
各径级马尾松树干CO2释放速率及其温度敏感性
2017-12-05 05:15:22樊后保王勇刚李志鹏
林业科学 2017年10期
涂 洁 樊后保 王勇刚 李志鹏
(南昌工程学院生态与环境科学研究所 江西省退化生态系统修复与流域生态水文重点实验室 南昌 330099)
各径级马尾松树干CO2释放速率及其温度敏感性
涂 洁 樊后保 王勇刚 李志鹏
(南昌工程学院生态与环境科学研究所 江西省退化生态系统修复与流域生态水文重点实验室 南昌 330099)
【目的】 探明退化红壤区重要先锋树种马尾松不同径级间树干CO2释放速率(FCO2)的差异及其温度敏感性,以期为正确评价马尾松人工林树干呼吸对森林生态系统的碳汇贡献提供基础数据。【方法】 采用全自动便携式光合仪与自制PVC呼吸环相连,对马尾松人工林4个径级(15.9~17.9,18.9~20.5,22.8~24.8和27.1~29.8 cm)共12棵样树的树干CO2释放速率FCO2进行为期1年的活体原位监测,并同步监测树干1 cm深处温度,分析FCO2的季节变化及径级间差异,并建立FCO2与树干温度的指数函数模型。【结果】 4个径级FCO2均呈现出一致的单峰季节性变化趋势,生长季FCO2极显著高于非生长季(Plt;0.01),7月份最高,1月份最低,且生长季各径阶样树FCO2间差异显著(Plt;0.01); 4个径级FCO2与树干温度均存在显著的指数关系,但两者间的回归模型及其决定系数因径阶而异,表现出大径阶FCO2与树干温度的相关性高于小径阶; 4个径级树干呼吸的温度敏感系数(Q10)间差异不显著(1.47~1.64); 马尾松FCO2与DBH显著正线性相关(R2=0.89),表明DBH可作为针叶树种FCO2预测和推算的一个简便实用指标。【结论】 在采用局部通量法进行森林生态系统树干呼吸估算时,应同时考虑FCO2的季节变化及径级差异,以提高估算精度。
马尾松; 树干温度; 树干CO2释放速率; 径级
树干作为树木生物量的主体,一方面是森林生态系统长期稳定的碳汇(Lavigneetal., 1997),另一方面因树干内活细胞始终要维持呼吸而成为不容忽视的碳源(Edwardsetal., 2002)。树干呼吸占地上部分总自养呼吸的14%~48%(Voseetal., 2002),在一定条件下甚至超过叶呼吸量(Wieseretal., 2009)。近几十年来,随着原位测量技术的快速发展以及人们对陆地生态系统碳平衡过程的不断认识,树干呼吸逐渐成为森林生态系统碳循环研究的热点问题(Zhaetal., 2004; Teskeyetal., 2008; Arakietal., 2010)。目前,测定树干呼吸最普遍的方法是将局部测得的树干CO2释放速率(FCO2)作为树干呼吸的表征参数,再以林分调查常见的空间纯量如胸径作为转换因子,估测整株树木或林分尺度的呼吸通量。由于局部测得的FCO2往往存在显著的种间(Wangetal., 2008a)或种内差异(Ceschiaetal., 2002; Wangetal., 2003),并且FCO2与DBH间的回归关系也存在很大的不确定性(肖复明等, 2005; 石新立等, 2010)。温度是控制呼吸速率的重要因子(Amthor, 2000),FCO2与树干温度间存在显著的自然指数关系(Arakietal., 2010; 2014)。然而,树干CO2释放速率并没有严格遵循与温度的函数关系,晴朗天气测得的树干CO2释放速率比利用指数方程预测的值低25%~50%(Maieretal., 2006),在野外条件下,两者间的关系变得不明显(Etzoldetal., 2013)甚至不相关(Zhuetal., 2012)。因此,精细量化FCO2及其对温度的敏感性对于深入理解森林碳循环过程及其驱动机制、减少森林碳收支估测中的不确定性等都具有重要意义。
