
辣椒炭疽病菌Colletotrichum gloeosporioides对啶氧菌酯的敏感基线及抗性突变体生物学性状
2017-11-29 03:21周建波刘慧平曹俊宇赵晓军
植物保护 2017年6期
任 璐, 周建波, 刘慧平, 曹俊宇, 殷 辉, 赵晓军*
(1. 农业有害生物综合治理山西省重点实验室, 太原 030031; 2. 山西农业大学农学院,太谷 030801; 3. 山西省农业科学院植物保护研究所, 太原 030006)
辣椒炭疽病菌Colletotrichumgloeosporioides对啶氧菌酯的敏感基线及抗性突变体生物学性状
任 璐1,2, 周建波1,3, 刘慧平2, 曹俊宇2, 殷 辉1,3, 赵晓军1,3*
(1. 农业有害生物综合治理山西省重点实验室, 太原 030031; 2. 山西农业大学农学院,太谷 030801; 3. 山西省农业科学院植物保护研究所, 太原 030006)
为评估辣椒炭疽病菌Colletotrichumgloeosporioides对啶氧菌酯的抗性风险,建立了辣椒炭疽病菌对啶氧菌酯的敏感基线,室内筛选获得了辣椒炭疽病菌抗啶氧菌酯突变体,并对突变体生物学性状进行了研究。在山西省3个未使用过啶氧菌酯及QoI类药剂的地区采集并分离到45株辣椒炭疽病菌。采用菌丝生长速率法测定其对啶氧菌酯的敏感性,结果显示,EC50值介于1.404~16.650 μg/mL,平均EC50值为(6.783±3.499)μg/mL。啶氧菌酯与水杨肟酸(SHAM)共同处理时(SHAM的处理浓度为100 μg/mL),EC50+S为0.022~0.275 μg/mL,平均(0.109±0.058)μg/mL,EC50+S呈连续性单峰曲线,且敏感性频率分布呈近似正态分布,EC50+S平均值可作为辣椒炭疽病菌对啶氧菌酯的敏感基线。且水杨羟肟酸旁路氧化贡献值F(F=EC50/EC50+S)最低为6.039,最高为301.441,平均78.026。室内诱导获得8株抗性突变体,其中低抗突变体6株,中抗突变体2株。中抗突变体无性繁殖10代,其抗性可稳定遗传。突变体产孢量和菌丝生长速率与敏感菌株相比均无显著差异,但无药条件下,突变体致病力较敏感菌株有所降低。抗感菌株均以淀粉作为碳源、硝酸钾作为氮源时利用率最高;最适pH均为5和6;菌丝的最适生长温度均为25℃。表明辣椒炭疽病菌抗啶氧菌酯突变体具有较高的适合度,对啶氧菌酯具有较高抗性风险,这一研究结论为指导生产用药,延缓抗药性发展提供理论依据。
辣椒炭疽病菌; 啶氧菌酯; 敏感基线; 抗性风险; 生物学性状
辣椒炭疽病是辣椒生产中常见的真菌性病害,也是辣椒生产过程中三大病害之一。该病害由子囊菌门炭疽菌属Colletotrichum的多个种侵染造成,如Colletotrichumgloeosporioides、C.capsici、C.acutatum等。炭疽病菌可危害辣椒叶片和果实,尤以对果实危害最为明显与严重,导致辣椒腐烂,失去经济价值。在夏季天气闷热、湿度大时易发生,造成辣椒大面积减产[1]。报道显示,在很多亚洲国家,如韩国、印度、泰国,由于该病害的发生,辣椒产量明显下降[2]。Colletotrichumspp.还可以侵染甜椒[3]。有报道指出,C.acutatum是引起韩国辣椒和甜椒炭疽病的主要致病菌,C.capsici是泰国辣椒炭疽病的主要致病菌[3-4],而在我国的各主要辣椒产区,不同省份所报道的优势致病菌有所不同[5-6],根据我们的取样和调查,C.gloeosporioides是引起山西省辣椒产区辣椒炭疽病的优势致病菌。
目前,化学防治是生产上果蔬炭疽病的主要防治手段,常用杀菌剂主要有芳烃类、有机硫类及苯并咪唑类等。已有报道显示炭疽病菌对多种常用药剂产生了不同程度的抗性。例如,Sander等[7]报道,南非芒果上的C.gloeosporioides已对多种苯并咪唑类杀菌剂敏感性降低。Maymon等[8]报道,羽扇豆和鳄梨上C.gloeosporioides对苯并咪唑类杀菌剂产生了抗性。在中国,有些省份也出现炭疽病菌对多种类型的药剂敏感性下降。曹学仁等[9]报道了海南省橡胶炭疽病菌Colletotrichumspp.对多菌灵和咪鲜胺的敏感性降低;韩国兴等[10]发现浙江省杭州地区的草莓炭疽病菌C.gloeosporioides对多菌灵和乙霉威产生了抗性,抗性频率高达90%以上;叶佳等[11]分别对辽宁和浙江的葡萄炭疽病菌群体Colletotrichumspp.对多菌灵及甲基硫菌灵的敏感性进行测定,发现病菌已产生一定程度抗性。
啶氧菌酯(picoxystrobin)于2001年由先正达在欧洲首次推出,并于2006年被杜邦公司收购,之后在拉美、北美市场登记,在中国于2012年7月获得临时登记并于同年11月正式登记,是甲氧基丙烯酸酯类杀菌剂中内吸活性最强的品种,主要用于防治蔬菜灰霉病菌Botrytiscinerea、黄瓜霜霉病菌Pseudoperonosporacubensis、香蕉黑星病菌Macrophomamusae、辣椒炭疽病菌Colletotrichumspp.和葡萄黑痘病菌Sphacelomaampelinum等。甲氧基丙烯酸酯类杀菌剂的作用位点主要是线粒体上的细胞色素b,通过抑制线粒体氧化呼吸所必经的细胞色素b和c1 之间的电子转移来阻断生物的呼吸路径,从而起到杀菌效果[12-13]。啶氧菌酯可用于防治已对14α-脱甲基抑制剂和苯并咪唑类杀菌剂产生抗性的植物病原菌。炭疽菌属Colletotrichumspp.对啶氧菌酯抗性的相关报道还很少见,但不同病原真菌对QoI类杀菌剂的抗性分子机理显示,抗性突变位点单一,抗性风险较高[14-17]。
