
闽北闽粤栲天然林高度级结构分析
2017-12-19 06:12缪三华郑德祥蔡杨新许鲁东范胜煌詹旋常钟兆全
中南林业科技大学学报 2017年6期
缪三华,郑德祥,蔡杨新,许鲁东,范胜煌,詹旋常,钟兆全
(1.福建农林大学 林学院,福建 福州 350002;2.顺昌国有林场,福建 顺昌 353200)
闽北闽粤栲天然林高度级结构分析
缪三华1,郑德祥1,蔡杨新1,许鲁东1,范胜煌1,詹旋常2,钟兆全2
(1.福建农林大学 林学院,福建 福州 350002;2.顺昌国有林场,福建 顺昌 353200)
在福建省北部顺昌县的闽粤栲天然林林分中设置50 m×50 m 样地,分别不同高度级调查群落物种种类与数量,运用失稳率、分享度、α多样性指标、Whittaker指数和Morista-Horn指数对闽粤栲天然林群落高度级结构与动态更新规律进行研究。研究结果表明:在闽北闽粤栲天然林群落中,物种数及个体数沿高度级梯度上升而下降,除第2高度级受前期林下抚育影响外,闽粤栲及其他主要树种在第1~3高度级中占有较大优势,群落间失稳率达到50%以上,而整体失稳率达到10.65%,群落物种多样性指数与高度级呈负相关关系,反映群落更新速率的Whittaker指数随着高度级呈递增趋势,Morisita-Horn指数则在较高值域不均匀波动,以上结果均表明在闽北闽粤栲天然林随高度级上升,各高度级间物种数下降,物种更新速率加快,各高度级间差异明显,群落整体处于演替发育阶段。
闽粤栲;群落;高度级;天然林;更新演替
在植物生态学领域里,所有植物种在其生境内空间和时间上的配置状况属于群落结构研究的重要组成部分,涉及到物种关系、多样性表达以及群落演替等诸多方面,对森林可持续经营和生物多样性保护具有指导意义[1-7]。高度级结构借助高度等级来划分植物群落,以空间尺度代替时间尺度定量分析群落空间垂直结构,科学揭示各种群不同发展阶段的生态学特征,是当前研究群落结构的一种行之有效的手段[8-10]。前人经过一系列的植物种群诸如丝栗栲、青冈、胡杨等的高度级结构研究[11-13],也认为高度级结构研究不仅能客观描述植物个体及其种群在群落垂直结构中的配置、地位和作用[8],定量分析群落高度级结构对把握整体群落动态特征[14],推测群落演替发展方向,指导森林生态可持续发展具有重要意义。
闽粤栲Castanopsis fi ssa为壳斗科Fagaceae栲属树种,又名黧蒴栲、大叶栲等,其性喜光暖、适应能力强,萌芽能力旺盛、根系发达,固土保水能力强,是南方集体林区中亚热带常绿阔叶林林常见主要树种之一[15]。目前关于闽粤栲的研究仅见于幼苗栽培[16]、遗传多样性分析[17]以及森林经营[18]等方面。至于生态学方面鲜有报道[15,19]。基于此,本文以闽北常见闽粤栲天然林为研究对象,对其进行群落高度级结构研究,以期为闽粤栲天然林的保护与可持续利用提供理论依据。
1 材料与方法
1.1 研究区概况
