
越冬长耳鸮(Asio otus)食物分析
——以上海江苏两地样本为例
2017-12-14 07:28:01王军馥李必成薄顺奇唐思贤
复旦学报(自然科学版) 2017年4期
王军馥,李必成,薄顺奇,何 鑫,杨 刚,袁 晓,唐思贤
(1. 上海科技馆 自然史研究中心,上海 200127;2. 上海市野生动植物保护管理站,上海 200023;3. 华东师范大学 生命科学学院,上海 200062)
研究简报
越冬长耳鸮(Asiootus)食物分析
——以上海江苏两地样本为例
王军馥1,李必成1,薄顺奇2,何 鑫1,杨 刚1,袁 晓2,唐思贤3
(1. 上海科技馆 自然史研究中心,上海 200127;2. 上海市野生动植物保护管理站,上海 200023;3. 华东师范大学 生命科学学院,上海 200062)
2010—2012年、2014—2015年,采用食团内容物分析法分别对上海崇明、江苏盐城越冬长耳鸮的食物组成进行分析.两个越冬地3年时间内共收集完整食团717份,辨认出7种804只动物个体,其中鼠类、麝鼩等小型哺乳动物4种,占总数的93.7%,雀形目鸟类3种,及少量无法鉴定到种的鸟类.结果表明,两地越冬的长耳鸮对黑线姬鼠(Apodemusagrarius)的捕食量最大,捕食数量占食物总数的89.2%;对黑线姬鼠成年Ⅰ组的捕食比例最高,为40.2%;对黑线姬鼠各年龄组的捕食量与其当地种群数量变化基本一致.由此推测,越冬长耳鸮捕食的主要对象与周围环境中可获得的食物来源有关.
长耳鸮; 黑线姬鼠; 越冬期; 食物组成
长耳鸮,隶属于鸟纲、鸮形目、鸱鸮科,主要分布于欧亚大陆及北美洲[1],共分化为指名亚种、北美东部亚种、加那利群岛亚种及北美西部亚种这四个亚种,我国仅分布有指名亚种[2],被列为国家二级重点保护野生动物.生活在我国境内的长耳鸮除青海、西宁、新疆喀什和天山等少数地区为留鸟外,其他大部分地区均为候鸟[3].本文研究地点位于上海崇明和江苏盐城,长耳鸮在两地均属冬候鸟.
国内对于长耳鸮食性的研究地点多分布于我国北部或西部地区,研究表明它们的食物主要是鼠类、蝙蝠以及一些鸟类、鱼类和昆虫[4-7],而我国南方鲜见相关报道.近年来,本文研究人员陆续在上海崇明和江苏盐城(地理位置归属南方)发现长耳鸮越冬种群,种群数量分别可达16只、29只,其中崇明连续观测两年后越冬种群消失.本文对两地越冬种群开展三年野外调查,通过收集长耳鸮完整食团、栖息地生境资料等,分析两地长耳鸮越冬食物组成,进而为越冬种群保护提供科学依据.
1 研究区域
崇明岛位于上海市(SH)区东北方向,2010年初在岛中北部东平国家森林公园附近的宝岛度假村和怡沁园度假村(31°40′32.81″~31°40′38″N,121°28′28.38″~121°28′40.60″E)发现长耳鸮越冬种群,并对2010—2012年两个越冬期进行研究.长耳鸮栖息地周围生境主要有农田、居民区、人工林地等.其中,农田生境是崇明岛面积最大的生境类型,而人工林主要包括香樟(Cinnamomumcamphora)、白榆(Ulmuspumila)、水杉(Metasequoiaglyptostroboides)、柳杉(Cryptomeriafortunei)等.长耳鸮主要栖息在柳杉和水杉等高大乔木上,植被高度10~25m,郁闭度60%~75%,冬季水杉叶落后郁闭度降为40%左右.栖息点0~5m内植被郁闭度普遍较低为2%~40%或无植被覆盖.两个度假村相邻,交界处有一条大约4m的河道.
2014年初,在江苏盐城林场(JS)发现长耳鸮越冬种群(33°36′24.77″N,120°28′17.29″E),林场以人工林地为主,周围生境主要为居民区、林地和农田.林场及周围林地主要植被是杜仲(Eucommiaulmoides)、栾树(Koelreuteriapaniculata)、池杉(Taxodiumascendens)、铅笔柏(Sabinavirginiana)、柳杉等.长耳鸮主要栖息在铅笔柏上,植被高度8~12m,郁闭度约50%.栖息点0~10m内主要植被为培植树苗或杜仲、枫杨等落叶乔木,冬季郁闭度较低.距长耳鸮栖息的铅笔柏20m处有一条河道.