山西省气候干燥、阳光充足、热量丰富,是中国华北地区主要的辣椒种植和生产区域。据我们调查,截至2015年,山西省辣椒种植面积近4万 hm2。在发病较为严重的山西省朔州市应县的辣椒产区,辣椒炭疽病的发病率高达30%,该病害的防治不利给生产上带来较为严重的损失。啶氧菌酯在山西省辣椒产区还未广泛应用。本研究于2014-2016年从山西省各地区的辣椒上采集并分离了炭疽病菌菌株,通过建立敏感基线,检测未使用啶氧菌酯的辣椒产区辣椒炭疽病菌的敏感性,对比抗感菌株适合度及其他生物学特性,评估辣椒炭疽病菌对啶氧菌酯的抗性风险,为生产上指导用药,延缓抗药性产生、延长药剂使用寿命提供理论依据。
1 材料与方法
1.1 供试病原菌
用于建立敏感基线的辣椒炭疽病菌菌株采自山西省晋中市3个未使用过啶氧菌酯及QoI类杀菌剂的辣椒种植地。将具有典型辣椒炭疽病病状的辣椒果实带回实验室,用无菌去离子水冲洗表面3次后,挑取病组织接种于PDA培养基中25℃培养3 d,分离纯化菌种至新的PDA平板,25℃培养5 d后进行单孢分离,共获得68株辣椒炭疽病菌菌株,经鉴定,其中45株为C.gloeosporioides,23株为C.capsici,菌株以采集地首字母+数字分别命名。本研究检测了其中45株C.gloeosporioides对啶氧菌酯的敏感性。
1.2 供试药剂
95%啶氧菌酯(picoxystrobin)原药,湖北健源化工有限公司生产。用丙酮制成10 000 μg/mL母液,4℃保存备用。
99%水杨羟肟酸(salicylhydroxamic acid,简称SHAM)分析试剂,上海伊卡生物技术有限公司生产。用丙酮制成10 000 μg/mL母液,4℃保存备用。
1.3 含药培养基制备
配制10 mg/mL啶氧菌酯及10 mg/mL水杨羟肟酸,用无菌水分别稀释成所需浓度,按照药剂与加热后冷却至50℃左右的熔融状态PDA培养基(马铃薯200 g、葡萄糖20 g、琼脂15~17 g、水1 000 mL)1∶9比例混合均匀。
1.4 辣椒炭疽病菌对啶氧菌酯及对啶氧菌酯+SHAM的敏感性测定
根据Rebollar-Alviter等[18]及Zhou等[19]的试验结果,培养基中加入SHAM可抑制病原菌的旁路氧化途径,从而更有效抑制病菌菌丝的生长。因此本研究参照Rebollar-Alviter等[18]及Markoglou等[16]的方法设计试验。根据预试验结果(未提供数据)配制含药PDA培养基,其中啶氧菌酯的终浓度分别为0、1、2、4、8、10和50 μg/mL,SHAM的终浓度为100 μg/mL。对照只在PDA培养基中加入相同浓度的SHAM。
将保存备用的菌株接种到PDA平板中培养5 d,用打孔器从菌落边缘打取5 mm菌饼接种到含不同浓度啶氧菌酯和SHAM的PDA平板上,每浓度重复处理3次。放置于25℃恒温培养箱中培养5 d,用十字交叉法测量菌落直径,计算各个浓度药剂对辣椒炭疽病菌的菌丝生长抑制率。菌丝生长抑制率(%)=[1-(待测菌落直径-5 mm)/(对照菌落直径-5 mm)]×100。对抑菌率进行线性回归分析,求出回归方程及有效抑制中浓度EC50。将啶氧菌酯单独作用下测得的敏感性记为EC50;啶氧菌酯与SHAM(100 μg/mL)共同作用下得到的EC50记为EC50+S。计算出旁路氧化的贡献值F=EC50/EC50+S。
1.5 啶氧菌酯抗性突变体的获得
在室内,采用紫外诱导和药剂驯化筛选辣椒炭疽病菌对啶氧菌酯的抗性突变菌株。紫外诱导:从来源于3个采集地的辣椒炭疽病菌中随机挑选3株敏感菌株SN13、JX3、YZ6,接种于PDA平板上培养3 d,在距20 W紫外灯10 cm距离下照射20、40、60 min后在黑暗环境下接种到含亚致死剂量(50 μg/mL)的啶氧菌酯PDA平板上,每处理重复3次,在黑暗环境中培养3~4 d,将能够在含药PDA平板上形成扇形菌落的角变区,用相同方法连续紫外照射及黑暗培养,增加啶氧菌酯的浓度,直到其可以在500 μg/mL啶氧菌酯平板上生长,测定其敏感性,确定其抗性倍数。
药剂驯化:随机选取辣椒炭疽病菌敏感菌株SN1、JX1、YZ1在PDA平板上培养5 d,在菌落边缘打取5 mm的菌饼,接种到含20 μg/mL啶氧菌酯的平板上,置于恒温培养箱中培养8 d后,采用相同的方法将菌饼接种到更高浓度的含药平板上,浓度每次增加10 μg/mL,且每增加一次算作一代,直到其可以在500 μg/mL啶氧菌酯浓度含药平板生长,测定其敏感性,确定其抗性倍数。
抗性倍数(resistance factor, RF)=抗性菌株EC50/敏感菌株EC50。当菌株抗性水平达到5倍,则表明有抗性产生,抗性水平大于5倍的菌株在整个群体中所占的比例即为抗性频率。抗性分级标准:抗性倍数<5倍为敏感;5倍≤抗性倍数<10倍为低抗;10~40倍为中抗;40倍以上为高抗[20]。
1.6 抗性突变体遗传稳定性测定
将抗性突变体在无药PDA培养基上25℃黑暗条件下培养10代,每5 d将菌株转到新的无药培养基上,转接10次,分别测其第2、4、6、8、10代菌株的EC50值。
1.7 抗性突变体生物学特性测定
选取敏感菌株JX3、SN13、YZ6与抗性突变体JX3R2、SN13R1、YZ6R2、JX3MR3、YZ6MR4进行以下试验。
