
杉木天冬酰胺合成酶基因的克隆与生物信息学分析
2017-12-09 07:37薛爽董毅饶丽莎许珊珊林思祖
安徽农学通报 2017年22期
薛爽+董毅+饶丽莎+许珊珊+林思祖
摘 要:为了解杉木天冬酰胺合成酶基因(ASN)的相关信息,以杉木组培苗105为材料,克隆获得杉木ASN3基因,该基因为一个完整的开放阅读框,共编码231个氨基酸,生物信息学分析显示:ASN3基因编码蛋白为不含卷曲螺旋结构和信号肽的非跨膜稳定亲水蛋白,定位于微体(过氧化物酶体),具有多样的磷酸化位点,其中丝氨酸15个,苏氨酸7个,酪氨酸3个,其二级及三级结构主要由无规则卷曲和α-螺旋组成。
关键词:杉木;天冬酰胺合成酶基因;克隆;生物信息学
中图分类号 S791.27 文献标识码 A 文章编号 1007-7731(2017)22-0028-03
Abstract:In order to understand the relevant information of ASN gene of Cunninghamia lanceolata,ASN3 gene which was a complete open reading frame with encoding 231 amino acids was cloned from tissue culture seedling 105 of Cunninghamia lanceolata.According to bioinformatics analysis, protein encoded by ASN3 gene was stable and closed to water, without the signal peptide, winded helix and transmembrane domain, located in microbody(peroxisome) and had a variety of phosphorylation sites, with 15 serine, 7 threonine and 3 tyrosine.The secondary structures and three-dimensional structure of ASN3 gene were mainly composed of random coil and alpha helix.
Key words:Cunninghamia lanceolata;ASN gene;cloning;Bioinformatics analysis
天冬酰胺合成酶是广泛存在于植物体内的一类由小基因家族编码的氨基转移酶,主要以氨或谷氨酰胺及天冬氨酸为底物催化合成天冬酰胺。根据其底物的不同,天冬酰胺合成酶可分为AS-A和AS-B,前者为氨依赖性,仅存于原核生物;后者为谷氨酰胺依赖性,广泛存在于原核和真核生物[1]。因此,普遍认为植物天冬酰胺主要由AS-B催化合成。目前,已从大豆[2-3]、小黑杨[4]、玉米[5]、海岸松[6-8]、桑树[9]、小黑麦[10]、向日葵[11]等多种植物中成功分离ASN基因,且进一步的功能研究发现ASN基因具有一定的抗逆功能。杉木作为我国南方重要的造林树种,已成功分离出一个ASN1基因[12],本研究将在此基础上进一步分离杉木ASN基因其他新成员,以期为后续系统研究ASN基因在杉木生长发育及逆境过程中的调控作用奠定基础。
1 材料与方法
1.1 材料 供试材料为国家林业局杉木工程技术研究中心保存的杉木组培苗105。
1.2 方法
1.2.1 RNA提取和cDNA合成 参考官清娜[12]方法进行杉木总RNA提取,经酶标仪和琼脂糖凝胶(1%)检测合格的RNA用于cDNA合成。根据TIANScript cDNA第一链合成试剂盒(TIANGEN)说明书反转录cDNA。
1.2.2 引物設计与PCR扩增 将已扩杉木ASN1基因序列与杉木转录组中预测的ASN基因序列比对,设计上游引物5-ATGAAAGCTTTGCACGATGACTG-3和下游引物5-TTAACCCTGAATGACAACTCCCCT-3用于PCR扩增。扩增反应程序:94℃预变性3min;94℃变性30s,60℃退火30s,72℃延伸2min,35个循环;72℃延伸7min。将PCR扩增产物送北京六合华大基因生物技术公司测序。
1.2.3 生物信息学分析 利用在线软件对目的基因所编码蛋白的理化性质(ExPASy)、信号肽(SignaIP 4.1 Server)、跨膜结构(ExPASy)、亚细胞定位(PSORT)、磷酸化位点(NetPhos 2.0 Sever)、卷曲螺旋结构(EMBnet Coils)、二级结构(GOR IV)和三级结构(SWISSMODEL)进行预测和分析。
2 结果与分析
2.1 杉木ASN基因新成员的cDNA序列的获得 根据所设引物进行PCR扩增,经电泳后发现有两条条带,分别是1300bp和700bp,经PCR产物直接测序后,只得到一条696bp的序列,经DNAMAN软件翻译后,发现该转录本含有起始密码子ATG和终止密码子TAA,可完整编码,为一个完整的编码框,共编码231个氨基酸。将该转录本的核苷酸序列及其翻译的氨基酸序列在NCBI上进行在线比对,显示该转录本与海岸松(HQ625490.1)、樟子松(AJ496567.2)及落叶松(KF515226.1)等已登录物种的ASN基因高度同源,表明该转录本为杉木ASN基因新成员,命名为ASN3,其所编码的氨基酸序列如下:
MKALHDDCLRANKSTSAWGLEARVPFLDKEFINVAMDIDPDWKMVRPDQGRIEKWILRKAFDDKENPYLPQHILYRQKEQFSDGVGYSWIDGLKAHAAEHVSDMMLMNAKYVFPHNTPSTKEAFYYRMIFERFFPQNAARLTVPGGPSVACSTAKAVEWDAAWSNHLDPSGRAALGVHASAYKDKDKLSSSSSHSNLPLLTVTAGLGTSLDKSPGPLMGDPLTARGendprint
VVIQG.