2 方 法
2.1 调查方法
长耳鸮每年11月到达,次年3月左右离开,越冬期内每月上旬到其栖息的树下收集食团1次,收集时间4:30am~6:30pm.用双筒望远镜(Olympus,8LYM)观测长耳鸮,记录其飞出及返回时间;用BK8731型号照度计对光照强度进行测量,同时记录研究地点周围鸟类种类及数量.
2.2 分析方法
2.2.1 食团处理
将采集后的食团晾干,至60℃恒温箱内烘干至连续两次重量无明显变化.
2.2.2 数据测量及分析
挑选烘干后的完整食团,用上海佑科FA2004B电子天平(最小精确值0.1mg)称量干重,游标卡尺(0.01mm)测量长、短径;SPSS18.0软件分析上述数据差异.采用食团内容物分析法[8]分析长耳鸮食性,兽类以完整头骨为鉴定依据,依据其特征鉴定物种[9-10].鼠类年龄鉴定以上颌牙齿生长及磨损情况为依据[11],幼体臼齿齿突彼此独立,M3未长出,或未与M1、M2等高;亚成体M1、M2因磨损出现横嵴和纵桥,但两者2、3横嵴没有联结成环,M1前具两横列突头,M2中列、M3后列的各突头都分别连在一起,M1、M2后列突头部分尚未完全连结;成体Ⅰ组臼齿咀嚼面约有40%暴露,M1、M2中后列突头出现纵向连接形若马蹄;成体Ⅱ组咀嚼面已暴露出80%左右,M1、M2中后列突头形成封闭环,M3突头已融合;老年组臼齿面已磨平,磨面接近齿根.鸟类鉴定依据头骨和喙的特征,以一对上下喙记为一只.
黑线姬鼠的被捕食强度以本物种的个体数占鼠类总个体数的比例来表示;以含有鼠类头骨食团数占总食团数的比例表示该物种出现率[6].
3 结 果
3.1 食物组成及黑线姬鼠被捕食强度
两地共采集完整食团717份,共分离出816个头骨.江苏盐城采集完整食团140份,平均干重范围为1.11~5.56g.鉴定出头骨148个,其中黑线姬鼠94个、黄胸鼠(Rattusflavipectus)8个、[树]麻雀(Passermontanus)17个、山东小麝鼩(Crocidurasuaveolens)19个以及无法鉴别头骨10个.上海崇明岛两个越冬期共采集完整食团577份,平均干重范围为0.26~6.00g.鉴定出头骨共668个,其中黑线姬鼠623个、
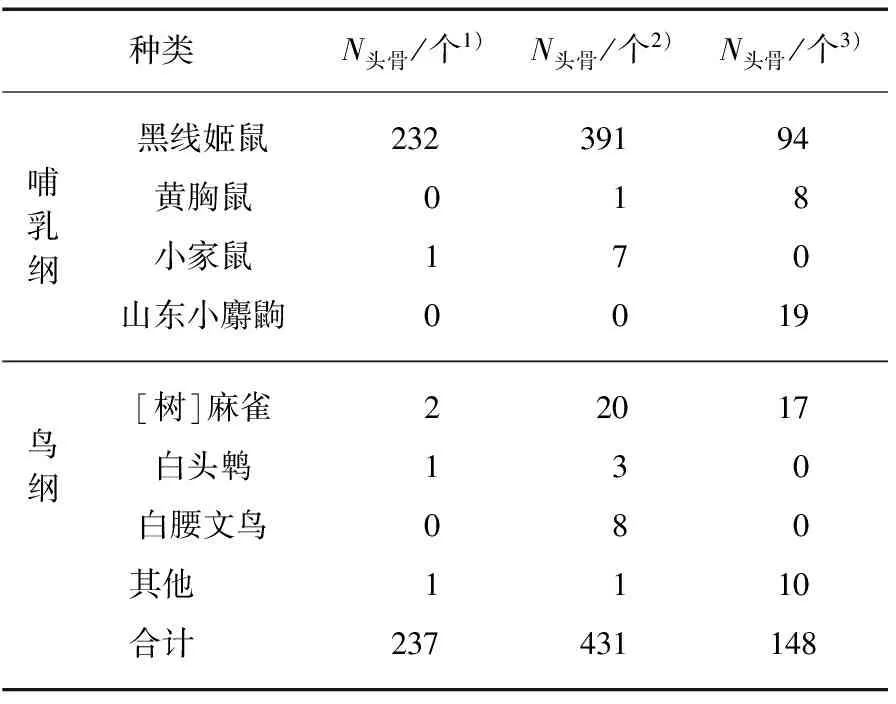
表1 长耳鸮越冬期食物组成Tab.1 Food composition of Long-eared owls in wintering period
注: 1) 上海2010—2011(n=181);2) 上海2011—2012(n=396);3) 江苏2014—2015(n=140).
