asymptotic height as a predictor of growth and allometric characteristics in Malaysian rain forest trees[J].American Journal of Botany,1996,83(5):556-566.
30.Cornelissen J H C,Diez P C,Hunt R.Seedling growth,allocation and leaf attributes in a wide range of woody plant species and types[J].Journal of Ecology,1996,84(5):755-765.
31.Wright I J,Westoby M.Cross-species relationships between seedling relative growth rate,nitrogen productivity and root vs leaf function in 28 Australian woody species[J].Functional Ecology,2000,14(1):97-107.
32.Wright I J,Reich P B,Westoby M.Strategy shifts in leaf physiology,structure and nutrient content between species of high- and low-rainfall and high- and low- nutrient habitats[J].Functional Ecology,2001,15(4):423-434.
33.Reich P B,Ellsworth D S,Walters M B,et al.Generality of leaf trait relationships:a test across six biomes[J].Ecology,1999,80(6):1955-1969.
Special Fund of National Environmental Protection for Non-profit Industry(201409055)
introduction:ZHANG Ming(1982—),male,master,research assistant,mainly engaged in the study of ecological restoration and protection.
date:2017-04-24
LeafTraitsoftheSaplingsofLaterSuccessionalStageTropicalMontaneRainForestinHainanIsland
ZHANG Ming1,2LIU Fu-De3AN Shu-Qing2CAO Xue-Zhang1*
(1.Nanjing Institute of Environmental Sciences,Ministry of Environmental Protection,Nanjing 210042; 2.School of Life Science,Nanjing University,Nanjing 210093; 3.Environmental Science and Safety Engineering,Tianjin University of Technology,Tianjin 300384)
Physiological and ecological characteristics of plant greatly affect its distribution pattern and abundance in ecosystem. The objectives of this study are to find differences of leaf traits between dominant species and companion species, and differences among three different layers. By sample-plot survey, leaf traits of six dominant species and nine companion species from three different layers of later successional stage tropical montane rain forest were measured in Hainan Island, by using a Li-6400 portable photosynthesis system. Leaves were sampled to measure its specific leaf area(SLA), leaf nitrogen(LN) and leaf phosphorus(LP). SLA(specific leaf area),A-mass(light-saturated photosynthetic rate) and Ф(photosynthetic quantum yields) of middle and upper layer species were higher than those of lower layer species, and the upper layer species had a lowestRd(day-time dark respiration rate) among the three groups. LN and LP of saplings decreased with their mature tree height increasing, and had the order of lower layer species>middle layer species>upper layer species, but there was no significant differences among there groups(P>0.05). Photosynthetic parameters of dominant and companion species were correlated with SLA. LN of companion species was positively correlated with SLA(P<0.01), but their LP was negatively correlated with SLA(P<0.05). But for dominant species, there was no correlation between LN and SLA or LP and SLA(P>0.05).
tropical montane rain forest;later successional stage;dominant species;companion species;photosynthesis;specific leaf area;leaf nitrogen;leaf phosphorus
国家环境保护公益性行业科研专项(重要生态功能区退化生态系统修复模式研究与应用示范201409055)
张明(1982—),男,硕士研究生,助理研究员,主要从事生态修复与保护研究。
* 通信作者:E-mail:caoxuezhang@126.com
2017-04-24
* Corresponding author:E-mail:caoxuezhang@126.com
S718.5
A
10.7525/j.issn.1673-5102.2017.06.014
猜你喜欢
空间科学学报(2021年2期)2021-07-21
空间科学学报(2020年2期)2020-04-01
河北果树(2020年1期)2020-02-09
现代园艺(2017年11期)2017-06-28
少年文艺·开心阅读作文(2017年1期)2017-02-24
考古与文物(2016年5期)2016-12-21
现代企业(2015年6期)2015-02-28
小天使·四年级语数英综合(2014年3期)2014-03-21
中国果业信息(2013年7期)2013-01-22
中国移动周刊(2009年46期)2009-05-13