闽粤栲天然林在福建省闽北地区分布较为广泛,本研究在闽北顺昌县国有林场经营区内选择保护良好的闽粤栲天然林林分进行调查研究(基本情况见表1)[14]。顺昌县地处福建省西北部,位闽江上游金溪和富屯溪的交汇处,地理坐标为东经117°29′~ 118°14′,北纬 26°38′~ 27°12′。中亚热带海洋性季风气候,同时受到一定的大陆性气候影响,年平均气温为19.2℃,年平均日照时间为1 736.3 h,无霜期305 d,年平均降水量1 738 mm,冬多西北风,夏多东南风。在所调查的林分样地中乔木层的主要树种有闽粤栲Castanopsis fi ssa、木 荷Schima superba、 米 槠Castanopsis carlesii、丝栗栲Castanopsis fargesii、黄毛润楠Machilus chrysotricha、青冈Cyclobalanopsis glauca等。

表1 调查区基本情况Table 1 Summary of the survey district
1.2 样地的设置与调查
在闽粤栲天然林林分的阴坡和阳坡分别设置50 m×50 m的样地,调查样方内各物种数量、种类等,调查前期通过咨询植物学专家、查阅前人研究报道[20],本研究组参考李俊清等编著的《森林生态学》[21]中对森林植物群落垂直结构的划分标准,按照3 m高度级单位标准,采用上限排外法从下而上(1~k级)进行高度级划分[22],量测记录样方内所有植株的高度与直径,统计各高度级的数量特征,总体调查情况见表2。

表2 闽粤栲天然林高度级物种数量调查Table 2 The survey of species number in different height levels of the Castanopsis fissa natural forest community
1.3 高度级结构指标
1.3.1 失稳率
种群的整体失稳率(population lost stable rate,即Vp)等于各级失稳率[]的加权平均值:
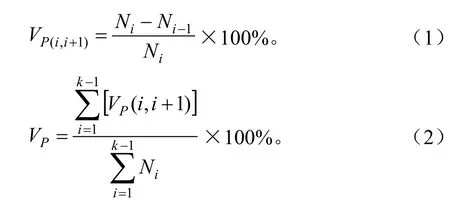
式中:和分别表示第i和第i+1个高度级的种群个体数,k代表此物种高度级的最高等级。
对于群落而言,群落高度级间失稳率VC(i,i+1)和整体失稳率VC应是所属优势种群级间失稳率或整体失稳率的连乘积。即:
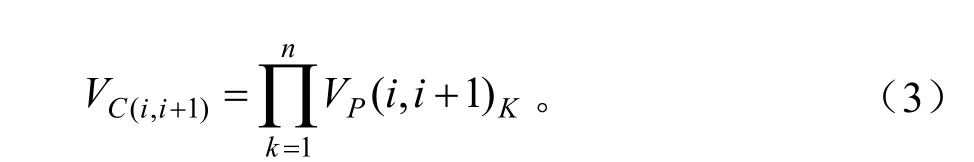
式中:n代表群落中的优势树种类数,k代表第i个优势树种。同时规定当出现一个以上VP(i,i+1)或VP为负时,VC(i,i+1)或VC取负值。
1.3.2 群落物种多样性
基于高度级垂直结构,本研究以α多样性指标对闽粤栲群落物种多样性进行分析:

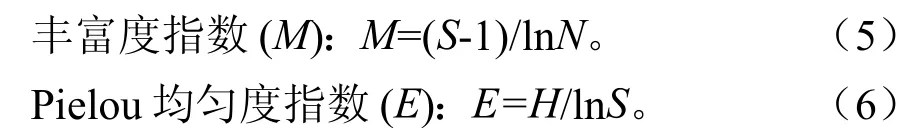
Shannon-Wiener指数 (H):式中:S代表群落高度级上的物种数量,代表物种相对多度,代表种i的个体数,N代表高度级内的总个体数[23]。
1.3.3 分享度
物种分享度 (species sharing degree)以某一高度级物种的种数()占整个群落物种数(S)的比重来表达[24]。

群落整体分享度(community sharing degree,Cssd)以各高度级分享度的均值表示:
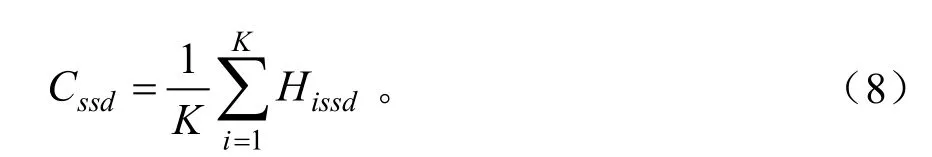
1.3.4 群落高度级物种更替速率
