
亮腹釉小蜂不同发育阶段的形态及寄生行为
2017-12-01 09:50:21李翌菡周雅婷沈祖乐邱宝利
华南农业大学学报 2017年6期
李翌菡,周雅婷,,沈祖乐,邱宝利,
(1 广东省生物农药创制与应用重点实验室/广东省农业害虫生物防治工程技术研究中心,广东 广州 510640;2 华南农业大学 农学院/生物防治教育部工程研究中心,广东 广州 510642)
亮腹釉小蜂不同发育阶段的形态及寄生行为
李翌菡1,周雅婷1,2,沈祖乐2,邱宝利1,2
(1 广东省生物农药创制与应用重点实验室/广东省农业害虫生物防治工程技术研究中心,广东 广州 510640;2 华南农业大学 农学院/生物防治教育部工程研究中心,广东 广州 510642)
【目的】研究柑橘木虱Diaphorina citri优势寄生蜂亮腹釉小蜂Tamarixia radiata的个体发育形态学及寄生行为,为柑橘黄龙病的生物防控提供理论和技术指导。【方法】在温度为(26±1) ℃、相对湿度为60%~80%的试验条件下,观察亮腹釉小蜂寄生行为及个体发育形态,并测量形态学指标。【结果】亮腹釉小蜂寄生柑橘木虱3~5龄若虫产卵;亮腹釉小蜂整个发育历程分为卵、幼虫、蛹、成虫4个阶段;室内条件下其世代发育历期约为13.40 d;雌虫个体明显大于雄虫;雄虫触角刚毛发达,几乎覆盖整个触角基节,而雌虫触角刚毛很短,触角基节清晰可见;雌虫腹部背面黄色区域明显大于雄虫。【结论】亮腹釉小蜂为柑橘木虱体外寄生蜂,雌、雄成虫形态有显著差异。
柑橘木虱;亮腹釉小蜂;寄生行为;发育形态;生物防治
柑橘木虱Diaphorina citri属半翅目Hemiptera木虱科Psyllidae,广泛分布于亚洲的大部分地区以及北美洲、南美洲、拉丁美洲和非洲等部分国家的柑橘产区,是柑橘黄龙病最重要的传播媒介[1-3]。有效防控柑橘木虱,减少其种群发生及危害,是控制柑橘黄龙病传播蔓延的关键[4]。化学农药的频繁使用,造成了柑橘木虱抗药性增加、农药残留以及生态污染等一系列问题,利用天敌昆虫开展柑橘木虱的生物防治,已成为近年来柑橘木虱综合防控研究的核心与热点[5-6]。
亮腹釉小蜂Tamarixia radiata是柑橘木虱的优势寄生蜂种[7-9]。田间寄生率调查结果表明,在广东地区,秋季亮腹釉小蜂对柑橘木虱的自然寄生率可以达到46%[10],在美国佛罗里达州寄生率为39%~56%[11],而在巴西的一些柑橘园内亮腹釉小蜂对柑橘木虱的自然寄生率达到了88.8%[12]。钱景泰等[13]通过在实验网室内饲养亮腹釉小蜂发现,亮腹釉小蜂寄生柑橘木虱5龄若虫后,卵发育至成虫的存活率高达90.4%,且表现出高度的寄主专化性。这些研究表明亮腹釉小蜂对柑橘木虱的田间防治效果良好。然而,由于亮腹釉小蜂个体微小,鲜见关于其个体发育形态学的研究报道,这在一定程度上限制了人们对亮腹釉小蜂发育生物学的认识及其在柑橘木虱防治中的应用。本文研究亮腹釉小蜂发育形态学及其寄生行为,以期为柑橘木虱的有效绿色防控提供参考。