2.2 杉木ASN3基因的生物信息学分析 蛋白质是生命的物质基础,其氨基酸序列、空间结构、修饰基团等决定了其相应的蛋白功能。因此,利用蛋白质分析软件对ASN3基因编码蛋白质的理化性质、蛋白结构、翻译后磷酸化修饰进行分析,有助于推断其功能和作用机理,为后续的功能分析和作用机理研究提供依据。
2.2.1 ASN3蛋白理化性质分析 利用在线软件ExPASy对ASN3蛋白进行理化性质分析,结果显示,该蛋白分子式为C1148H1770N316O335S10,相对分子质量为25679.20Da,由20种氨基酸组成,其中丙氨酸所占比例最大,为10.4%;该蛋白带27个正电荷(Asp+Glu)和27个负电荷(Arg+Lys),pI等电点为7.09;不稳定系数和亲水性分别为34.79和-0.397,为稳定的亲水蛋白。进一步利用ExPASy和SignaIP 4.1软件对ASN3蛋白的跨膜结构及信号肽进行分析,结果显示,ASN3蛋白不具有跨膜结构和信号肽,属于非分泌蛋白。亚细胞定位(PSORT)分析显示ASN3蛋白定位于微体(过氧化物酶体)的可能性最大,分值为0.657。
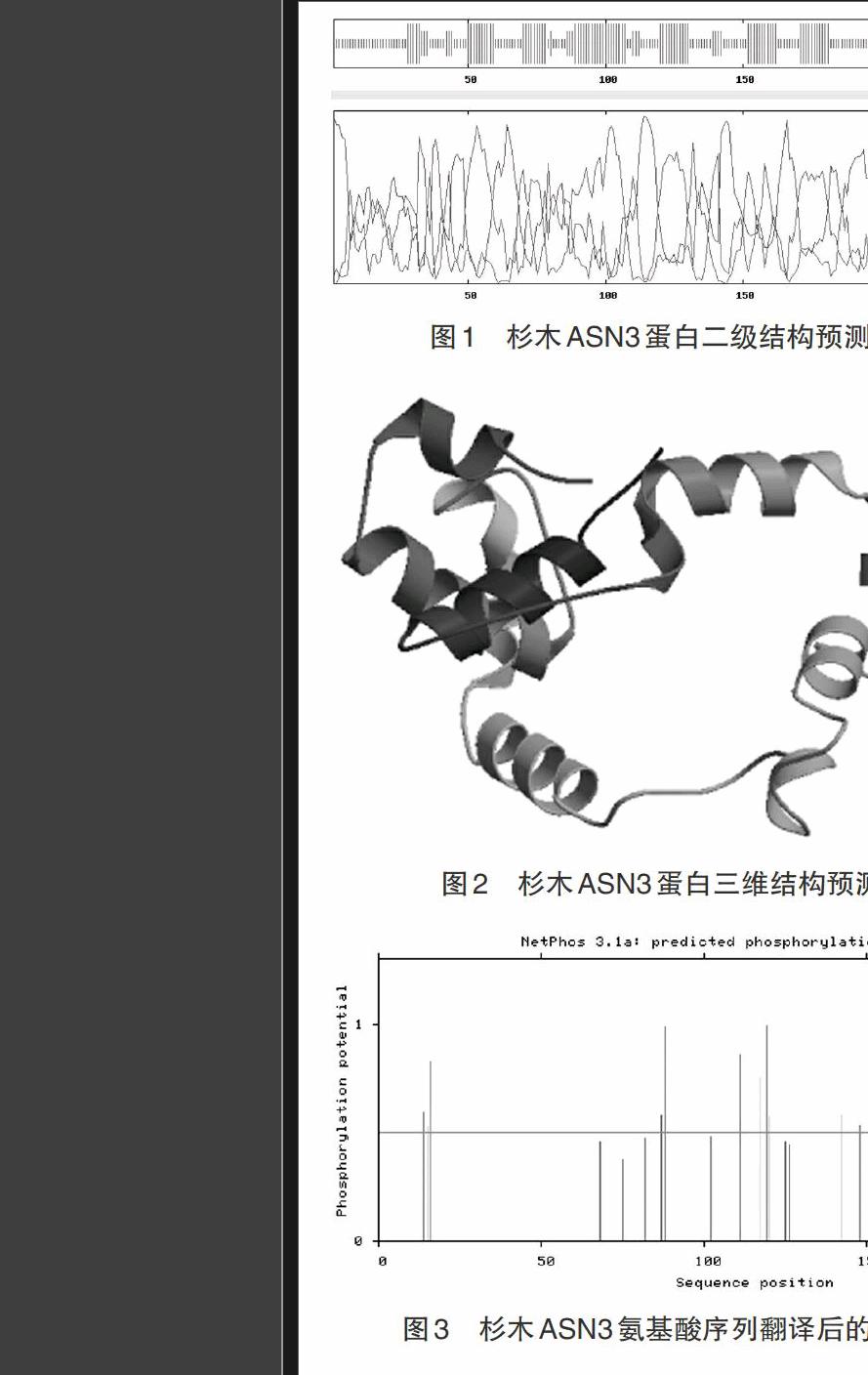
2.2.2 ASN3蛋白结构特征分析 利用EMBnet Coils软件对ASN3蛋白质卷曲螺旋进行预测,结果表明,以window=14、21和28为参数,ASN3蛋白在整个肽链上不存在卷曲螺旋结构。二级结构分析显示该蛋白含有55.41%的无规则卷曲、32.9%的α-螺旋和11.69%的延伸链(图1)。三级结构分析结果(图2)和二级结构分析结果相似,表明该蛋白主要由无规则卷曲和α-螺旋组成。
2.2.3 翻译后磷酸化修饰分析 利用NetPhos 2.0 Sever对ASN3蛋白潜在的磷酸化位点进行分析,结果显示该蛋白共有25个磷酸化位点,其中丝氨酸磷酸化位点15个,苏氨酸磷酸化位点7个,酪氨酸磷酸化位点3个(图3)。
3 结论与讨论
ASN参与植物体内氮从源组织到库组织的重新定位[13],是植物体氮代谢过程中的关键酶。本试验成功分离杉木ASN3基因,该基因为一个完整的编码框,共编码231个氨基酸,对其所编码的蛋白进行生物信息学分析,发现该蛋白为不含卷曲螺旋结构和信号肽的非跨膜稳定亲水蛋白,具有多样的磷酸化位点,其中丝氨酸15个,苏氨酸7个,酪氨酸3个,推测其可能以丝氨酸磷酸化为主和苏氨酸磷酸化为辅的方式影响杉木生长发育过程中ASN酶活性及分子构象,进而参与调控杉木的生长发育。
参考文献
[1]Gaufichon L, Reisdorf-Cren M, Rothstein S J, et al.Biological functions of asparagine synthetase in plants[J].Plant Science, 2010, 179(3): 141-153.
[2]张继, 王相晶, 于丹, 等.大豆天冬酰胺合成酶 β 基因的克隆及在大肠杆菌中的表达[J].东北农业大学学报, 2013, 44(7): 17-21.
[3]Wan T F, Shao G H, Shan X C, et al.Correlation between AS1 gene expression and seed protein contents in different soybean (Glycine max [L.]Merr.) cultivars[J].Plant Biology, 2006, 8(02): 271-276.