本 研 究 采 用 Whittaker指 数 (βw) 和 Morista-Horn指数(C)两种不同类型的数据指标对群落高度级物种更替速率[24-26]。其中:Whittaker指数(βw)是较为经典的二元属性数据指标,侧重于检测两个数据组间的组成元素差异性,将其与高度级相结合,公式如下:

式中:S和mα分别为相邻两个高度级的出现的所有物种数和共有物种数,其值越小,物种更替速率越小。
Morista-Horn指数(C)在本研究中主要是用来描述相邻高度级间关于物种数量差异程度的数量数据指标,其值越大,差异程度越大,公式如下:

式中:S代表A和B高度级中的所有物种数;aNi和bNi是A和B高度级中第i种的个体数;aN和bN是相对应高度级中的物种数。
2 结果与分析
2.1 高度级个体结构特征及其动态分析
2.1.1 各高度级个体数分布特征
鉴于彼此高度级间个体数量的差异较大,将各高度级上种群及群落个体数比例制成条形图以分析个体数分布特征。如图1所示:

图1 闽粤栲天然林和种群个体—高度图Fig. 1 Individual-highness proportion of Castanopsis fi ssa community and its dominant population
闽粤栲天然林群落个体数主要集中分布于第1~3高度级上,占总体的91.45%,随着高度级梯度上升而下降。除在第2高度级外(其原因是前期抚育间伐中,由于闽粤栲成材材质常出现空心现象,而使其经济价值受到经营者质疑,而成为抚育采伐对象所致),建群优势种闽粤栲个体数分布与群落整体的个体数分布具有高度的相似度,尤其是第1高度级中闽粤栲的个体数比重高达46.49%,表明在闽粤栲天然林分中闽粤栲幼树幼苗数量众多,表现出年轻而富有潜力的生命动态,其有利于闽粤栲的后续更新,根据生态学理论研究图1时,作为K对策者,闽粤栲天然林种群在群落演替发展的初期阶段,为了进一步巩固自身的优势地位,预防资源被其它竞争者掠夺,闽粤栲种群会以较快的速度从第1高度级发展开来,这与本研究的调查分析结果基本一致[25]。
2.1.2 个体数结构动态分析
群落结构的发展方向以种群的稳定性为基础,高度级结构的优势种群失稳率是衡量群落结构增长、衰退和稳定的可靠性指标,分别以失稳率值的正负值和零作为判定依据[26]。本次调查的闽北闽粤栲天然林群落为多优群落,优势树种主要有闽粤栲、木荷以及米槠,三者高度级间失稳率以及群落失稳率如表3所示。
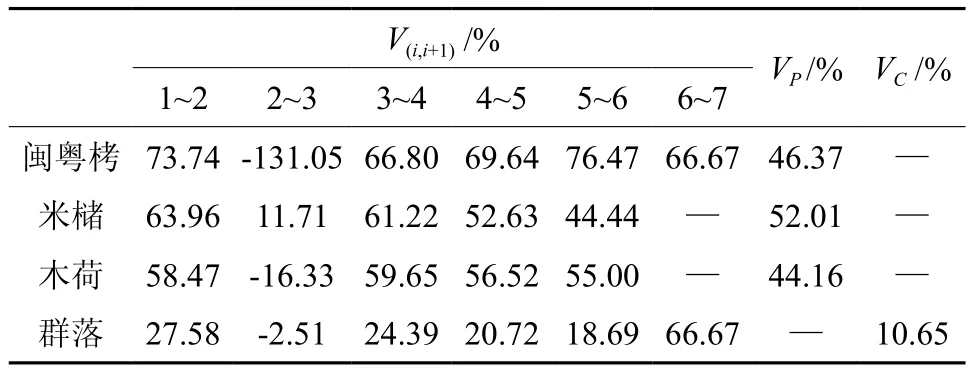
表3 闽粤栲群落高度级个体数结构动态Table 3 Dynamics of the individual structure of Castanopsis fissa community highness class
从结果来看,各优势种种群和群落整体在高度级2到3之间的失稳率异常印证了抚育间伐作业对群落组成格局存在明显干扰作用的事实。除此之外,闽粤栲群落内的优势种群整体失稳率值均为正值,具备增长性结构,三者正朝着加快现有群落组成格局稳定的方向发展,群落正处于未成熟的发展阶段。群落内优势种群彼此间的失稳率数值虽然相近,但参考相关种群在高度结构上的个体密度差异,据此推测,未来闽粤栲种群在群落中的优势地位可能会随着幼苗幼树的生长而得以较大幅度提升。
2.2 群落高度级物种多样性分析
为进一步分析群落物种结构与分布格局,分别计算闽粤栲天然林群落在各高度级上物种α多样性指标值见表4。