1.7.1 抗性突变体生长速率及产孢量测定
将培养5 d的敏感菌株与抗性突变体,用直径5 mm的打孔器在菌落边缘相同位置打取菌块,接种于无药PDA培养基上,25℃下恒温黑暗培养,每处理重复3次,每24 h用十字交叉法测量菌落直径。培养10 d后,在每个平板中加入约10 mL灭菌水,用玻璃棒轻轻在菌落表面不断地拨动,用4层纱布过滤,定容至5 mL,充分混匀,在血球计数板上计算抗感菌株的产孢量。
1.7.2 抗性突变体与敏感菌株致病力测定
用离体组织法测定抗性突变体与敏感菌株致病力。取健康的辣椒果实用水清洗其表面,再用灭菌水冲洗3次,然后切成直径为3 cm大小的辣椒圆片。将辣椒片在75%乙醇中浸泡2 min左右,然后在浓度为150 μg/mL的啶氧菌酯药液中浸泡2 min,以无菌水浸泡为对照,最后放入带有无菌滤纸的培养皿中,采用针刺法将该病菌的孢子悬浮液接种到辣椒片中央,每个培养皿中4片,每处理4次重复。置于25℃恒温培养箱中进行培养,用潮湿的无菌滤纸进行保湿培养,3 d后用坐标纸计算病斑面积。
1.7.3 抗感菌株在不同酸碱度下的生长状况
在无菌操作台中,用0.1 mol/L的HCl和0.1 mol/L的NaOH调节PDA培养基的酸碱度,使其pH分别为3、4、5、6、7、8、9、10、11。在辣椒炭疽病菌菌落边缘打取5 mm的菌饼,接入含梯度酸碱度的PDA平板上,每处理3次重复,倒置于25℃的恒温培养箱中培养,6 d后用十字交叉法测定菌落直径。
1.7.4 抗感菌株在不同碳、氮营养条件下的生长状况
供试碳源为葡萄糖、麦芽糖、蔗糖、甘露糖、丙三醇和淀粉。供试氮源为蛋白胨、尿素、硫酸铵和硝酸钾。测定碳源对抗感菌株的影响时,将理查培养基(氯化铁FeCl30.02 g、硫酸镁MgSO42.5 g、硝酸钾KNO310 g、磷酸二氢钾KH2PO45 g、蔗糖50 g、琼脂17 g、无菌水1 000 mL)中的蔗糖换成待测碳源,研究氮源时,将理查培养基中的硝酸钾置换成待测氮源,取直径为5 mm的抗感菌株的菌饼接到不同碳、氮源平板上,每处理3次重复,25℃培养6 d后测定菌落直径。
1.7.5 抗感菌株在不同温度下的生长状况
从抗感菌株菌落边缘打取直径为5 mm的菌饼于无药PDA平板中,分别放置于15、20、25、30、35℃恒温培养箱培养,每处理设置3次重复,6 d后用十字交叉法测量菌落直径。
2 结果与分析
2.1 辣椒炭疽病菌对啶氧菌酯、啶氧菌酯+SHAM的敏感性
45株C.gloeosporioides的敏感性测定结果表明,在啶氧菌酯单独作用下,其EC50介于1.404~16.650 μg/mL,平均值为(6.783±3.499)μg/mL,EC50呈连续性变化(图1a)。在啶氧菌酯与SHAM共同作用下,其EC50+S的范围在0.022~0.275 μg/mL(图1b),平均值为(0.109±0.058)μg/mL,最大与最小值相差12.5倍,亦呈连续性变化。菌株对啶氧菌酯+SHAM的敏感性频率分布呈单峰曲线(图2),以S-W法对45株菌株的敏感性分布进行正态性检验,其W值为0.973,P=0.635>P0.05,表明病原菌对啶氧菌酯的敏感性分布呈近似正态分布,未出现敏感性明显下降的群体。因此,其EC50+S均值可作为辣椒炭疽病菌对啶氧菌酯的敏感基线。
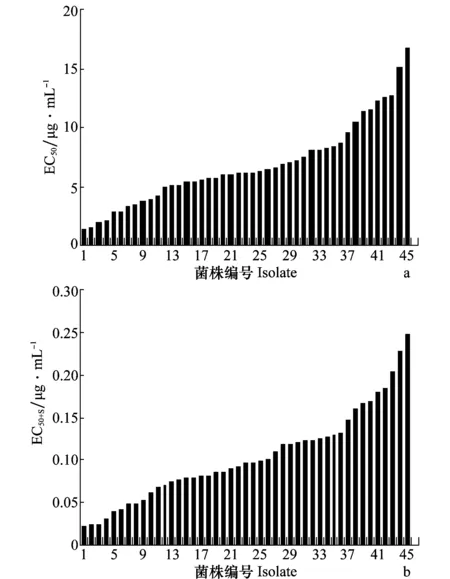
图1 啶氧菌酯(a)和啶氧菌酯+SHAM(b)对45株辣椒炭疽病菌的EC50值Fig.1 EC50 values of picoxystrobin (a) and picoxystrobin+ SHAM (b) to 45 Colletotrichum gloeosporioides isolates

图2 辣椒炭疽病菌对啶氧菌酯的敏感性频率分布Fig.2 Frequency distribution of Colletotrichum gloeosporioides isolates with different sensitivities to picoxystrobin
与啶氧菌酯单独作用相比较,啶氧菌酯与SHAM共同作用下,EC50下降明显,表明旁路氧化抑制剂SHAM对啶氧菌酯抑制C.gloeosporioides菌丝生长的活性表现出补偿作用,即增效作用,使得啶氧菌酯的抑菌活性大大加强。且旁路氧化贡献值F[F=EC50/(EC50+S)]最低为6.039,最高为301.441,平均78.026(图3),而且从图中可以明显看到4个尾点,这主要是由于SHAM在不同菌株中的旁路氧化贡献值不同所造成的。

图3 水杨肟酸旁路氧化贡献值(F值)Fig.3 Compensation efficiency value in the bypass oxidation (F values)
2.2 啶氧菌酯抗性突变体的抗性水平
在室内通过紫外诱导和药剂驯化共获得8株抗性突变体,分别为SN13R1、JX3R1、JX3R2、JX3MR3、YZ6R1、YZ6R2、YZ6R3、YZ6MR4,其中6株为低抗菌株,2株为中抗菌株(表1)。
表1室内紫外诱导及药剂驯化抗性突变体抗性水平
Table1Resistanceleveltopicoxystrobinofpicoxystrobin-resistantmutantsofColletotrichumgloeosporioidesobtainedinthelaboratoryviaUVradiationandfungicideselection