[4]郝丙青.小黑杨天冬酰胺合成酶基因AS功能的研究[D].哈尔滨:东北林业大学, 2016.
[5]Todd J, Screen S, Crowley J, et al.Identification and characterization of four distinct asparagine synthetase (AsnS) genes in maize (Zea mays L.)[J].Plant science, 2008, 175(6): 799-808.
[6]Ca?as R A, de la Torre F, Cánovas F M, et al.Coordination of PsAS1 and PsASPG expression controls timing of re-allocated N utilization in hypocotyls of pine seedlings[J].Planta, 2007, 225(5): 1205-1219.
[7]Canales J, Rueda-López M, Craven-Bartle B, et al.Novel insights into regulation of asparagine synthetase in conifers[J].Frontiers in plant science, 2012, 3.
[8]Canas R A, de la Torre F, Canovas F M, et al.High levels of asparagine synthetase in hypocotyls of pine seedlings suggest a role of the enzyme in re-allocation of seed-stored nitrogen[J].Planta, 2006, 224(1): 83-95.
[9]潘宝华, 潘刚, 方荣俊, 等.桑树天冬酰胺合成酶基因的克隆与表达分析[J].蚕业科学, 2011, 37(1): 1-8.
[10]张晓磊.小黑麦天冬酰胺合成酶基因克隆及功能验证[D].石河子:石河子大学, 2013.
[11]程维舜.植物天冬酰胺合成酶基因的抗病性调控功能分析[D].杭州:浙江大学, 2011.
[12]官清娜.杉木天冬酰胺合成酶基因的克隆与表达分析[D].福州:福建农林大学,2015.
[13]Lam H M, Wong P, Chan H K, et al.Overexpression of the ASN1 gene enhances nitrogen status in seeds of Arabidopsis[J].Plant physiology, 2003, 132(2): 926-935.
(責编:施婷婷)endprint
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
兽医导刊(2016年12期)2016-05-17
广西林业科学(2016年2期)2016-03-20
现代检验医学杂志(2015年4期)2015-02-06