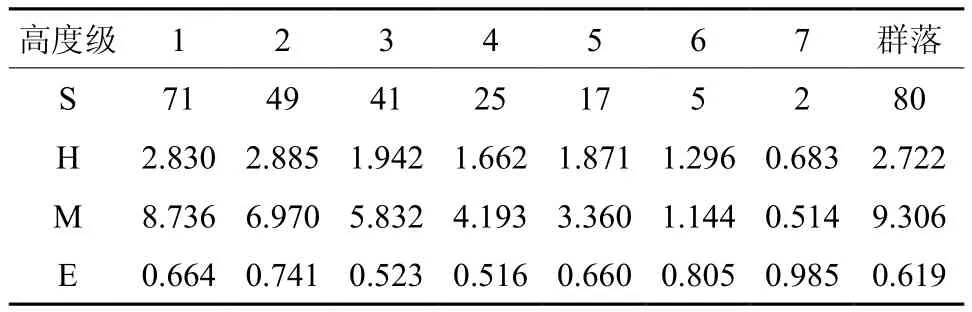
表4 闽粤栲群落高度级物种α-多样性†Table 4 The species α-diversity in different highness of the Castanopsis fissa natural forest community
在本研究地,虽然森林的经营管理和气候环境变化或多或少会对群落演替产生不同程度的干扰,但受益于森林生态系统强大的自我恢复功能,闽粤栲天然林群落总是朝着物种多样、结构稳定的方向发展。在第1~2高度级上多样性指数(H)和丰富度指数(M)明显高于之后的高度级相对应的指标值,从物种结构上进一步印证了该群落处于稳定增长阶段这一事实。从整体上看,群落多样性指数和丰富度指数在高度级梯度上是呈下降趋势的,这符合天然林群落的自然生长发育规律。至于表4中多样性指数(H)在第5高度级上出现的异常波动现象,这和生产经营过程中对干形较好或特殊的个别林木的保护有关,譬如群落中的青刚栎、杜英Elaeocarpus decipiens、杜鹃Rhododendron simsii、漆树Toxicodendron vernici fl uum等物种在此高度级上的分布就较为集中。受前期人工抚育干扰,除第2高度级外,第1~3高度级的均匀性指数(E)值呈现不规则波动,而从第4高度级开始,随着高度级梯度的上升,物种数不断减少,均匀性指数不断提高,均匀度在第7高度级甚至接近于1,究其原因是处于增长繁荣阶段的种群个体间的生态位竞争加剧,物种数急剧下降,上层林木竞争优势凸显与林木自然稀疏作用有关[27]。
2.3 高度级物种分享度分析
分享度能够反映群落物种在高度级上的配置分布,从宏观上描述物种组成在高度级梯度上的变化状况。计算天然闽粤栲林分各高度级的分享度并制成折线趋势分析图,如图2。
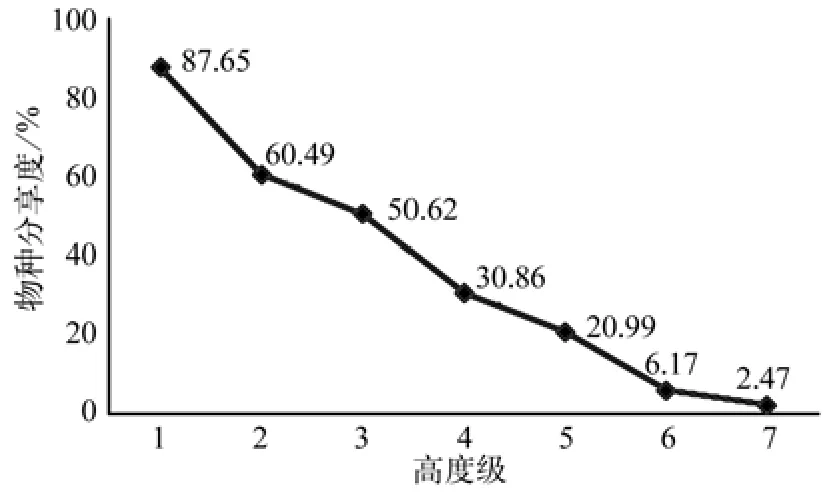
图2 闽粤栲天然林群落高度级结构物种分享度折线图Fig.2 The species sharing curve in different highness of the Castanopsis fi ssa natural forest community
在闽粤栲天然林分中,第1高度级上群落物种分享度高达87.65%,绝大多数物种在此高度级上均有分布,其中闽粤栲幼苗、米槠、木荷以及山茶Camellia japonica等物种更是展现出强大的生态适应性。从第2级起,受限于物种自身生物学特性以及前期营林抚育生产的影响,许多物种在这一高度级中直接消失或出现断层现象,如山血丹Ardisia lindleyana、红皮树Styrax suberifoltus、小叶黄杨Buxu sinica、菝契Smilax china、桃叶石楠Photinia prunifolia、交让木Daphniphyllum macropodum以及杜仲Eucommia ulmoides等,分享度直降至60.49%。