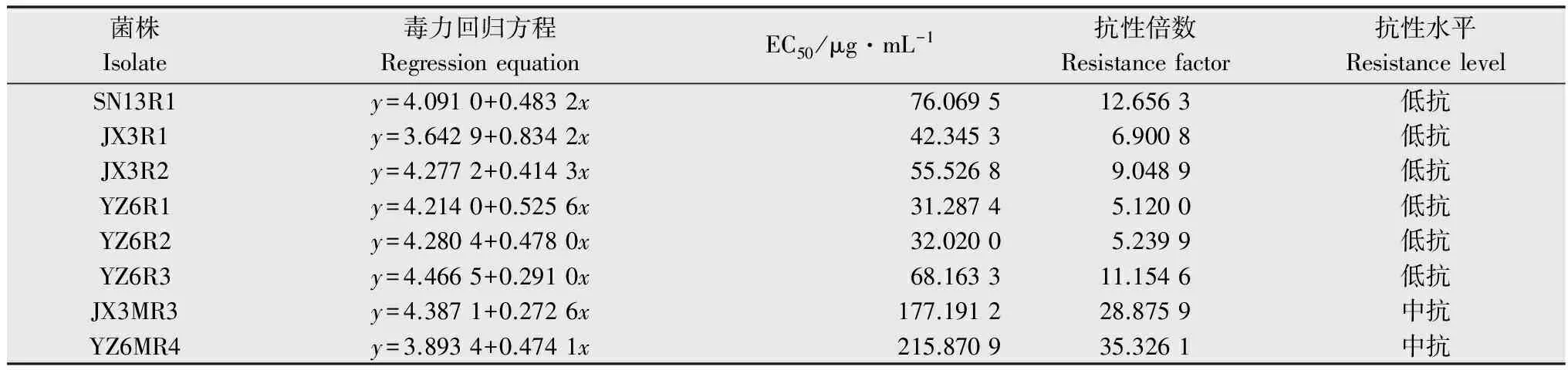
菌株Isolate毒力回归方程RegressionequationEC50/μg·mL-1抗性倍数Resistancefactor抗性水平ResistancelevelSN13R1y=4.0910+0.4832x76.069512.6563低抗JX3R1y=3.6429+0.8342x42.34536.9008低抗JX3R2y=4.2772+0.4143x55.52689.0489低抗YZ6R1y=4.2140+0.5256x31.28745.1200低抗YZ6R2y=4.2804+0.4780x32.02005.2399低抗YZ6R3y=4.4665+0.2910x68.163311.1546低抗JX3MR3y=4.3871+0.2726x177.191228.8759中抗YZ6MR4y=3.8934+0.4741x215.870935.3261中抗
2.3 抗性突变体遗传稳定性测定
从表2中可以看出,低抗菌株JX3R1、YZ6R2经过10代的无性繁殖后,抗性倍数下降较为明显,抗药性不能稳定遗传。低抗菌株SN13R1、中抗菌株JX3MR3、YZ6MR4的抗性倍数虽略有下降,但其菌丝生长良好,且经过10代的连续培养后,其抗性倍数仍然比较稳定,表明其抗药性能够遗传,不会丧失。
表2抗性突变体遗传稳定性
Table2StabilityoftheresistantmutantsofColletotrichumgloeosporioides
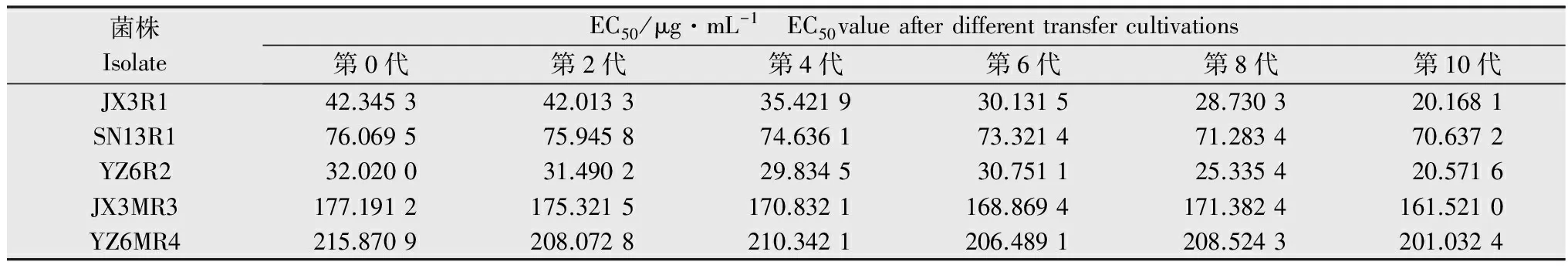
菌株IsolateEC50/μg·mL-1 EC50valueafterdifferenttransfercultivations第0代第2代第4代第6代第8代第10代JX3R142.345342.013335.421930.131528.730320.1681SN13R176.069575.945874.636173.321471.283470.6372YZ6R232.020031.490229.834530.751125.335420.5716JX3MR3177.1912175.3215170.8321168.8694171.3824161.5210YZ6MR4215.8709208.0728210.3421206.4891208.5243201.0324
2.4 抗性突变体生物学特性
2.4.1 抗性突变体与敏感菌株菌丝生长速率及产孢量
表3结果显示,低抗突变体菌丝生长速率略低于敏感菌株,但差异不显著;其中,YZ6R2、SN13R1菌丝生长与产孢量均与其敏感菌株YZ6和SN13没有显著差异。而中抗突变体JX3MR3、YZ6MR4菌丝生长速率显著低于敏感菌株JX3和YZ6;其中,JX3MR3的产孢量与其亲本敏感菌株JX3差异显著。
表3抗性突变体与敏感菌株生长速率与产孢量1)
Table3Myceliumgrowthandsporeproductionofsensitiveisolatesandresistantmutants

菌株Isolate菌落直径/cm Colonydiameter1d2d3d4d5d6d7d8d平均速率/cm·d-1Average产孢量/×106SporenumberJX31.59002.76003.92004.65005.58006.85007.85007.9800(1.03±0.43)a(5.80±0.20)cSN131.41672.95003.72174.58335.60677.25007.97008.5367(1.07±0.41)a(5.53±0.12)cdYZ61.94333.23334.00835.01006.06677.06507.97008.2417(1.03±0.47)a(6.75±0.22)aJX3R21.33332.33833.40174.25835.23336.33337.29177.5750(0.95±0.30)a(4.40±0.20)eSN13R11.23332.52173.33174.01675.19176.76177.50508.3617(1.05±0.32)a(4.96±0.32)dYZ6R21.95603.28334.06335.00506.04507.04677.94178.1333(1.02±0.50)a(6.27±0.12)aJX3MR31.55831.85172.59833.38834.78335.18676.20837.1333(0.89±0.44)b(4.00±0.10)fYZ6MR41.25501.65502.50173.29504.17504.80675.56006.2683(0.78±0.24)b(6.00±0.10)ab
1) 表中同列数据后标有不同字母者表示经Duncan氏新复极差法检验在P<0.05水平差异显著。下同。
Data followed by different lowercase letters indicate significant difference atP<0.05 level by Duncan’s new multiple range test. The same below.