以往对树干呼吸的研究主要集中在温带(王淼等, 2005)和热带森林(严玉平等, 2009),对亚热带森林的报道不多(黄玮等, 2009),这将影响我国森林碳平衡模型的构建。江西千烟洲曾是亚热带的土壤严重退化区域,经过近三十年的小流域治理与生态恢复,目前该区植被恢复重建工作已取得巨大成效,研究者围绕重建生态系统的碳库开展了大量工作(马泽清等, 2007),而对先锋树种树干呼吸的报道尚不多见。本研究采用全自动便携式光合仪(LC PRO-SD,ADC Ltd.),对该区典型树种不同径级(5.9~17.9,18.9~20.5,22.8~24.8和27.1~29.8 cm)马尾松(Pinusmassoniana)FCO2进行为期1年的活体原位测定,旨在探明马尾松FCO2径级间差异及其对温度的敏感性,以期为提高森林生态系统树干呼吸的估算精度、正确评价马尾松人工林对我国亚热带退化生态系统的碳汇贡献提供基础数据。
1 研究区概况
研究地位于江西省泰和县中国科学院千烟洲试验站(115°04′1″26°E,26°44′48″N),海拔100 m左右,相对高度20~50 m,属典型红壤丘陵地貌。主要土壤类型包括红壤、水稻土、潮土和草甸土等,成土母质多为红色砂岩、砂砾岩或泥岩以及河流冲积物。区内年均气温17.9 ℃,≥0 ℃年积温6 523 ℃,年日照时数1 406 h,太阳总辐射量4 223 MJ·m-2,全年无霜期323天。年均降水量1 542 mm,其中4—6月约占全年的50%,7—8月高温少雨,年均空气相对湿度84%,属典型亚热带季风气候。
研究样地设在1986年营造的马尾松人工纯林内,海拔约70 m,坡度15°,坡向西偏南10°。根据2013年样地调查结果,马尾松林分密度约1 640株hm-2,活立木平均树高15.5 m,平均胸径18.1 cm。由于该林分郁闭度达95%以上,林下只分布了少量白栎(Quercusfabri)、山芝麻(Helicteresangustifolia)和白茅(Imperatacylindrica)等小灌木和草本类植物。
2 研究方法
2.1 树干CO2释放速率的测定 采用全自动便携式光合仪(LC PRO-SD)与土壤呼吸室原位活体监测树干CO2释放速率(FCO2)。选取生长正常、无缺陷、4个径级(15.9~17.9,18.9~20.5,22.8~24.8和27.1~29.8 cm)的马尾松各3株,共12株树。为排除太阳直射对测量结果的影响,在树干北面离地1.35 m处安装自制PVC呼吸环(高6.0 cm、外径11.4 cm、内径6.5 cm)。选择树干相对平坦部位打磨树皮使其接近平面,以防止安装呼吸环时漏气,但要避免伤及树干活组织。将PVC环较小一端稍作打磨,使其与树干表面吻合,用100%中性透明防水硅酮胶将其固定在树干上,并且保持其在整个测定期间位置不变; 将呼吸环较大一端与土壤呼吸室密封连接。安装时小心除去松脱的表皮和附生物(避免伤及树皮活组织),以免安装时漏气。2015年1—12月,每月下旬连续2个晴天的8: 00-17: 00时间段测定FCO2。每个呼吸环的采样时间约为15 min,12个呼吸环全部完成采样约需3 h,每个呼吸环1天内进行2轮FCO2测定(μmol·m-2s-1)。树干FCO2月均值为4个径级当月所有观测结果的算术平均。