黄胸鼠1个、小家鼠(Musmusculus)8个,分别占总头骨数的93.3%、0.2%、1.2%;[树]麻雀22个、白头鹎(Pycnonotussinensis)4个、白腰文鸟(Lonchurastriata)8个,及无法鉴别种类头骨2个(表1).
2011年2月采集食团的平均干重与11月差异极显著(P=0.007lt;0.05),与12月差异显著(P=0.034lt;0.05);2010年11月采集食团的长径与1月(P=0.004lt;0.05)、2月(P=0.005lt;0.05)差异极显著,12月与1月(P=0.014lt;0.05)、2月(P=0.015lt;0.05)差异显著.
单个食团解剖结果显示,2011—2012年越冬期没有头骨的食团数所占比例最高,含有1或2个头骨的食团数相等,均接近30%.2014—2015年含有1个动物头骨的食团数占该年越冬期食团总数最多,多达43.6%(表2),本越冬期含3或4个头骨的食团所占比例较2011—2012年有所降低.
2010—2012年,在上海崇明岛共采集完整食团577份,392份中有动物头骨,其中鼠类的出现率为66%,长耳鸮在其中5个月内对黑线姬鼠的捕食强度为100%(图1).2014—2015年江苏盐城采集完整食团140份,100份中有动物头骨,鼠类出现率58.6%,其中2015年1月黑线姬鼠的被捕食强度为100%,其他3个月相差不大,分别为90%、90%、91.1%(图1).
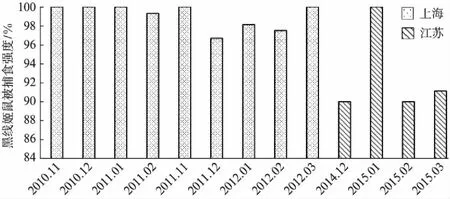
图1 长耳鸮对黑线姬鼠的捕食强度Fig.1 Predation rates on Apodemus agrarius by Long-eared owls
表2单个食团中头骨数量
Tab.2 The number of skulls in one pellet
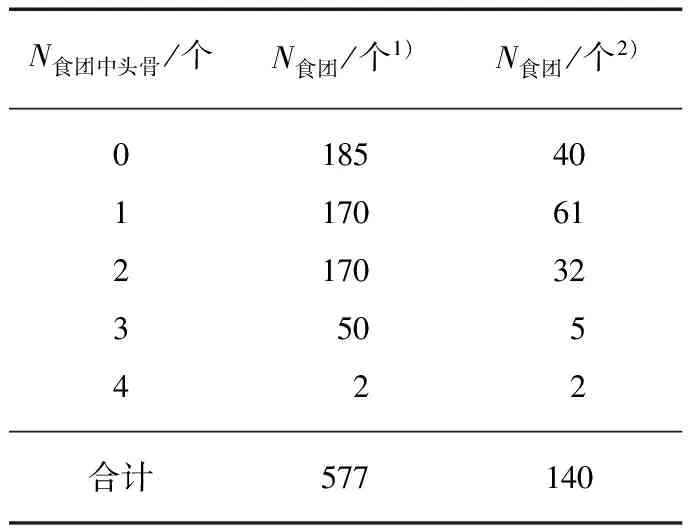
N食团中头骨/个N食团/个1)N食团/个2)0185401170612170323505422合计577140
1) 上海2011—2012;2) 江苏2014—2015.