1 材料与方法
1.1 材料
试验在生物防治教育部工程研究中心(华南农业大学)及其网室内进行。
九里香Murraya exotica幼苗购自广州市增城正果镇,盆栽于网室内,每盆种植1~2株,保证室内阳光充足,定时浇水,定期施肥、修剪,让其有充足的嫩梢嫩枝供试验使用。
柑橘木虱初始种群2014年9月采自华南农业大学,以九里香饲养于工程中心网室内,繁殖后代种群作为试验用虫;亮腹釉小蜂初始种群2015年6月采集于华南农业大学校内九里香植株上,此后在工程中心实验室内繁殖,建立九里香-柑橘木虱-亮腹釉小蜂3级营养研究体系。
1.2 方法
试验条件为温度(26±1) ℃,相对湿度60%~80%,光周期为14 h光:10 h暗,光照度为3 000 lx。1.2.1 亮腹釉小蜂寄生行为观察 采摘带有大量柑橘木虱若虫的九里香嫩芽,用棉花包住枝条断口加水保持湿润,将其放入底部垫有滤纸的玻璃培养皿中,用扎孔的保鲜膜封口,接入羽化时间少于48 h的亮腹釉小蜂雌蜂,在体视解剖镜(Motic K-400L)下观察其寄生产卵行为,试验观察30头雌蜂。
1.2.2 亮腹釉小蜂个体发育形态测定 选取带有大量柑橘木虱若虫的九里香嫩芽,套袋处理,接入充足的亮腹釉小蜂雌蜂,待其产卵12 h后,赶走所有的亮腹釉小蜂,每天定时翻转检查柑橘木虱若虫被亮腹釉小蜂寄生的情况,选取被寄生的木虱若虫在体视镜下观察,最后用联机显微镜(Stereo discovery V20)记录亮腹釉小蜂的发育过程。
收集亮腹釉小蜂雌、雄成虫标本,在联机显微镜下观察并拍照,测量雌、雄成虫的头长、头宽、体长、体宽等指标;试验雌、雄虫各重复20头。
1.3 数据处理
试验数据用Excel软件进行计算及整理,SPSS17.0软件进行独立样本t检验。
2 结果与分析
2.1 亮腹釉小蜂产卵寄生行为观察
亮腹釉小蜂的整个寄生过程可分为5个部分,首先是寄主定位,亮腹釉小蜂进入柑橘木虱若虫所在的栖息场所后,在九里香嫩枝上快速爬动,触角不停打探,搜索寄主;其次是寄主检查,检查寄主时,亮腹釉小蜂或跳到柑橘木虱若虫背部,或绕若虫2~3圈检查,期间用触角不停敲打木虱若虫背部,以此判断是否做出寄生行为或放弃该寄主;接下来是寄主选择,由于柑橘木虱若虫聚集分布在九里香嫩芽上,亮腹釉小蜂一般会对一定范围内的木虱若虫逐个进行寄主检查,检查完毕后,偏好健康的4~5龄的柑橘木虱若虫进行寄生产卵;第4步是产卵,当选定寄主后,亮腹釉小蜂停留在木虱若虫旁并调整身体方向,背向寄主,将产卵器插到木虱若虫胸腹连接处与叶片之间,身体高频振动,将卵产在木虱若虫胸腹连接处的凹槽处;最后一步是清理,产卵完后,雌蜂左右大幅摆动身体,拔出产卵器,使卵黏附在木虱若虫腹侧处,用后足摩擦清理产卵器,以除去产卵器上黏带的其他物质,之后用前足梳理触角以及头部,休憩。