2.2 树干温度的测定 在PVC环下方5 cm树干处钻深约1 cm的小孔(直径0.3 cm),在测定树干呼吸速率的同时,利用光合仪系统内置温度传感器同步测定树干1 cm 深处的温度(Ts)。
2.3 气室覆盖面积的测定 由于与树干接触端PVC环外径(6.5 cm)较小,且安装在树干较平坦部位,因此可以认为PVC环所覆盖的面积近似等于PVC环的底面积。
2.4 数据分析 树干CO2释放速率的所有观测数据由LC PRO-SD自动记录。考虑到该系统计算FCO2时采用的气室覆盖面积默认为97 cm2,因此需将仪器上该项参数调整为PVC环的实际覆盖面积33 cm2,实现对系统FCO2测定结果的校正。数据处理及统计分析由SPSS软件(SPSS 17.0 for windows, SPSS Inc., Chicago, IL, USA)完成,绘图利用KaleidaGraph软件(Synergy Software Inc., Reading, PA, USA)进行。树干CO2释放速率与温度间的回归关系采用自然指数函数拟合(公式1),同时按照公式(2)计算温度敏感系数Q10。不同径级和月份马尾松FCO2间的差异显著性采用单因素方差法分析,并进行LSD多重比较。
FCO2=β0eβ1Ts;
(1)
Q10=e10β1。
(2)
式中:β0为Ts为0 ℃时树干CO2释放速率(μmol·m-2s-1),β1为温度系数,Q10为树干温度每增加10 ℃呼吸速率增加的倍数。
3 结果与分析
3.1 不同径级马尾松FCO2的季节动态 对4个径级马尾松每月FCO2的观测结果进行算术平均可知,所有径级的FCO2均呈现出一致的单峰季节变化趋势(图1),这与马尾松的物候季节变化规律相吻合。3—7月随树木生长和气温升高,FCO2呈不断上升的趋势,7月达到全年最大值(2.10±0.22)~(3.26±0.68)μmol·m-2s-1,8~12月FCO2逐步递减,1,2月基本维持在较低水平(0.39±0.04)~(0.84±0.11)μmol·m-2s-1,1月达到全年最小值(0.39±0.04)~(0.69±0.12)μmol·m-2s-1,最大值约为最小值的8倍。在生长季,4个径级的FCO2间存在极显著差异(Plt;0.01),而在非生长季,除了(27.1~29.8)cm径级的FCO2极显著(Plt;0.01)高于其他3个径级,其余3个径级的FCO2间差异均不显著(Plt;0.05)。总体来看,在生长季马尾松(3—11月)的FCO2极显著(Plt;0.01)高于非生长季(1,2和12月)。
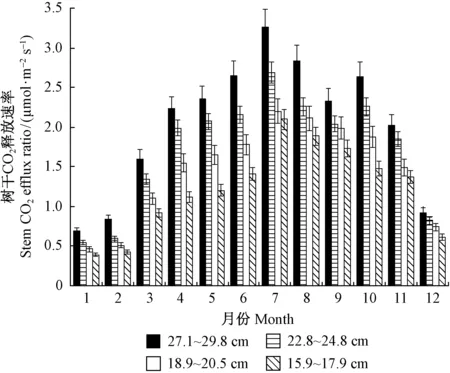
图1 4个径阶马尾松树干CO2释放速率的季节动态Fig.1 Seasonal dynamics of stem CO2 efflux ratio