群落中以山茶为代表的灌木由于自身生长高度问题也在第3高度级之后相继消失,受林木的生物学和生态学特性影响,生态位宽度大的乔木树种如如闽粤栲、米槠和木荷三个物种的优势逐步随着高度级梯度往上而突显,而其他更多的物种被迫在更高的高度级上淡出,如中华杜英Elaeocarpuschinensis、黄润楠Machilus chrysotricha、光叶石楠Photinia glabra、南岭栲Castanopsis fordii、大叶樟Cinnamomum parthenoxylon、青冈栎等。从整体而言,物种分享度沿着高度级梯度逐级减小,符合增长型群落结构的物种分布特征,群落的整体物种分享度达37.04%。高水平分享度也从侧面表现出群落优势物种分布具有较好连续性,生态系统趋于稳定。
2.4 群落高度级物种更替动态分析
为进一步分析群落物种结构的动态变化趋势,本研究结合二元属性数据指标Whittaker指数(βw)和数量数据指标Morista-Horn指数(C)对闽粤栲群落高度级间的物种更替速率进行测算,如图3所示。
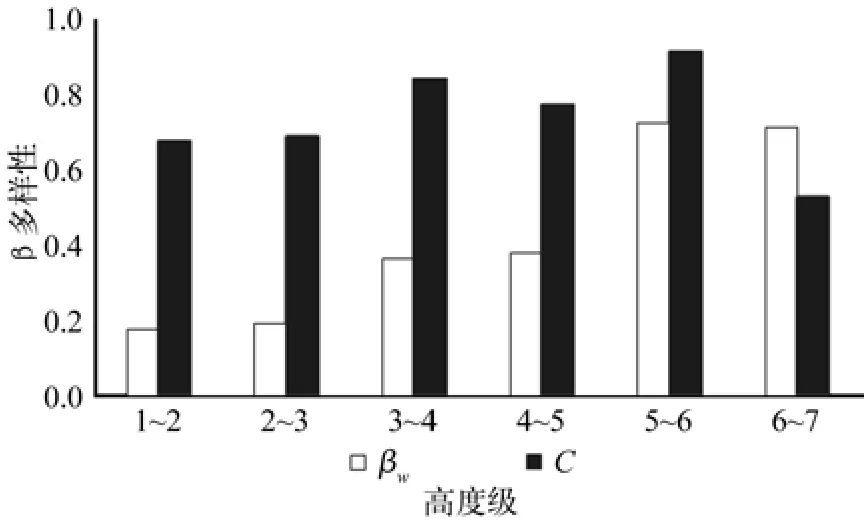
图3 相邻高度级β-多样性Fig.3 β-diversity of neighboring highness class
在群落前4个高度级中,代表高度级间物种更替速率的 Whittaker指数维持在较低且相近的水平,高度级间物种变化幅度相对平稳。从第3~4高度级起,生物学特性限制开始起显著作用,尤其是灌木树种绝大数开始绝迹,如山茶、檵木Loropetalum Chinensis、黄瑞木Adinandra millettii、中华杜英Elaeocarpuschinensis等。同时群落种内种间竞争在另一方面加剧了高阶高度级上的物种分化,随着高度级梯度的提升,物种种数下降,林木自我稀疏现象出现,幸存物种在高度级上所占比重不断上升,Whittaker指数不断提高,物种更替速率加快。受不同高度级的物种种类与数量变化影响,相邻高度级间的共有种数量差异明显,Morista-Horn指数在整个群落高度级间始终表现出在高指数值上的波动,均表明各高度级间的较大的物种差异性。结合Whittaker指数变化表明,除人为抚育干扰之外,闽北闽粤栲天然林群落的物种更替速率与高度级梯度呈正相关关系,各高度级间的物种数与个体数差异明显,整个群落生态系统的仍处于演替发育阶段。
3 小结与讨论
研究结果表明,闽北闽粤栲天然林的演替更新过程符合符合天然林演替规律。该群落内个体数和物种数的配置沿着高度级梯度递减,低阶高度级上的数量优势是增长型群落的典型特征。同时物种多样性指标在低阶高度级上高数值表现也很好地说明闽粤栲天然群落资源丰富,可以迎合不同物种的多种生态位需求。群落结构的研究需要动静结合,研究中的群落失稳率和物种分享度分别针对个体和物种在高度级间的变化情况展开评价,认为群落内优势种种群结构具备增长属性,整个群落的物种分布具备良好的连续性。这是增长型群落存在的普遍现象,于军等在对不同龄组胡杨种群高度级结构的研究中也得到相似结论[13]。另外,研究在原有物种丰富度、均匀度以及Shannon-Winner指数等描述群落整体多样性的基础上,利用Whittaker指数和Morista-Horn指数的二元属性分析高度级间物种组成差异程度,进一步确定了粤栲群落物种演替更新速率快的动态属性,也从侧面反映出群落物种间竞争激烈,越往高阶高度级,种群及其个体间生态位争夺越发频繁[27]。群落的演替过程是一个持续而缓慢的过程,在此过程中可能受到各种内外部因素的综合影响,具体原因还有待进一步探索。