图2 长耳鸮越冬期食物中黑线姬鼠的年龄组成Fig.2 Age composition of Apodemus agrarius in pellets of Long-eared Owls
3.2 黑线姬鼠的年龄组成
黑线姬鼠是长耳鸮在上海、江苏两地越冬的主要食物来源,分别占各自食物总数的93.3%,63.5%.上海食团中分离出的黑线姬鼠623个头骨,其中476个可鉴定出年龄,而江苏食团中分离的94个黑线姬鼠头骨均可鉴定出年龄.分析黑线姬鼠的年龄组成,黑线姬鼠成体Ⅰ在食物组成中所占比例最高,分别为41.1%、36.2%(图2),被捕食黑线姬鼠各年龄组在两个地点上无显著性差异,但成体Ⅰ组在11月差异性显著(P=0.026lt;0.05),成体Ⅱ组个体的数量在3月差异性显著(P=0.03lt;0.05),幼年组因数量过少未进行检验.3个越冬期内,长耳鸮对黑线姬鼠不同年龄组的捕食强度基本一致,且均对成体Ⅰ的捕食量最大.
3.3 对黑线姬鼠不同年龄组捕食随时间变化
上海越冬长耳鸮对黑线姬鼠不同年龄组的捕食随时间变化呈现一定的规律.2个越冬期内,长耳鸮对黑线姬鼠亚成体组的捕食百分比随时间的变化,呈现先增长后降低的趋势,分别由8%上升至20%、22.6%,然后降至11.6%,由25%上升至29.9%,然后降至22.2%、12.2%;对成体两个组的捕食总量呈现先减少后增加的趋势,如成年Ⅰ组分别由36%降至20%,然后上升到41.9%、50%,由31.2%降至29.9%后又上升到38.9%、43.8%;对老年组的捕食变化规律不明显(图3,见第520页).且仅在2011年11月对幼体组有捕食,数量为1只(图3(b)).
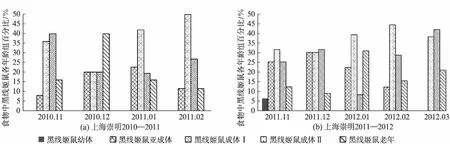
图3 黑线姬鼠年龄组成百分比Fig.3 The age composition and percentage of Apodemus agrarius
江苏越冬长耳鸮对亚成体组和成体Ⅰ、Ⅱ组的捕食量百分比变化趋势相反,对亚成体组捕食量减少时,对成体组捕食量增加,如亚成体由12月的55.6%降至12.5%然后上升33.3%时,成体Ⅱ组的捕食量由5.6%升至25%后又下降为11.1%;对老年组的捕食量百分比呈上升趋势(图4,见第520页).
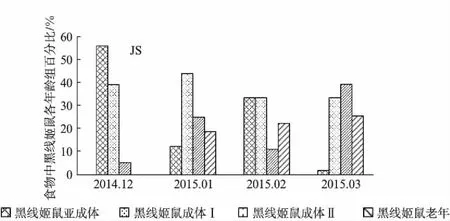
图4 2014—2015年黑线姬鼠年龄组成百分比Fig.4 The age composition and percentage of Apodemus agrarius from 2014 to 2015
4 讨 论
4.1 长耳鸮的主要食物组成
相对于同一地区生存的其他鸮类,长耳鸮的取食有一定的限定[12].一直以来,在北美洲、欧洲及亚洲等长耳鸮分布区对其食性的研究结果表明,长耳鸮主要捕食鼠类、昆虫和鸟类[12-15],近年来,北京等地研究发现,蝙蝠也是长耳鸮的主要捕食对象[5-6],而蝙蝠和鼠类所占比例较为相近[16].
本文两个研究地点中,长耳鸮的食物组成均主要是黑线姬鼠(表1),各月份中黑线姬鼠的被捕食强度均较高(图1).江苏省啮齿类调查结果中,以小家鼠、黑线姬鼠、黄胸鼠、及褐家鼠等4种分布最广,其中黑线姬鼠主要分布在野外,野外采集捕获率17.3%,是野外采集鼠类捕获率最高的物种;根据调查结果,淮河一线以南以黄胸鼠占50%以上.黄胸鼠在室内的捕获率最高,为21.7%,但野外也有采集,捕获率为0.5%[17].在江苏越冬的长耳鸮,黑线姬鼠是其食物的主要组成,这与当地黑线姬鼠是农田中优势种[18-20]研究结果一致.上海市室外鼠类以黑线姬鼠为主,为98.7%,室内黄胸鼠、小家鼠所占比例较高,分别为78.1%、16.1%[21].1958—1959年上海市郊越冬期野外农田捕获小鼠中,黑线姬鼠263只,占96%[22].本研究中,上海越冬长耳鸮的食物中黑线姬鼠所占比例为93.3%,偶尔也捕食黄胸鼠和小家鼠.上海奉贤区1988—2013年鼠类种群数量变动研究中结合气候、生态因子等探讨鼠类种群动态,结果显示黑线姬鼠是优势种,年代间变动不大,且将相关数据对照上海鼠类生态研究调查结果,显示近60年来上海野外农田鼠类种群构成比及优势种没有发生明显变化[23].