3.2 不同径级马尾松FCO2与树干温度的关系 4个径级FCO2与树干温度的回归分析结果见表1和图2。4个径级FCO2与树干温度均存在极显著(Plt;0.01)或显著(Plt;0.05)的自然指数函数关系,表明树干温度能较好地解释FCO2的季节变化。从回归模型的决定系数R2大小看出,树干温度分别能反映4个径级(由大到小)马尾松FCO2变化的70.4%,65.9%,64.1%和52.8%,而且大径级FCO2与树干温度的指数相关性较小径级更高。4个径级树干呼吸速率的Q10值分别为1.64,1.55,1.47和1.60,Q10值没有明显的变化规律。
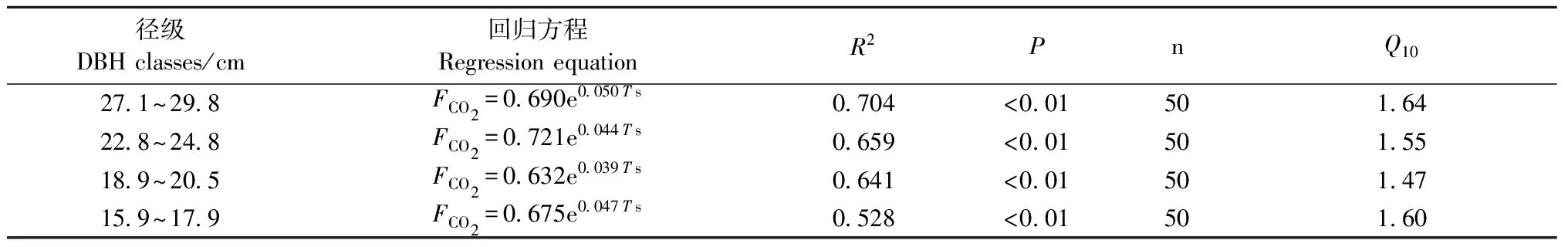
表1 马尾松树干CO2释放速率(FCO2)与树干温度的回归方程和Q10值Tab.1 Regression equations between stem CO2 efflux ratio and stem temperature and Q10 values
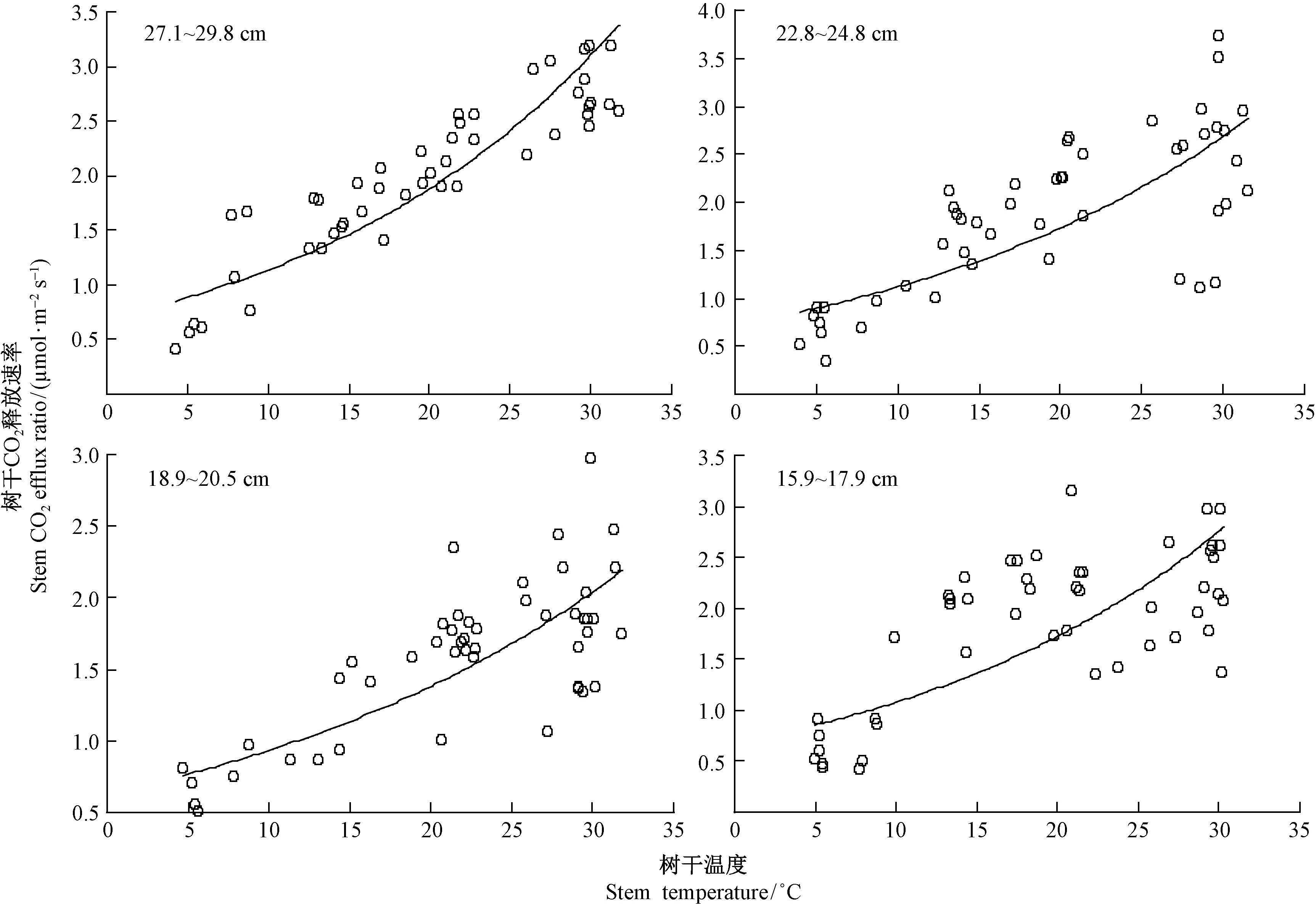
图2 不同径级马尾松树干CO2释放速率与树干温度的关系Fig.2 Relationships between stem CO2 efflux ratio and stem temperature of Pinus massoniana among different DBH classes
3.3 马尾松FCO2与胸径的相关关系 对马尾松12个月的FCO2观测数据统计分析发现,27.1~29.8 cm径级的FCO2年均值最大(2.39 μmol·m-2s-1),其他依次为22.8~24.8(2.02 μmol·m-2s-1),18.9~20.5(1.53 μmol·m-2s-1)和15.9~17.9 cm(1.16 μmol·m-2s-1),表明FCO2随径级的增大而增大。27.1~29.8与22.8~24.8 cm径级的FCO2间差异显著(Plt;0.05),且与其他径级的FCO2间也存在显著性差异(Plt;0.05),但18.9~20.5与15.9~17.9 cm径级的FCO2间差异不显著(Plt;0.05)。对4个径级的FCO2与胸径进行回归分析表明,FCO2与胸径存在极显著的正线性相关关系(Plt;0.01),胸径可以解释FCO2年际变化的89%。
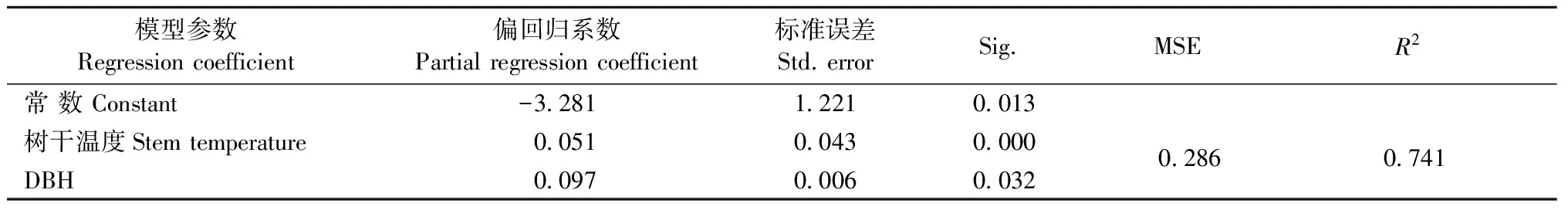
为了综合考虑树干温度和胸径对树干CO2释放速率的影响,参照Yang等(2012)的方法,利用Ln(FCO2)=a+bTs+cDBH建立FCO2与树干温度、胸径的二元线性回归模型(表2)。回归分析结果显示,树干CO2释放速率可由模型Ln(FCO2)=-3.281+0.051Ts+0.097DBH很好地拟合,回归方程和回归系数的相关性检验均达显著性水平(Plt;0.05)。对比表1和2中回归模型的决定系数R2大小可看出,树干温度和胸径可以共同解释FCO2的74.1%,比单独用树干温度因子建立的自然指数模型有所提高。
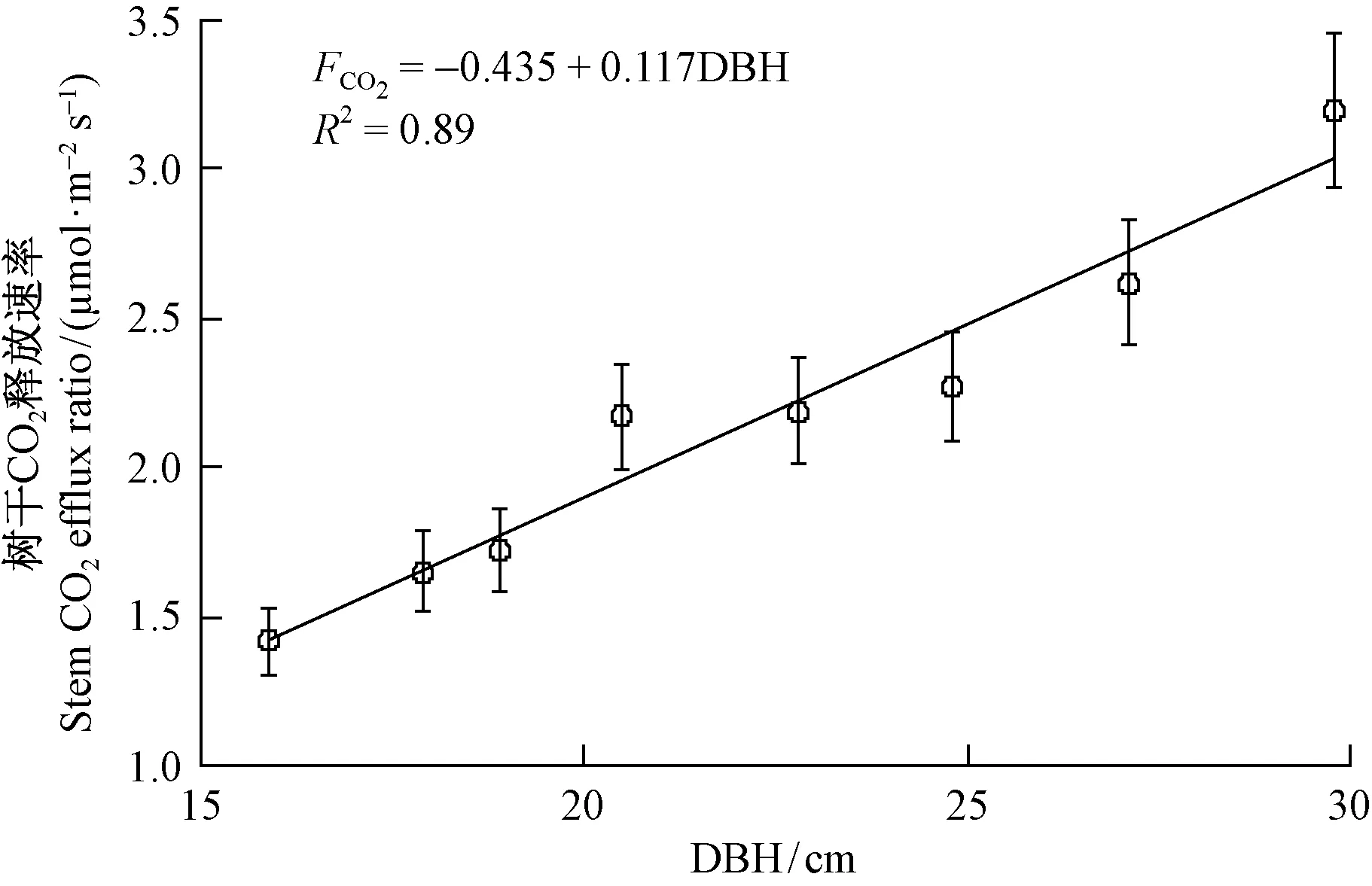
图3 马尾松树干CO2释放速率与胸径的关系Fig.3 Relationship between stem CO2 efflux ratio and DBH of Pinus massoniana