在本研究中,由于人为抚育干扰就对闽粤栲天然林群落高度级结构产生了一定的影响,由于抚育间隔期限短,目前暂未影响群落整体发展趋势。同时尽管受限于调查区域与资料分析有限,但经过研究论证,高度级结构的分析对于客观揭示闽粤栲群落结构的部分特征和动态趋势,指导物种多样性保护和森林生产经营还是具有积极意义的。至于其整体区域及森林类型的规律仍有待于进一步调查研究。
[1]沙 威,董世魁,刘世梁,等. 阿尔金山自然保护区植物群落生物量和物种多样性的空间格局及其影响因素[J]. 生态学杂志,2016, 35(2):330-337.
[2]宋 超,余琦殷,于梦凡,等. 北京地区黄檗种群数量结构及空间分布特征[J]. 生态学杂志,2015, 34(11):3040-3049.
[3]刘圣恩,林开敏,蔡锰柯,等. 近自然生态恢复条件下杉木老龄林群落优势树种种群结构与空间格局[J]. 应用与环境生物学报,2015(3):540-546.
[4]陈亚南,杨 华,马士友,等. 长白山2种针阔混交林空间结构多样性研究[J]. 北京林业大学学报,2015, 37(12):48-58.
[5]Alicia Ledo,Sonia Condés,Iciar Alberdi. Forest Biodiversity Assessment in Peruvian Andean Montane Cloud Forest[J].Journal of Mountain Science,2012,9(3):372-384.
[6]张梦弢,张 青,亢新刚,等.长白山云冷杉林不同演替阶段群落稳定性[J]. 应用生态学报,2015,26(6):1609-1616.
[7]刘万德,李帅锋,张志钧,等. 滇西北云南红豆杉群落结构与更新特征[J]. 生态学杂志,2012, 31(12):3024-3031.
[8]龚 辉. 9年生邓恩桉河岸人工林林木高度自然分化特征研究[J]. 西南林业大学学报,2016, 36(1):74-78.
[9]毕晓丽,洪 伟,吴承祯,等. 万木林自然保护区不同群落优势种高度结构研究[J].应用生态学报,2003, 14(12):2355-2358.
[10]李帅锋,刘万德,苏建荣,等. 普洱季风常绿阔叶林次生演替中木本植物幼苗更新特征[J]. 生态学报,2012,32(18):5653-5662.
[11]宋 萍,洪 伟,吴承祯, 等. 中亚热带丝栗栲次生林群落高度级结构分析[J]. 应用与环境生物学报,2003, 9(5):460-464.
[12]刘剑斌. 闽北中亚热带青冈次生林高度结构分析[J]. 亚热带农业研究,2014, 10(4):247-251.
[13]于 军,白冠章,梁继业,等. 塔里木河上、中、下游胡杨种群高度结构特征[J]. 干旱区资源与环境,2012, 26(7):103-109.
[14]Pielou. Mathematical Ecology[M].New York: Wiley-Interserence,1985.
[15]苏凌燕,郑德祥,钟兆全,等.闽北闽粤栲天然林林隙特征及干扰状况[J]. 森林与环境学报,2015,35(2):125-130.