2.2 亮腹釉小蜂的个体发育形态
亮腹釉小蜂世代发育历期为(13.40±0.16) d(n=30),整个发育过程经过卵、幼虫、蛹、成虫4个阶段。卵孵化成幼虫约需2.00 d;幼虫阶段分为4个龄期,整个发育阶段约为5.28 d;老熟幼虫经过1.12 d的预蛹期,进入蛹期,成熟蛹为黑褐色,蛹经过5.23 d羽化为成虫。
亮腹釉小蜂的世代发育过程如图1所示。亮腹釉小蜂产卵在柑橘木虱若虫胸腹连接处,初产卵呈乳白色米粒状,质地较硬,表面有黏性,黏附在寄主体表;1龄幼虫孵化后,质地变柔软,逐渐变成透明状,直接吸附在木虱若虫腹侧,吸取寄主体液;幼虫发育至2龄,体型变大,并开始吐丝使木虱若虫黏附在寄主植物上,寄主若虫行动力减弱;幼虫发育到3~4龄后,寄主木虱若虫因被寄生蜂幼虫吸食而干瘪,完全黏附于寄主植物上,变成僵尸状,其外表呈深褐色,扁平,据此可从寄主背部明显分辨出被寄生的柑橘木虱若虫与健康的柑橘木虱若虫,翻开寄主外壳,可发现亮腹釉小蜂4龄幼虫几乎占据寄主木虱整个腹面的4/5。亮腹釉小蜂高龄幼虫为浅黄色,透明状,质地柔软,与柑橘木虱若虫颜色相近,可清晰观测到幼虫体内橙红色中肠蠕动消化。
亮腹釉小蜂蛹为典型的离蛹,蛹初期为浅黄色,复眼红褐色,头部、胸部、腹部区分明显。随着蛹逐渐成熟,颜色不断加深,成熟蛹为黑褐色,复眼颜色加深,体长为 987.79 μm,体宽为 456.76 μm,头、眼、翅、足清晰可见。成虫羽化时,首先在柑橘木虱若虫前胸处咬出1个圆形孔洞,成为羽化孔,成虫从羽化孔中钻出寄主。
亮腹釉小蜂雌、雄成虫各形态指标见表1,其复眼均为深红色,两者的区别主要有以下几点:亮腹釉小蜂雌、雄成虫个体大小有较大区别,雌、雄成虫体长 (t=3.71,p<0.05)和体宽 (t=3.58,p<0.05)差异显著,雌成虫体长约为1 092.35 μm,体宽为383.71 μm,雄成虫体长约为971.24 μm,体宽约为328.82 μm,雌虫体型明显大于雄虫,雌、雄虫头长(t=3.68,p<0.05)和头宽(t=3.26,p<0.05)也差异显著;其次,雌、雄成虫可通过触角进行辨别,雄虫触角刚毛发达,几乎覆盖整个触角基节,而雌虫触角上的刚毛短小,触角基节清晰可见(图2);最后,雌、雄成虫腹部颜色有一定的区别,雌、雄成虫腹部中间均为黄色,边缘为黑色,但雌虫腹部背面黄色部分明显多于雄虫(图2)。
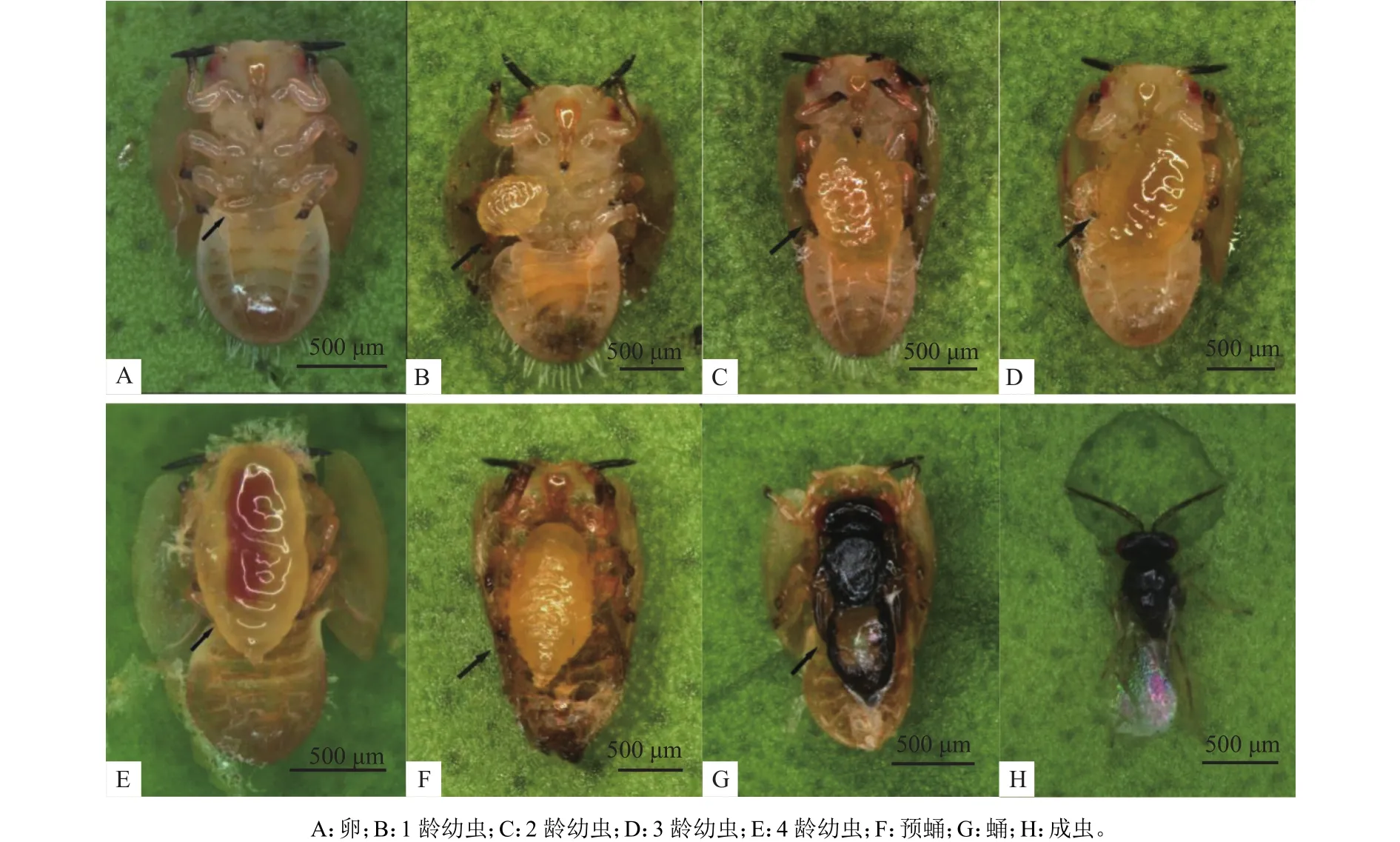
图1 亮腹釉小蜂生活史Fig. 1 The life history of Tamarixia radiata