2.4.2 抗性突变体与敏感菌株致病力
从表4中可知,在不含药的情况下,接种敏感菌株的所有辣椒圆盘均会产生病斑,侵染率分别为62.35%、62.63%、58.32%;抗性突变体在不含药圆盘上侵染率较敏感菌株低,接种后引起的发病面积较小,且突变体抗性倍数越高,所致发病面积及侵染率越低。低抗菌株JX3R2和YZ6R2致病力与亲本菌株差异不显著,但SN13R1与敏感菌株SN13差异明显;而中抗菌株致病力较亲本敏感菌株均显著降低。
在含50 μg/mL啶氧菌酯的辣椒圆盘上,接种敏感菌株的发病面积与侵染率均显著低于接种抗性突变体,说明抗性突变体均表现出一定的抗药性,其中,中抗菌株JX3MR3、YZ6MR4侵染率达到16.14%、34.01%。
表4抗性突变体及敏感菌株致病性
Table4Pathogenicityofresistantandsensitiveisolates

菌株Isolate发病面积/cm2 LesionareaCK50μg/mL侵染率/% InfectionfrequencyCK50μg/mLJX3(17.62±0.38)a(0.95±0.07)e62.35a3.36eSN13(17.70±0.29)a(0.96±0.07)e62.63a3.40eYZ6(16.48±0.68)b(0.96±0.07)e58.32b3.40eJX3R2(17.16±0.05)ab(2.51±0.11)d60.72ab8.89dSN13R1(16.82±1.33)b(2.95±0.28)c59.52b10.44cYZ6R2(15.75±0.19)b(2.81±0.22)c55.73b9.94cJX3MR3(16.83±0.08)b(4.56±0.04)b59.55b16.14bYZ6MR4(15.54±0.08)c(9.61±0.21)a54.99c34.01a
2.4.3 抗感菌株在不同酸碱度下的生长状况
抗感菌株在pH为5和6时生长状况均最好(表5)。敏感菌株与低抗菌株在强酸或强碱条件下生长较差,且敏感菌株与低抗菌株间差异显著,说明低抗菌株在不同pH生长条件下的适应能力低于敏感菌株。中抗菌株JX3MR3、YX6MR4在pH 3时均不能生长,但偏酸偏碱条件下,JX3MR3、YX6MR4菌落直径均大于其亲本敏感菌株,说明中抗菌株对偏酸偏碱条件的适应性均优于敏感菌株。
2.4.4 抗感菌株在不同碳、氮营养条件下的生长状况
从表6中可以看出,抗感菌株在几种供试碳源下均能正常生长,其中低抗菌株SN13R1在含有甘露糖的培养条件下生长最快,而其余菌株均在以淀粉为碳源的培养基中生长最快。敏感菌株JX3、SN13及抗性突变体JX3R2、SN13R1、YZ6R2、YX6MR4在以丙三醇作为碳源的条件下生长最慢,敏感菌株YZ6在以麦芽糖为碳源的培养基上生长速率最低,中抗菌株JX3MR3在以蔗糖为碳源的培养基上生长的最慢。说明不同病原菌对不同碳源的利用率有差异。
表5不同pH下抗感菌株的菌落直径
Table5ColonydiameterofsensitiveisolatesandresistantmutantsunderdifferentpHvalues
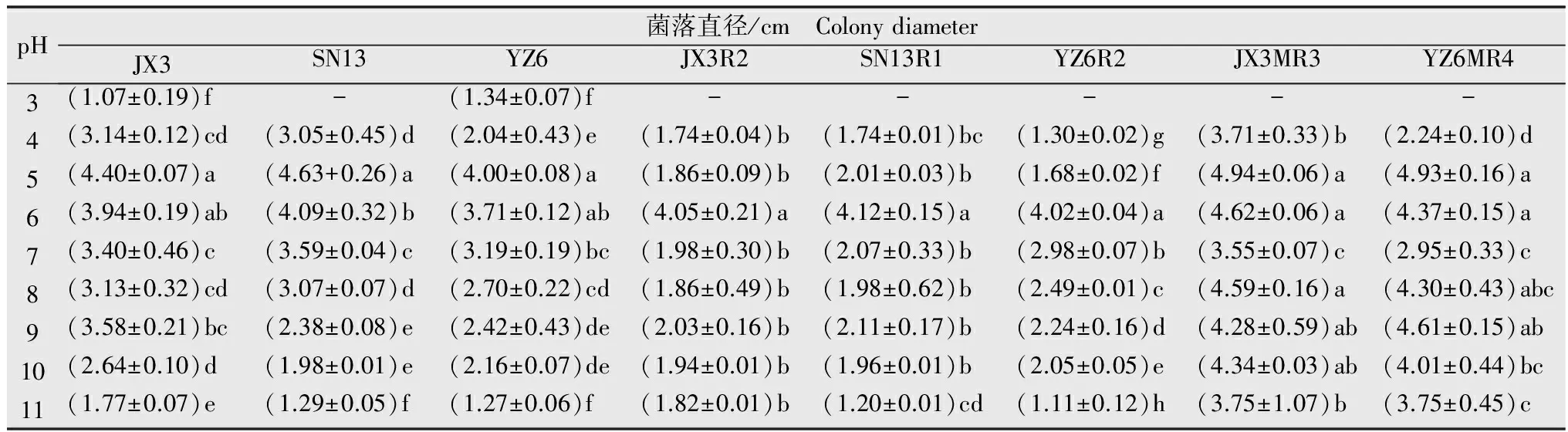
pH菌落直径/cm ColonydiameterJX3SN13YZ6JX3R2SN13R1YZ6R2JX3MR3YZ6MR43(1.07±0.19)f -(1.34±0.07)f -----4(3.14±0.12)cd(3.05±0.45)d(2.04±0.43)e(1.74±0.04)b(1.74±0.01)bc(1.30±0.02)g(3.71±0.33)b (2.24±0.10)d 5(4.40±0.07)a(4.63+0.26)a(4.00±0.08)a(1.86±0.09)b(2.01±0.03)b(1.68±0.02)f(4.94±0.06)a(4.93±0.16)a6(3.94±0.19)ab(4.09±0.32)b(3.71±0.12)ab(4.05±0.21)a(4.12±0.15)a(4.02±0.04)a(4.62±0.06)a(4.37±0.15)a7(3.40±0.46)c(3.59±0.04)c(3.19±0.19)bc(1.98±0.30)b(2.07±0.33)b(2.98±0.07)b(3.55±0.07)c(2.95±0.33)c8(3.13±0.32)cd(3.07±0.07)d(2.70±0.22)cd(1.86±0.49)b(1.98±0.62)b(2.49±0.01)c(4.59±0.16)a(4.30±0.43)abc9(3.58±0.21)bc(2.38±0.08)e(2.42±0.43)de(2.03±0.16)b(2.11±0.17)b(2.24±0.16)d(4.28±0.59)ab(4.61±0.15)ab10(2.64±0.10)d(1.98±0.01)e(2.16±0.07)de(1.94±0.01)b(1.96±0.01)b(2.05±0.05)e(4.34±0.03)ab(4.01±0.44)bc11(1.77±0.07)e(1.29±0.05)f(1.27±0.06)f(1.82±0.01)b(1.20±0.01)cd(1.11±0.12)h(3.75±1.07)b(3.75±0.45)c
表6不同碳源条件下敏感菌株和抗性突变菌落直径
Table6Colonydiametersofsensitiveisolatesandresistantmutantsindifferentcarbonsources
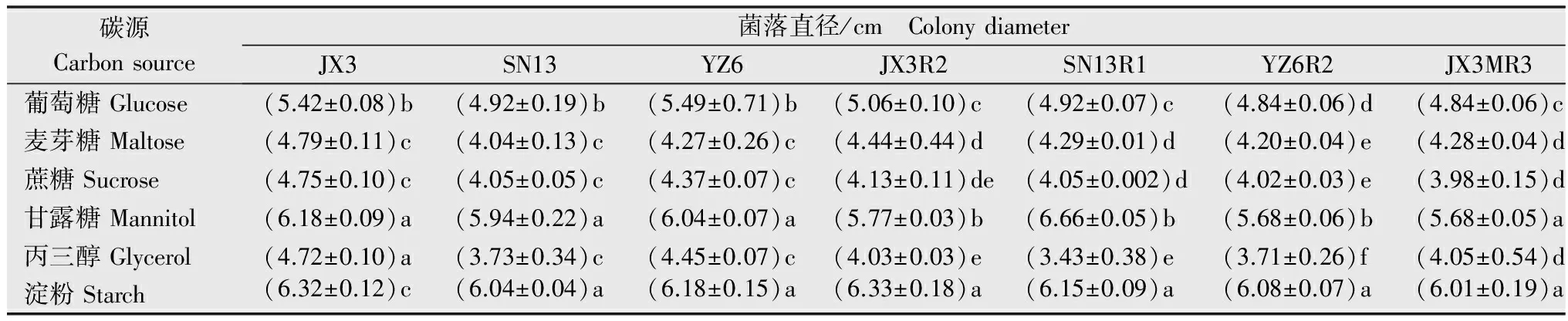
碳源Carbonsource菌落直径/cm ColonydiameterJX3SN13YZ6JX3R2SN13R1YZ6R2JX3MR3YX6MR4葡萄糖Glucose(5.42±0.08)b(4.92±0.19)b(5.49±0.71)b(5.06±0.10)c (4.92±0.07)c (4.84±0.06)d(4.84±0.06)c(4.98±0.08)b麦芽糖Maltose(4.79±0.11)c(4.04±0.13)c(4.27±0.26)c(4.44±0.44)d(4.29±0.01)d(4.20±0.04)e(4.28±0.04)d(4.33±0.50)c蔗糖Sucrose(4.75±0.10)c(4.05±0.05)c(4.37±0.07)c(4.13±0.11)de(4.05±0.002)d(4.02±0.03)e(3.98±0.15)d(4.30±0.03)c甘露糖Mannitol(6.18±0.09)a(5.94±0.22)a(6.04±0.07)a(5.77±0.03)b(6.66±0.05)b(5.68±0.06)b(5.68±0.05)a(5.96±0.19)a丙三醇Glycerol(4.72±0.10)a(3.73±0.34)c(4.45±0.07)c(4.03±0.03)e(3.43±0.38)e(3.71±0.26)f(4.05±0.54)d(4.03±0.26)c淀粉Starch(6.32±0.12)c(6.04±0.04)a(6.18±0.15)a(6.33±0.18)a(6.15±0.09)a(6.08±0.07)a(6.01±0.19)a(6.27±0.01)a