综合上述研究结果,表明长耳鸮的食物组成与当地食物资源多样性和可获得性相关[14].
分析长耳鸮单个食团内动物头骨数量可知,每个越冬期单个食团含有0~2个头骨的食团比例较高(表2),这在某种程度上反映了长耳鸮前一天的进食量.若要得到越冬单只长耳鸮进食量,需连续观测单只食团吐出情况.
4.2 食物中黑线姬鼠年龄组成及随时间变化情况
上海、江苏越冬长耳鸮主要捕食黑线姬鼠种群中的成体Ⅰ组、Ⅱ组,分别占黑线姬鼠个体总量的69.4%、63.8%(图2).以上海为例,长耳鸮食物中成体组占总量69.4%,这与上海郊区黑线姬鼠10月份到次年2月份种群中成体Ⅰ组、Ⅱ组占到70%~90%基本一致[11].黑线姬鼠的繁殖高峰期为每年9、10月份,幼体日龄在30d,亚成体的日龄在30~90d[11],所以12月前后,长耳鸮对黑线姬鼠亚成体组有较高的捕食量,之后随着亚成体生长为成体组后捕食量降低,而后对成体组的捕食量逐渐增加.食团采集时间一般在每年11月,此时种群中幼体基本生长为亚成体,所以仅在2011年11月间对黑线姬鼠幼年组有捕食记录.而江苏长耳鸮对亚成体组捕食量减少时,对成体组捕食量相应增加(图5),2月亚成体量偏高,可能与当月食团采集时间较早有关.长耳鸮对黑线姬鼠各年龄组的捕食量变化与当地种群中各年龄组的动态变化基本相符,进而推测长耳鸮对栖息地黑线姬鼠的捕食无年龄选择,为随机捕食.
上海和江苏越冬长耳鸮栖息地均以农田生境为主,农田生境为其提供了丰富的食物来源,栖息地周围郁闭度较高的柳杉、水杉及铅笔柏也提供了很好的遮蔽作用.栖息地为鸟类的生存提供充足的食物资源、适宜的繁殖地点及生存的保护条件,因而鸟类对栖息地具有一定的选择性[24].因此,越冬长耳鸮食物分析结果能为该物种的保护提供资料和依据.
致谢: 盛和林教授指导小型哺乳动物物种及年龄鉴定,刘楠、张航、董美麟、朱文杰、彭博对野外调查工作给予帮助,在此一并感谢.
[1] KÖNIG C, WEICK F. Owls of the world [M]. 2 ed. New Haven and London: Yale University Press, 2008: 479-481.
[2] 李湘涛.中国猛禽 [M].北京: 中国林业出版社,2004: 163-164.
[3] 张健旭.长耳鸮的迁徙与数量 [J].大自然,1996(6): 30.
[4] 曹玉萍.保定城区越冬长耳鸮的食性研究 [J].河北大学学报(自然科学版),1990(1): 63-68.
[5] 李晓京,鲍伟东,孙来胜.北京市区越冬长耳鸮的食性分析 [J].动物学杂志,2007,42(2): 52-55.
[6] 袁梨,鲍伟东,王安梦,等.北京市越冬长耳鸮食物中的蝙蝠成分 [J].林业科学,2010,46(5): 109-112.
[7] 汪洋,曾亚琦,高一峰,等.乌鲁木齐市区越冬期长耳鸮的食性分析 [J].动物学杂志,2012,47(4): 36-40.
[8] LOVARI S, RENZONI A, FONDI R. The predatory habits of the barn owl(Tytoalba, scopoli) in relation to the vegetation cover [J].BollZool, 1976,43: 173-191.
[9] 韩崇选,李金钢,杨学军,等.中国农林啮齿动物与科学管理 [M].西安: 西北农林科技大学出版社,2005.
[10] 潘清华,王应祥,岩崑,等.中国哺乳动物彩色图鉴 [M].1版.北京: 中国林业出版社,2007: 316-333.
[11] 祝龙彪,钱国桢.黑线姬鼠种群的年龄结构及种群更新的研究 [J].兽类学报,1982,2(2): 211-217.
[12] MARTI C D. A review of prey selection by long-eared owl [J].Condor, 1976,78(3): 331-336.
[13] 曹玉萍,张健旭.河北保定市长耳鸮越冬种群及其捕鼠效应 [J].四川动物,1995,14(3): 134-135.