模型参数Regressioncoefficient偏回归系数Partialregressioncoefficient标准误差Std.errorSig.MSER2常数Constant-328112210013树干温度Stemtemperature005100430000DBH00970006003202860741
4 讨论
马尾松4个径级的FCO2呈一致的单峰季节变化趋势,且生长季FCO2显著高于非生长季,这与大多数文献的结论相同(严玉平等, 2009; Britoetal., 2010; Zhuetal., 2012)。研究区7月为全年高温时期,加之前期储备的大量土壤水分,水热同步的优越环境有利于树木木质部生长,从而促进了树木生长呼吸代谢(Lavigneetal., 2004),叠加全年都具有的维持呼吸,总体表现为这一时期树干呼吸速率最大。进入冬季后,大气温度逐渐降低,马尾松生长速度开始减缓,至1月份温度降至最低,严重抑制了树干的碳代谢,树木生长几乎停滞,自养呼吸则以较低的维持呼吸为主,因此树干呼吸速率最小。
马尾松4个径级的FCO2与树干温度间存在极显著或显著的自然指数关系,与大多数研究结论相同(Arakietal., 2014; Etzoldetal., 2013)。而Zach 等(2010)对热带山区森林的研究表明,树干FCO2与温度间存在显著的线性相关关系,Wang 等(2003)在兴安落叶松(Larixgmelinii)的研究中指出,树干FCO2与温度间的线性相关关系仅存在于早上。McGuire等(2007)认为大径级FCO2与树干温度间的指数相关性较小径级高,这可能是由于不同径级树木体内活细胞的数量以及CO2扩散到大气中的速率存在差异所致。本研究计算出4个径级树干呼吸的Q10值分别为1.64,1.55,1.47和1.60,跟以往对针叶树的计算结果相近,如石新立等(2010)计算出红松(Pinuskoraiensis)和兴安落叶松的Q10为1.25和1.69,黄玮等(2009)得出马尾松树干南、北面的Q10为1.86和1.57,Gruber等(2009)则计算出五针松(Pinuscembra)生长季和停滞期的Q10为2.0和1.81。各径阶的Q10值间不存在显著性差异,与Q10值随DBH增加而减小的结论不同(Arakietal., 2014; Kimetal., 2007)。这可能是由于树体大小和树皮厚度差异导致的树木热量交换速度(Damesinetal., 2002)、树干径向CO2扩散受阻程度(Ceschiaetal., 2002)和扩散距离(Hölttaetal., 2009)等不同造成的。因此,不同径级树木的Q10值可能存在差异,不能简单地统一利用Q10=2计算树干呼吸速率,以免造成森林生态系统树干呼吸速率的估算偏差。