由表7可知,菌株在蛋白胨、尿素、硫酸铵、硝酸钾4种氮源培养基上都可生长,但生长差异明显。其中硝酸钾为最适氮源,且与其他氮源处理差异显著。其次是蛋白胨;而在以尿素、硫酸铵为氮源的培养基上生长缓慢。抗感菌株在不同氮源处理的培养基上生长状况一致,说明抗感菌株对不同氮源环境的适应性没有显著差异。
表7不同氮源条件下敏感菌株和抗性突变体的菌落直径
Table7Colonydiametersofsensitiveisolatesandresistantmutantsindifferentnitrogensources
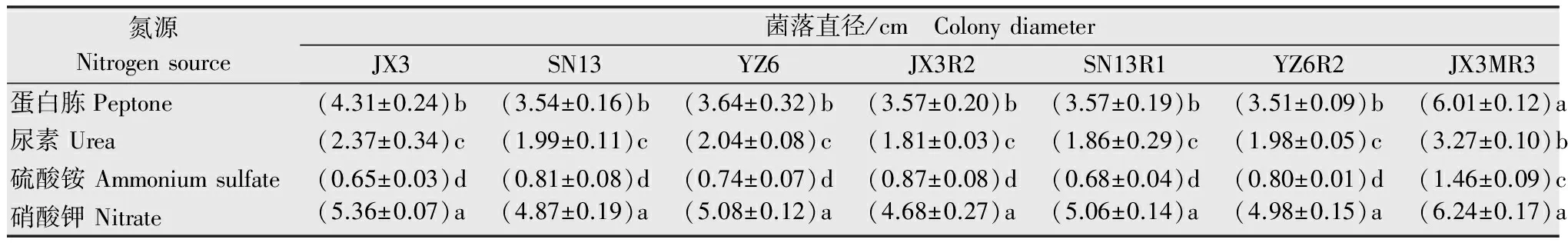
氮源Nitrogensource菌落直径/cm ColonydiameterJX3SN13YZ6JX3R2SN13R1YZ6R2JX3MR3YX6MR4蛋白胨Peptone(4.31±0.24)b(3.54±0.16)b(3.64±0.32)b(3.57±0.20)b(3.57±0.19)b(3.51±0.09)b(6.01±0.12)a(3.89±0.18)b尿素Urea(2.37±0.34)c(1.99±0.11)c(2.04±0.08)c(1.81±0.03)c(1.86±0.29)c(1.98±0.05)c(3.27±0.10)b(1.98±0.20)c硫酸铵Ammoniumsulfate(0.65±0.03)d(0.81±0.08)d(0.74±0.07)d(0.87±0.08)d(0.68±0.04)d(0.80±0.01)d(1.46±0.09)c(0.78±0.01)d硝酸钾Nitrate(5.36±0.07)a(4.87±0.19)a(5.08±0.12)a(4.68±0.27)a(5.06±0.14)a(4.98±0.15)a(6.24±0.17)a(5.25±0.15)a
2.4.5 抗感菌株在不同温度下的生长状况
试验结果(表8)表明,抗感菌株在15~35℃下均可生长,但最适温度均为25℃,与其他处理温度相比差异显著。在温度为15℃、35℃时,抗感菌丝生长速率均显著下降。
表8不同温度处理下抗感菌株的菌落直径差异
Table8Colonydiametersofsensitiveisolatesandresistantmutantsatdifferenttemperatures

温度/℃Temperature菌落直径/cm ColonydiameterJX3SN13YZ6JX3R2SN13R1YZ6R2JX3MR3YX6MR415(1.65±0.05)cd(1.43±0.18)d(1.83±0.17)c(1.40±0.10)cd(1.67±0.21)c(1.81±0.10)c(1.75±0.05)c(1.38±0.18)d20(3.48±0.05)b(3.19±0.12)b(3.23±0.04)b(3.25±0.03)b(3.20±0.18)b(3.65±0.18)b(4.02±0.12)b(3.86±0.19)b25(5.58±0.65)a(5.62±0.04)a(6.07±0.48)a(5.24±0.52)a(5.19±0.33)a(6.04±0.30)a(4.49±0.02)a(4.12±0.12)a30(1.97±0.05)c(1.73±0.08)c(1.79±0.04)c(1.74±0.21)c(1.59±0.39)cd(2.00±0.12)c(1.75±0.07)c(1.74±0.09)c35(1.32±0.01)d(1.19±0.05)e(1.22±0.01)d(1.15±0.10)d(1.15±0.13)d(1.36±0.14)d(1.17±0.05)d(1.20±0.04)d
3 结论与讨论
啶氧菌酯在中国被用于防治多种病害,但在山西省该药剂目前还未被广泛用于辣椒炭疽病的防治。本试验用菌丝生长速率法测定了从山西省晋中市3个未使用过QoI类杀菌剂的辣椒种植区分离的45株C.gloeosporioides对啶氧菌酯的敏感性。在啶氧菌酯及SHAM共同作用下,EC50+S平均值为(0.109±0.058)μg/mL,正态性检验显示敏感性分布呈近似正态分布,未出现敏感性明显下降的群体,其EC50+S平均值可作为C.gloeosporioides对啶氧菌酯的敏感基线,作为田间抗药性监测的参考标准。
由于QoI类杀菌剂独特的作用位点,一些病原菌可以通过旁路氧化补偿作用来降低对杀菌剂的敏感性[21]。旁路氧化途径可以在电子传递主路被阻断时,交替运行,以维持生物的活性。因此,病原菌体内所存在的旁路氧化补偿机制使离体条件下病原菌菌丝生长对QoI类杀菌剂敏感性下降。SHAM是旁路氧化途径中的交替氧化酶抑制剂,它可以阻断病菌体内的旁路氧化途径。在用菌丝生长速率法测定该类药剂敏感性时,加入SHAM可提高抑菌活性[22-23]。从SHAM+啶氧菌酯的联合毒力测定可知,其EC50+S的范围在0.022~0.275 μg/mL,旁路氧化的平均贡献值为78.026,说明旁路氧化途径在C.gloeosporioides菌丝生长过程中起着重要作用。陈聃所报道的葡萄炭疽病对吡唑醚菌酯旁路氧化平均贡献值为3.78[24],叶佳等报道的葡萄炭疽病菌对醚菌酯旁路氧化途径平均贡献值为20.09[11],明显低于本文中的78.026,推断旁路氧化作用在不同甲氧基丙烯酸酯类杀菌剂对不同病原菌中的贡献值存在较大差异。
QoI类杀菌剂通常被认为具有高的抗性风险[12,14,23], Zheng 等[25]和De Miccolis等[26]的研究也证实,在室内采用紫外线照射和药剂驯化的方式很容易得到抗性水平较高的抗性突变体。但低抗菌株抗药稳定性较差,推测可能是由于病原菌在药剂选择压下暂时表现出的耐药性反应,但在生产中有可能在持续用药的情况下,进一步突变为高抗菌株,加剧了病原菌产生抗性的风险。另外,突变位点不同的抗性突变体抗性水平和适合度差异很大,如F129L点突变表现为中等抗性,G143A点突变表现为高抗;又如Magnaporthegrisea和MycosphaerellagraminicolaG143A突变体与敏感菌株相比,适合度没有显著差异[19, 27],但G143S突变体的生长速率和产孢量与敏感菌株差异显著[28]。本研究比较了C.gloeosporioides抗感菌株的生长速率、产孢量,发现与敏感菌株相比,低抗突变体均没有显著差异。但在无药条件下,接种抗性突变体所产生的病斑面积以及侵染率明显低于敏感菌株,表明无药情况下,敏感菌株的致病能力强于抗性突变体;在药剂存在下,抗性突变体表现出一定的耐药性仍能使果实产生病斑面积。