[14] BERTOLINO S, GHIBERTI E, PREEONE A. Feeding ecology of long-eared owl(Asiootus) in northern Italy: Is it a dietary specialist [J].CanadianJournalofZoology, 2001,79(12): 2192-2198.
[15] TIAN L, ZHOU X W, SHI Y,etal. Bats as the main prey of wintering long-eared owl(Asiootus) in Beijing: Integrating biodiversity protection and urban management [J].IntegrativeZoology, 2015,10: 216-226.
[16] 张逦嘉,王安梦,鲍伟东,等.不同栖居地和越冬时期长耳鸮的食物组成 [J].生态学杂志,2009,28(8): 1664-1667.
[17] 周开亚,钱伟娟,杨光平,等.江苏省啮齿类的调查 [J].动物学杂志,1981(3): 38-42.
[18] 李世良,刘树法.田间鼠害动态分析 [J].江苏农业科学,1989(1): 27-28.
[19] 沈兆昌,阮治安,颜正生,等.江苏农田黑线姬鼠的生态特点与防治对策 [C]∥中国有害生物综合治理论文集.北京: 中国农业科技出版社,1996: 1030.
[20] 王加连.江苏盐城自然保护区陆生兽类资源调查研究 [J].四川动物,2009,28(1): 140-144.
[21] 钱国桢,祝龙彪.上海鼠类生态的研究 [J].动物学研究,1983,4(4): 339-344.
[22] 盛和林,施银柱,马世全,等.上海市郊黑线姬鼠的越冬地点及其消灭方法 [J].动物学杂志,1959(5): 189-192.
[23] 费蕾,汪祖国,姚瑶,等.上海奉贤区农田鼠类种群数量变动以及气候和耕作因素的影响 [J].应用生态学报,2015,26(2): 579-587.
[24] 蒋爱伍,周放,覃玥,等.中国大陆鸟类栖息地选择研究十年 [J].生态学报,2012,32(48): 5918-5923.
AnalysisofFoodCompositionofWinteringLong-earedOwl(Asiootus)—ACaseStudyBasedonTwoLocalitiesofShanghaiandJiangsuProvinceinChina
WANGJunfu1,LIBicheng1,BOShunqi2,HEXin1,YANGGang1,YUANXiao2,TANGSixian3
(1.NaturalHistoryResearchCenter,ShanghaiScienceamp;TechnologyMuseum,Shanghai200127,China;2.ShanghaiWildlifeConservationandManagementCenter,Shanghai200023,China;3.SchoolofLifeSciences,EastChinaNormalUniversity,Shanghai200062,China)
During 2010—2012 and 2014—2015, pellets of wintering Long-eared owls in Chongming county(Shanghai) and Yancheng City(Jiangsu Province) were collected and the composition of the food were analyzed. Totally, 717 intact pellets were collected which include bones of 804 individuals from 7 species, 4 species were small mammals, such as rodents and musk shrews, accounting for 93.7% of the total pellet samples, and the other 3 species were small birds of Passeriformes. The results showed that the amount of the predation onApodemusagrariuswas the largest, accounting for 89.2%. And, the predation on adult groupⅠof theApodemusagrariuswas taken by the highest percent, accounting for 40.2% in numbers. Moreover, the proportion of predation amount of different age group is the same as that of native population. We supposed that the predation targets of wintering Long-eared Owl changed according to the surroundings of local prey and the compositon.
Long-eared owl;Apodemusagrarius; wintering period; food composition
0427-7104(2017)04-0517-06
2016-11-04
王军馥(1984—),女,硕士研究生;唐思贤,高级工程师,通信联系人,E-mail: sxtang@bio.ecnu.edu.cn.
Q943.2
A
猜你喜欢
林业科技(2021年2期)2021-07-08 05:02:50
湖北农业科学(2021年2期)2021-02-07 03:32:40
少年文艺·我爱写作文(2020年11期)2020-11-23 01:52:52
湖北农业科学(2019年20期)2019-12-11 10:03:58
贵州农业科学(2019年7期)2019-08-03 09:46:30
湖北农业科学(2019年23期)2019-01-11 17:38:36
野生动物学报(2018年4期)2018-11-23 03:04:20
饮食与健康·下旬刊(2017年12期)2017-12-06 17:53:41
吉林大学学报(医学版)(2015年1期)2015-12-17 07:47:24
初中生之友·中旬刊(2015年10期)2015-11-03 00:08:50