马尾松的FCO2随树木胸径增大而增加,两者间呈显著的线性正相关关系,与前人的结论一致(许飞等, 2011; Arakietal., 2010; Britoetal., 2010)。这是由于随着树木胸径增大、边材宽度增加(Wangetal., 2008b),维持代谢和合成新物质所需要的能量相应也增加,从而导致生长呼吸量和维持呼吸量增加(Cesehiaetal., 2002)。然而,FCO2与DBH间的相关关系也存在一定的不确定性,如肖复明等(2005)得出湖南会同杉木FCO2与直径呈负相关关系; 许飞等(2011)对东北14个温带树种的研究表明,阔叶树FCO2与直径呈二次多项式函数关系; 石新立等(2010)得出白桦(Betulaplatyphylla)的呼吸通量随直径增大呈先增后减的趋势。综合以上分析,树干呼吸与胸径间的关系比较复杂,在对树干呼吸进行尺度扩展时,不能采用统一的扩展方法,以免出现较大的偏差。考虑到树干氮含量、边材体积等因子的测定比较困难且存在破坏性,DBH不但测定方便,而且还是森林调查中最常用的一个指标,因而依然可以考虑将其作为树木整株或林分水平尺度上树干呼吸量估测的一个实用指标(许飞等, 2011)。
5 结论
马尾松4个径级的FCO2均呈现出一致的单峰季节变化趋势,与树干温度的变化较为一致。生长季的FCO2显著高于非生长季,且在生长季各径级的FCO2间存在显著性差异。马尾松FCO2与树干温度间呈极显著的自然指数回归关系,但回归模型的参数及其决定系数因径阶大小而异,大径级FCO2与树干温度的相关性较小径阶更高。马尾松FCO2随径级的增大而增加,FCO2与胸径间呈极显著的正线性相关关系,因此胸径可以作为推算某些树种整株树木或林分水平树干呼吸的一个简便实用的指标。此外,相比温度自然指数函数模型,同时考虑树干温度和胸径因子的二元线性回归模型可以更好地解释树干CO2释放速率。
黄 玮, 朱锦懋, 黄儒珠, 等. 2009. 福建长汀冬季植物树干CO2释放速率日变化特征. 亚热带资源与环境学报, 4(1): 38-46.
(Huang W, Zhu J M, Huang R Z,etal. 2009. Diurnal variation of plant stem CO2efflux rates in winter, Changting County, Fujian. Journal of Subtropical Resource and Environment, 4(1): 38-46. [in Chinese])
马泽清, 刘琪璟, 徐雯佳, 等. 2007. 江西千烟洲人工林生态系统的碳蓄积特征. 林业科学, 43(11): 1-7.
(Ma Z Q, Liu Q J, Xu W J,etal. 2007. Carbon storage of artificial forest in Qianyanzhou, Jiangxi Province. Scientia Silvae Sinicae, 43(11): 1-7. [in Chinese])
石新立, 王传宽, 许 飞, 等. 2010. 四个温带树种树干呼吸的时间动态及其影响因子. 生态学报, 30(15): 3994-4003.
(Shi X L, Wang C K, Xu F,etal. 2010. Temporal dynamics and influencing factors of stem respiration for four temperate tree species. Acta Ecologica Sinica, 30(15): 3994-4003. [in Chinese])
王 淼, 姬兰柱, 李秋荣, 等. 2005. 长白山地区红松树干呼吸的研究. 应用生态学报, 16(1): 7-13.