表1 亮腹釉小蜂成虫形态1)Tab. 1 The morphology of Tamarixia radiata adult μm

图2 亮腹釉小蜂雌、雄成虫标本Fig. 2 Specimens of Tamarixia radiata female and male adults
3 讨论与结论
近年来,柑橘黄龙病给世界范围内的柑橘产业造成了巨大的经济损失[14-15],作为柑橘黄龙病的重要传播媒介,柑橘木虱的有效防治显得极其重要。随着对柑橘木虱天敌昆虫资源调查与利用研究的不断深入,包括寄生性天敌和捕食性天敌在内的多个类群资源都受到了关注,而亮腹釉小蜂因其寄生效率高、寄主专一性等特点[9,15],成为柑橘木虱寄生性天敌研究利用的重点对象。本研究结果表明,亮腹釉小蜂对柑橘木虱的寄生方式为体外寄生,整个寄生行为可分为寄主定位、寄主检查、寄主选择、产卵、清理5个部分。与大多数寄生蜂类似,亮腹釉小蜂寄生产卵之前,会对柑橘木虱若虫进行检查,选择对后代有利的寄主。至于亮腹釉小蜂在长期进化过程中,为何形成了外寄生方式,其生理选择机理如何,尚需进一步深入研究。
本研究中,在(26±1) ℃、相对湿度60%~80%条件下,亮腹釉小蜂整个发育历期为13.40 d,而Gómez-Torres等[16]研究发现在30℃、相对湿度60%~80%条件下,亮腹釉小蜂的世代发育历期为11.78 d,在一定范围内,随着温度的升高,亮腹釉小蜂的发育速度加快,世代历期缩短,这符合温度对生物体发育影响的自然规律。
通过观察发现,亮腹釉小蜂雌、雄成虫外形有显著差异,在显微镜下区别明显,其主要区别是体型大小、触角以及腹部背面颜色。亮腹釉小蜂雄蜂触角有发达的刚毛,这可能与其搜寻雌蜂的能力有关,亮腹釉小蜂93%以上的雌蜂只在羽化第1天内进行交配,极少在羽化后第2天有交配行为[13],推测雌蜂这一交配现象要求雄蜂有更强的搜寻能力。
近年来国内外对亮腹釉小蜂的生物学特性研究有了较为系统的进展,除雌成虫寿命、单雌产卵量的特性外[6,16-17],相关研究还发现亮腹釉小蜂雌成虫会取食低龄的柑橘木虱若虫以及若虫分泌的白色蜜露,在一定程度上减少了木虱种群的数量以及煤污病的发生几率[18-20]。但是,亮腹釉小蜂作为柑橘木虱的优势寄生蜂,在利用其防控柑橘木虱的应用方面尚有许多未知问题,比如蜂卡的制作、储存、田间释放技术等。本研究通过观察亮腹釉小蜂的发育过程和形态特征,一方面可以为了解柑橘木虱其他寄生蜂的个体发育提供借鉴,另一方面也有助于推动柑橘木虱寄生蜂的规模化繁殖技术研究,从而进一步推动柑橘木虱生物防治的研究与应用。
[1]BOVÉ J M. Huanglongbing: A destructive, newly-emerging, century-old disease of citrus[J]. J Plant Pathol,2006, 88(1): 7-37.