[16]宋 倩,卜朝阳,卢家仕,等. 不同基肥处理对黧蒴栲幼苗生理特性的影响[J]. 农学学报,2015(6):52-58.
[17]刘振华,童方平,蒋 燚,等. 黧蒴栲遗传多样性的AFLP分析[J]. 中国农学通报,2014(4):42-48.
[18]李 贵,童方平,刘振华,等. 黧蒴栲生长过程及数量成熟、工艺成熟的初步研究[J]. 中南林业科技大学学报,2013,33(12):53-56.
[19]蓝文升,钟兆全,郑德祥,等. 闽北天然闽粤栲种群空间分布格局研究[J]. 北华大学学报(自然科学版),2014,15(1):105-108.
[20]宋享镇. 三种采伐更新模式对米槠群落高度级结构的影响[D].福州: 福建农林大学,2008.
[21]李俊清. 森林生态学:第二版[M].北京: 高等教育出版社,2010.
[22]郑德祥,苏凌燕,钟兆全,等. 闽北闽粤栲天然林林隙物种多样性变化分析[J]. 西北农林科技大学学报(自然科学版),2016,(12):81-87.
[23]金 慧,赵 莹,赵 伟,等. 长白山牛皮杜鹃群落物种多样性的海拔梯度变化及相似性[J]. 生态学报, 2015,35(1):125-133.
[24]胡喜生,洪伟,吴承祯,等. 长苞铁杉群落优势种群高度生态位研究[J]. 广西植物,2004, 24(4):323-328.
[24]王世雄,王孝安,郭 华. 黄土高原植物群落演替过程中的β多样性变化[J]. 生态学杂志,2013, 32(5):1135-1140.
[25]WhittakerRH. Evolution and measurement of species diversity[J].Taxon, 1972, 21(2/3):213-251.
[26]SemeniukV, CresswellID. A Proposed Revision of Diversity Measures[J]. Diversity, 2013, 5(3):613-626.
[27]李俊清,等. 森林生态学:第二版[M].北京:高等教育出版社,2010.
Highness class structure of natural Castanopsis Cissa stand in Northern Fujian
MIAO Sanhua1, ZHENG Dexiang1, CAI Yangxin1, XU Ludong1, FAN Shenghuang1, ZHAN Xuanchang2, ZHONG Zhaoquan2
(1.College of Forestry, Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China;2.State-owned Forest Farm of Shunchang, Shunchang 353200, Fujian, China)
A 50 m×50 m sample plot was set to investigate the types and number of community species in each highness class of natural Castanopsis fi ssa standin Shunchang, Northern Fujian. Then population lost stable rate, species sharing degree, α-diversity index,Whittaker index and Morista-Horn index were applied to analyze theindividual distribution and regeneration dynamics for each highness class. The results showed that the individual number of each species was decreased with the raising highness class gradient except the 2nd which was in fl uenced by understory tending. Castanopsis fi ssa and some other main tree species were dominated in the fi rst three highness class. The population lost stable rate was high (over 50%) between each highness class, and in the whole community it was 10.65%. The community species diversity index and highness class had a negative correlation, indicating an uptrend of Whittaker index(i.e. used to re fl ect the community regeneration rate) with the raising highness class gradient. The Morisita-Horn index was unevenly fl uctuated in higher value range. The study results indicated that, in the natural Castanopsis fi ssa stand, with the raising highness class,the species number was decreased while the species regeneration rate was accelerated. The species number was obvious different in each highness class, the whole community of natural Castanopsis fi ssa stand was in succession stage.
Castanopsis fi ssa; community; highness class; natural stand; succession
S718.5
A
1673-923X(2017)06-0045-06
10.14067/j.cnki.1673-923x.2017.06.007
2016-07-26
福建农林大学校重点项目建设专项(6112C035H);国家自然科学基金项目(31170513)
缪三华,硕士研究生
郑德祥,副教授,博士;E-mail:fjzdx@126.com
缪三华,郑德祥,康海军,等. 闽北闽粤栲天然林高度级结构分析[J].中南林业科技大学学报,2017, 37(6): 45-50.
[本文编校:吴 彬]
猜你喜欢
浙江林业科技(2022年4期)2022-12-29
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
农村百事通(2020年21期)2020-11-23
绿色中国(2019年19期)2019-11-26
绿色中国(2019年19期)2019-11-26
铁军(2019年11期)2019-11-11
红土地(2019年10期)2019-10-30