(Wang M, Ji L Z, Li Q R,etal. 2005. Stem respiration ofPinuskoraiensisin Changbai Mountains. Chinese Journal of Applied Ecology, 16(1): 7-13. [in Chinese])
肖复明, 汪思龙, 杜天真, 等. 2005. 湖南会同林区杉木人工林呼吸量测定. 生态学报, 25(10): 3472-3481.
(Xiao F M, Wang S L, Du T Z,etal. 2005. Respiration of Chinese fir in plantations in Huitong, Hu’nan Province. Acta Ecologica Sinica, 25(10): 3472-3481. [in Chinese])
许 飞, 王传宽, 王兴昌. 2011. 东北东部14个温带树种树干呼吸的种内种间变异. 生态学报, 31(13): 3581-3589.
(Xu F, Wang C K, Wang X C,etal. 2011. Intra- and inter-specific variations in stem respiration for 14 temperate tree species in northeastern China. Acta Ecologica Sinica, 31(13): 3581-3589.[in Chinese])
严玉平, 沙丽清, 曹 敏. 2009. 西双版纳三叶橡胶林树干呼吸特征. 生态学报, 29(4): 1841-1847.
(Yan Y P, Sha L Q, Cao M. 2009.Stem respiration characteristics of rubber (Heveabrasiliensis) plantations in Xishuangbanna. Acta Ecologica Sinica, 29(4): 1841-1847. [in Chinese])
Amthor J S. 2000. The McCree-de Wit-Penning de Vries-thornley respiration paradigms: 30 years later. Annals of Botany, 86: 1-20.
Araki M G, Kajimoto T, Han Q M,etal. 2014. Effect of stem radial growth on seasonal and spatial variations in stem CO2efflux ofChamaecyparisobtuse. Trees, 29(2): 499-514.
Araki M G, Utsugi H, Kajimoto T,etal. 2010. Estimation of whole-stem respiration, incorporating vertical and seasonal variations in stem CO2efflux rate ofChamaecyparisobtusatrees. Journal of Forest Research, 15(2):115-122.
Brito P, Morales D, Wieser G,etal. 2010. Spatial and seasonal variations in stem CO2efflux ofPinuscanariensisat their upper distribution limit. Trees, 24(3): 523-531.
CeschiaÉ, Damesin C, Lebaube S,etal. 2002. Spatial and seasonal variations in stem respiration of beech trees (Fagussylvatica). Annals of Forest Sciences, 59 (8): 801-812.
Damesin C, Ceschia É, LeGoff N,etal. 2002. Stem and branch respiration of beech: from tree measurements to estimations at the stand level. New Phytologist, 153(1): 159-172.
Edwards N T, Tschaplinski T J, Norby R J. 2002. Stem respiration increases in CO2-enriched sweetgum trees. New Phytologist, 155(2): 239- 248.
Etzold S, Zweifel R, Ruehr N K,etal. 2013. Long-term stem CO2concentration measurements in Norway spruce in relation to biotic and abiotic factors. New Phytologist, 197(4): 1173-1185
Gruber A, Wieser G, Oberhuber W. 2009. Intra-annual dynamics of stem CO2efflux in relation to cambial activity and xylem development inPinuscembra. Tree Physiology, 29(5): 641-649.
Höltta T, Kolari P. 2009. Interpretation of stem CO2efflux measurements. Tree Physiology, 29(11): 1447-1456.
Kim M H, Nakane K, Lee J T,etal. 2007. Stem/branch maintenance respiration of Japanese red pine stand. Forest Ecology and Management, 243(2/3): 283-290.
Lavigne M B, Little C H A, Riding R T. 2004. Changes in stem respiration rate during cambial reactivation can be used to refine estimates of growth and maintenance respiration. New Phytologist, 162(1): 81-93.
Lavigne M B, Ryan M G. 1997. Growth and maintenance respiration rates of aspen, black spruce and jack pine stems at northern and southern BORLAS sites. Tree Physiology,17 (8/9):543-551.
Maier C A, Clinton B D. 2006. Relationship between stem CO2efflux, stem sap velocity and xylem CO2concentration in young loblolly pine trees. Plant Cell and Environment, 29(8): 1471-1483.
McGuire M A, Cerasoli S, Teskey R O. 2007. CO2fluxes and respiration of branch segments of sycamore (PlatanusoccidentalisL) examined at different sap velocities, branch diameters and temperatues. Journal of Experimental Botany, 58(8): 2159-2168.
Teskey R O, Saveyn A, Steppe K,etal. 2008. Origin fate and significance of CO2in tree stems. New Phytologist,177(1): 17-32.
Vose J, Ryan M. 2002. Seasonal respiration of foliage, fine roots and woody tissues in relation to growth tissue N, and synthesis. Global Change Biology, 8(2): 182-193.