抗感菌株在不同营养条件、温度、酸碱度下菌丝生长速率结果表明:在不同碳源条件下,抗感菌株均能正常生长,且淀粉为抗感菌株最佳碳源。在不同氮源条件下,抗感菌株均在以硝酸钾为氮源的培养基上生长最快,而在以硫酸铵为氮源的培养基中几乎不生长,说明抗感菌株对不同氮源适应性没有明显差别。在不同温度下,抗感菌株的最适温度均为25℃,温度偏高和偏低抗感菌株的菌丝生长速率均会下降。不同酸碱度下低抗菌株与敏感菌株菌丝生长状况差异不明显,但中抗菌株较敏感菌株对不同酸碱度的调节能力表现出一定优越性。总的来说,辣椒炭疽病菌C.gloeosporioides对啶氧菌酯产生抗性后,与敏感菌株相比,抗性突变体适合度没有明显下降,说明其对啶氧菌酯具有较高的抗性风险,因此,在生产中建议轮换使用不同作用机制的杀菌剂,使得C.gloeosporioides对啶氧菌酯始终保持低水平抗性频率,即能很大程度上延缓抗药性的产生。
今后,将进一步研究和比较Colletotrichum属不同种之间对啶氧菌酯敏感性的差异,探索Colletotrichum属中不同种的抗性突变体适合度及对啶氧菌酯的抗性风险。并针对高效药剂建立敏感基线,加强田间抗药性监测,根据抗性出现频率制定科学的用药策略,延长药剂的使用寿命。
[1] Lee J, Do J W, Yoon J B.Development of STS markers linked to major QTLs for resistance to the pepper anthracnose caused byColletotrichumacutatumandC.capsici[J]. Horticulture Environment and Biotechnology, 2011, 52(6): 596-601.
[2] Than P P, Jeewon R, Hyde K D, et al. Characterization and pathogenicity ofColletotrichumspecies associated with anthracnose on chilli (Capsicumspp.) in Thailand [J]. Plant Pathology, 2008, 57: 562-572.
[3] Kang B K, Min J Y, Kim Y S, et al. Semi-selective medium for monitoringColletotrichumacutatumcausing pepper anthracnose in the field [J]. Research of Plant Disease, 2005, 11: 21-27.
[4] Pakdeevaraporn P, Wasee S, Taylor P W J, et al. Inheritance of resistance to anthracnose caused byColletotrichumcapsiciin Capsicum [J]. Plant Breeding, 2005, 124: 206-208.
[5] 周传波,谢圣华,吉训聪,等.海南省辣椒炭疽病病原菌鉴定与防治试验[J].现代农业科技,2008(1):63-64.
[6] 李小霞,肖仲久. 贵州省辣椒炭疽病病原菌鉴定及室内毒力测定[J]. 广东农业科学, 2011, 44 (17): 55-57.
[7] Sanders G M, Korsten L, Wehner F C.Survey of fungicide sensitivity inColletotrichumgloeosporioidesfrom different avocado and mango production areas in South Africa [J]. European Journal of Plant Pathology, 2000, 106: 745-752.
[8] Maymon M, Zveibil L A, Pivonia S.Identification and characterization of benomyl-resistant and-sensitive populations ofColletotrichumgloeosporioidesfrom statice (Limoniumspp.)[J]. Phytopathology, 2006, 96: 542-548.
[9] 曹学仁,车海彦,杨毅,等.2014年海南省橡胶炭疽病菌对多菌灵和咪鲜胺的敏感性测定[J].植物病理学报,2015,45(6):626-631.
[10] 韩国兴,礼茜,孙飞洲,等.杭州地区草莓炭疽病病原鉴定及其对多菌灵和乙霉威的抗药性[J].浙江农业科学,2009,1(6):1169-1172.
[11] 叶佳, 张传清. 葡萄炭疽病菌对甲基硫菌灵、戊唑醇和醚菌酯的敏感性检测[J]. 农药学学报, 2012, 14(1): 111-114.
[12] Bartlett D W, Clough J M, Godwin J R, et al. The strobilurin fungicides [J]. Pest Management Science, 2002, 58: 649-662.
[13] Torriani S F F, Linde C C, Mcdonald B A.Sequence conservation in the mitochondrial cytochrome b gene and lack of G143A QoI resistance allele in a global sample ofRhynchosporiumsecalis[J]. Australasian Plant Pathology,2009,38:202-207.
[14] Gisi U, Chin K M, Knapova G, et al. Recent developments in elucidating modes of resistance to phenylamide, DMI and strobilurin fungicides [J]. Crop Protection, 2000, 19: 863-872.