[2]HALL D G, RICHARDSON M L, AMMAR E D, et al.Asian citrus psyllid, Diaphorina citri, vector of citrus Huanglongbing disease[J]. Entomol Exp Appl, 2013,146(2): 207-223.
[3]代晓彦, 李翌菡, 许炜明, 等. 亚洲柑橘木虱 2 株高致病力病原真菌菌株的筛选[J]. 华南农业大学学报, 2016,37(5): 62-65.
[4]代晓彦, 李翌菡, 沈祖乐, 等. 球孢白僵菌与玫烟色棒束孢制剂对柑橘木虱的防治[J]. 华南农业大学学报, 2017,38(1): 63-68.
[5]HODDLE M S, HODDLE C D. Classical biological control of Asian citrus psyllid with Tamarixia radiata in urban Southern California[J]. Citrograph, 2013, 4(2):52-58.
[6]SULE H, MUHAMAD R, OMAR D, et al. Parasitism rate, host stage preference and functional response of Tamarixia radiata on Diaphorina citri[J]. Int J Agric Biol, 2014, 16(4): 783-788.
[7]CHIEN C C. The role of parasitoids in the pest management of citrus psyllid[C]//Proceedings of the Symposium on Research and Development of Citrus in Taiwan.Taiwan: Taichung, 1995: 245-261.
[8]代晓彦, 任素丽, 周雅婷, 等. 黄龙病媒介昆虫柑橘木虱生物防治新进展[J]. 中国生物防治学报, 2014, 30(3):414-419.
[9]BARR N B, HALL D G, WEATHERSBEE A A, et al.Comparison of laboratory colonies and field populations of Tamarixia radiata, an ecto-parasitoid of the Asian citrus psyllid, using internal transcribed spacer and cytochrome oxidase subunit I DNA sequences[J]. J Econ Entomol, 2009, 102(6): 2325-2332.
[10]YANG Y P, HUANG M D, BEATTIE G A, et al. Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: A status report for China[J]. Int J Pest Manage, 2006, 52(4): 343-352.
[11]QURESHI J A, ROGERS M E, HALL D G, et al. Incidence of invasive Diaphorina citri(Hemiptera: Psyllidae)and its introduced parasitoid Tamarixia radiata (Hymenoptera: Eulophidae) in Florida citrus[J]. J Econ Entomol,2009, 102(1): 247-256.
[12]PARRA J R P, ALVES G R, DINIZ A J F, et al. Tamarixia radiata (Hymenoptera: Eulophidae) ×Diaphorina citri(Hemiptera: Liviidae): Mass rearing and potential use of the parasitoid in Brazil[J]. J Integr Pest Manag, 2016,7(1): 5.
[13]钱景秦, 朱耀沂, 古琇芷. 亮腹釉小蜂(Tamarixia radiata)之形态, 生活史及其寄生策略[J]. 中华昆虫, 1991,11(4): 264-281.
[14]TEXEIRA D D C, AYRES J, KITAJIMA E W, et al.First report of a Huanglongbing-like disease of citrus in São Paulo State, Brazil and association of a new Liberibacter species, “Candidatus Liberibacter americanus”, with the disease[J]. Plant Dis, 2005, 89(1): 107.
[15]MEYER J M, HOY M A. Molecular survey of endosymbionts in Florida populations of Diaphorina citri (Hemiptera: Psyllidae) and its parasitoids Tamarixia radiata(Hymenoptera: Eulophidae) and Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae)[J]. Fla Entomol, 2008,91(2): 294-304.