Wang M, Wu Y X, Wu J L.2008a. Stem respiration of dominant tree species in broad-leaved Korean pine mixed forest in Changbai Mountains. Chinese Journal of Applied Ecology, 19(5): 956-960.
Wang X C, Wang C K, Zhang Q Z,etal. 2008b. Growth characteristics of heartwood and sapwood of the major tree species in northeastern China. Scientia Silvae Sinicae, 44(5): 102-108.
Wang W J, Yang F J, Zu Y G,etal. 2003. Stem respiration of a Larch (Larixgmelini) plantation in northeast China. Acta Botanica Sinica, 45(12): 1387-1397.
Wieser G, Gruber A, Bahn M,etal. 2009. Respiratory fluxes in a Canary Islands pine forest. Tree Physiology, 29(3): 457-466.
Zha T S, Kellanaki S, Wang K Y,etal. 2004. Seasonal and annual stun respiration of Scots pine trees under boreal conditions. Annals of Botany, 94(6): 889-896.
Zach A, Horna V, Leuschner C. 2010. Diverging temperature responses of tree stem CO2release under dry and wet season conditions in a tropical montane moist forest. Trees, 24(2): 285-296.
Zhu L W, Zhao P, Cai X A,etal. 2012. Effects of sap velocity on the daytime increase of stem CO2efflux from stems ofSchimasuperbatrees. Trees, 26(2): 535-542.
(责任编辑 于静娴)
StemCO2EffluxRatioofPinusmassonianaofVariousDBHClassesandItsSensitivitytoTemperature
Tu Jie Fan Houbao Wang Yonggang Li Zhipeng
(ResearchInstituteofEcologyamp;EnvironmentalSciences,NanchangInstituteofTechnologyJiangxiProvincialKeyLaboratoryforRestorationofDegradedEcosystemsamp;WatershedEcohydrologyNanchang330099)
【Objective】 Stem CO2efflux rates of various DBH classes and its sensitivity to temperature were investigated of the important pioneer tree speciesPinusmassonianain the degraded red soil region. The aims of this study were to providefundamental data for evaluating the carbon sink contribution of stem respiration fromPinusmassonianaplantation to the ecosystem.【Method】 We conducted year-roundinsituchamber measurements ofFCO2on twelve stems ofPinusmassonianausing an automated, portable gas-exchange system (LC PRO-SD, ADC BioScientific Ltd., Hoddesdon, UK), which was equipped with an infrared gas analyzer and a transparent plexiglas cuvette. The sample trees were selected from four DBH classes (15.9-17.9, 18.9-20.5, 22.8-24.8, and 27.1-29.8 cm) with three replicates. At least 24 h prior to measurements, loose bark was removed from the surface of measurement and then collars were mounted horizontally on the north-facing stems at about 135 cm above-ground with 100% silicone sealant. During the process of measurements, the chamber was wrapped with aluminum foil to avoid excess heating by direct sun exposure. Stem temperature at 1 cm depth (Ts) were recorded synchronously. The measurements were conducted from 8:00 to 17:00 for the selected days each month between January and December in 2015 to ensure that the temperature range of data was collected as widely as possible. The temporal fluctuations ofFCO2and difference between various DBH classes were analyzed. And the exponential regression models were also established betweenFCO2andTs.【Result】 There was clear seasonality inFCO2for four diameter classes following similar remarkable single-peak patterns, with the maximum values occurring in July and the minimum in January. Obviously, the monthlyFCO2values were significantly higher(Plt;0.01)during the fast-growing season than in the slow-growing season. The values ofFCO2differed significantly(Plt;0.01)among various DBH classes during the growing season but not in the non-growing season. Significant exponential relationships were found betweenFCO2and Ts, with the correlations of determination higher in the bigger DBH classes than those in the smaller DBH classes. Furthermore, the values of temperature sensitivity (Q10) for four diameter classes varied from 1.47 to 1.64, which were within the range of values previously reported for most conifers. Additionally,FCO2were significantly correlated positively with DBH(R2=0.89), suggesting that DBH could be used as a proxy for predicting stem respiration for conifers.【Conclusion】 Therefore, the temporal variations inFCO2and difference between various DBH classes should be simultaneously considered when estimating forest carbon budgets so as to improve the accuracy of evaluation.
Pinusmassoniana; stem temperature; stem CO2efflux ratio; diameter classes
10.11707/j.1001-7488.20171017
2016-05-22;
2017-08-28。
国家自然科学基金项目(31260172)。
S718
A
1001-7488(2017)10-0154-06
猜你喜欢
吉林林业科技(2023年6期)2023-11-20 02:13:16
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
内蒙古林业调查设计(2021年5期)2022-01-05 02:50:40
绿色科技(2019年5期)2019-11-29 13:17:37
林业勘察设计(2017年4期)2017-07-06 08:46:12
广西林业科学(2016年1期)2016-03-20 05:33:00
广西林业科学(2016年1期)2016-03-20 05:32:59
广西林业科学(2016年1期)2016-03-20 05:32:58