[16]GÓMEZ-TORRES ML, NAVA D E, PARRAJ R P. Life table of Tamarixia radiata (Hymenoptera: Eulophidae)on Diaphorina citri (Hemiptera: Psyllidae) at different temperatures[J]. J Econ Entomol, 2012, 105(2): 338-343.
[17]章玉苹, 李敦松, 黄少华, 等. 柑桔木虱的生物防治研究进展[J]. 中国生物防治学报, 2009, 25(2): 160-164.
[18]HOY M A, NGUYEN R, JEYAPRAKASH A. Classical biological control of Asian citrus psyllid in Florida[J/OL]. Integrated Pest Management Florida: University of Florida. (2012-03-13) [2016-10-10]. http://ipm. ifas.ufl. edu/agriculture/citrus/psyllid.shtml.
[19]SKELLEY L H, HOY M A. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine[J]. Biol Control, 2004, 29(1): 14-23.
[20]MANN R S, QURESHI J A, STANSLY P A, et al. Behavioral response of Tamarixia radiata Waterston (Hymenoptera: Eulophidae) to volatiles emanating from Diaphorina citri Kuwayama (Hemiptera: Psyllidae) and citrus[J].J Insect Behav, 2010, 23(6): 447-458.
Morphology and parasitic behavior of Tamarixia radiata at different development stages
LI Yihan1, ZHOU Yating1,2, SHEN Zule2, QIU Baoli1,2
(1 Key Laboratory of Bio-Pesticide Innovation and Application/Engineering Technology Research Center of Agricultural Pest Biocontrol, Guangzhou 510640, China; 2 College of Agriculture, South China Agricultural University/Engineering Research Center of Biological Control, Ministry of Education, Guangzhou 510642, China)
【Objective】To study the ontogenetic morphology and parasitic behavior of Tamarixia radiata, the predominant parasitoid of Diaphorina citri, and provide theoretical and technical information for sustainable management of Huanglongbing.【Method】The parasitic behavior and ontogenetic morphology of T. radiata were observed under laboratory conditions of (26±1) ℃ and 60% to 80% relative humility and natural lighting.Morphologic indexes of T. radiata were measured.【Result】The adult T. radiata females lay eggs on 3rd to 5th instar nymphs of D. citri. The life cycle of T. radiata includes the egg, larva, pupa and adult stages. The developmental duration is about 13.40 d under laboratory condition. The females are distinctly larger than males in body size. The setae of male antennas are well developed and almost cover all the antenna segments, while the setae of female antennas are short and the antenna segments are clearly visible. The yellow area on the dorsal surface of female abdomen is obviously larger than that of male.【Conclusion】T. radiata is an ecto-parasitoid of D. citri. There are significant morphological differences between T. radiata female and male adults.
Diaphorina citri; Tamarixia radiata; parasitic behavior; developmental morphology; biocontrol
S433.3
A
1001-411X(2017)06-0084-05
李翌菡, 周雅婷, 沈祖乐, 等. 亮腹釉小蜂不同发育阶段的形态及寄生行为[J]. 华南农业大学学报, 2017, 38(6): 84-88.
2017-02-28 优先出版时间:2017-11-06
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.S.20171106.1412.028.html
李翌菡 (1988—),女,博士研究生,E-mail:eyvonneli@hotmail.com; 通信作者: 邱宝利 (1973—),男,教授,博士,
E-mail: baileyqiu@scau.edu.cn
广州市科技计划项目(201509010023);广州市产学研协同创新重大专项(201604030029);广东省岭南水果创新团队
专项(2016LM1106);广东省农业技术需求研究与示范项目(2016LM3175)
【责任编辑 霍 欢】
猜你喜欢
特产研究(2024年1期)2024-03-12 05:40:48
声屏世界(2022年21期)2023-01-07 12:53:10
华南农业大学学报(2022年6期)2022-10-31 07:29:54
科技进步与对策(2020年22期)2020-12-08 00:47:44
贵州农业科学(2019年9期)2019-10-10 09:18:16
小学生作文·小学中高年级适用(2017年9期)2018-03-13 18:49:09
小学生作文(中高年级适用)(2017年9期)2017-10-13 08:20:53
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:02
湖南农业(2017年1期)2017-03-20 14:04:46
中国医院用药评价与分析(2016年11期)2016-12-19 08:21:14