[15] Miessner S, Stammler G.Monilinialaxa,M.fructigenaandM.fructicola: risk estimation of resistance to QoI fungicides and identification of species with cytochromebgene sequences [J]. Journal of Plant Diseases and Protection,2010,117(4):162-167.
[16] Markoglou A N, Malandrakis A A, Vitoratos A G, et al. Characterization of laboratory mutants ofBotrytiscinerearesistant to QoI fungicides [J]. European Journal of Plant Pathology, 2006, 115: 149-162.
[17] Pasche J S, Piche L M, Gudmestad N C.Effect of the F129L mutation inAlternariasolanion fungicides affecting mitochondrial respiration [J]. Plant Disease, 2005, 89: 269-278.
[18] Rebollar-Alviter A, Madden L V, Jeffers S N, et al. Baseline and differential sensitivity to two Qo I fungicides among isolates ofPhytophthoracactorumthat cause leather rot and crown rot on strawberry[J]. Plant Disease, 2007, 91(12): 1625-1637.
[19] Zhou Yuxin, Chen Lei, Hu Jian, et al. Baseline sensitivity of natural population and resistance risk ofPeronophythoralitchiito four novel QoI fungicides [J]. European Journal of Plant Pathology, 2016, 146: 71-83.
[20] Zhao Xiaojun, Ren Lu, Yin Hui, et al. Sensitivity ofPseudoperonosporacubensisto metalaxyl, dimethomorph and fosetyl-aluminium in Shanxi of China [J].Crop Protection,2013,43(1):38-44.
[21] Ma Zhonghua, Michailieds T J. Advances in understanding molecular mechanisms of fungicide resistance and molecular detection of resistant genotypes in phytopathogenic fungi [J]. Crop Protection, 2005, 24(10): 853-863.
[22] Kraiczy P, Haase U, Gencic S, et al. The molecular basis for the natural resistance of the cytochromebc1 complex from strobilurin-producing basidiomycetes to center Qp inhibitors[J]. European Journal of Biochemistry, 1996, 235: 54-63.
[23] Fernadez-ortuno D, Tores J A, De Vicente A,et al. Mechanisms of resistance to qol fungicides in phytopathogenic fungi[J]. International Microbiology, 2008, 11(1): 1-9.
[24] 陈聃. 葡萄炭疽病菌的抗药性检测和治理研究[D]. 临安: 浙江农林大学, 2013.
[25] Zheng Desen, Olaya G, Koller W. Characterization of laboratory mutants ofVenturiainaequalisresistant to strobilurin-related fungicide kresoxim-methyl[J]. Current Genetics, 2000, 38(3): 148-155.
[26] De Miccolis A R M, Rotolo C, Masiello M, et al. Genetic analysis and molecular characterisation of laboratory and field mutants ofBotryotiniafuckeliana(Botrytiscinerea) resistant to Qo I fungicides[J]. Pest Management Science, 2012, 68(9): 1231-1240.
[27] Siah A, Deweer C, Morand E, et al. Azoxystrobin resistance of FrenchMycosphaerellagraminicolastrains assessed by fourinvitrobioassays and by screening of G143A substitution [J]. Crop Protection, 2010, 29: 737-743.
[28] Avila-adame C, Koller W. Characterization of spontaneous mutants ofMagnaporthegriseaexpressing stable resistance to the Qo-inhibiting fungicide azoxystrobin [J]. Current Genetics, 2003, 42(6): 332-338.
(责任编辑: 田 喆)
BaselinesensitivityofColletotrichumgloeosporioidestopicoxystrobinandbiologicalcharacteristicsofresistantmutants
Ren Lu1,2, Zhou Jianbo1,3, Liu Huiping2, Cao Junyu2, Yin Hui1,3, Zhao Xiaojun1,3
(1.ShanxiKeyLaboratoryofIntegratedPestManagementinAgriculture,Taiyuan030031,China; 2.CollegeofAgriculture,ShanxiAgriculturalUniversity,Taigu030801,China; 3.InstituteofPlantProtection,ShanxiAcademyofAgriculturalSciences,Taiyuan030006,China)
To evaluate the resistance risk ofColletotrichumgloeosporioidesto picoxystrobin, the baseline sensitivity ofC.gloeosporioidesto this fungicide was established, and the biological characteristics of the picoxystrobin-resistant mutants obtained in the laboratory were studied. Forty-fiveC.gloeosporioidesisolates were collected from three areas of Jinzhong, Shanxi Province, where picoxystrobin and the other fungicides with the same mode of action had not been used before. The sensitivities of 45C.gloeosporioidesto picoxystrobin were determined by growth rate method. The results showed that the EC50values ranged from 1.404 μg/mL to 16.650 μg/mL, with a mean of (6.783±3.499)μg/mL. The EC50+Svalues of 45C.gloeosporioidesto picoxystrobin combined with 100 μg/mL salicylic acid oxime (SHAM) ranged from 0.022 μg/mL to 0.275 μg/mL, with a mean of (0.109±0.058)μg/mL. The frequency of EC50+Svalues was distributed as a unimodal curve, and the frequency of sensitivity was in approximately normal distribution, and the mean EC50+Svalues could be used as the relative baseline sensitivity ofC.gloeosporioidesto picoxystrobin. The compensation efficiency value in the bypass oxidation way showed that the lowest value was 6.039, and the highest was 301.441, with a mean value of 78.026. Eight picoxystrobin-resistant mutants were obtained, of which 6 were low resistance and 2 medium resistance to picoxystrobin. The resistance of moderately resistant mutants was stable after 10 generations of asexual cultivation. The mycelium growth rates, spore germination rates of mutants were similar with those of sensitive isolates, but the pathogenicity of the mutants was lower than that of the sensitive ones in absence of fungicides. The best carbon source of both sensitive and resistant isolates was starch and best nitrogen source was potassium nitrate; the optimum pH values were 5 and 6 and the optimum temperature was 25℃ for the mycelial growth of both sensitive and resistant isolates. The results showed that theC.gloeosporioidesresistant mutants were in high fitness and in high risk of resistance to picoxystrobin. The conclusion provides the theoretical basis for guiding fungicide use and the strategic buildup of delaying resistance development.
Colletotrichumgloeosporioides; picoxystrobin; baseline sensitivity; resistance risk; biological characteristics
2016-12-21
2017-02-04
山西省重点研发计划重点项目(201603D21110-2);农业有害生物综合治理山西省重点实验室开放课题(YHSW2015002);山西省农业科学院特色农业技术攻关项目(YGG17114);山西省农科院重点攻关项目(YGG1603)
* 通信作者 E-mail:zhaoxiaojun0218@163.com
S 481.4
A
10.3969/j.issn.0529-1542.2017.06.005
猜你喜欢
当代水产(2022年7期)2022-09-20
河北科技师范学院学报(2022年2期)2022-08-26
亚热带农业研究(2022年1期)2022-08-08
方圆(2021年19期)2021-11-03
中国饲料(2021年17期)2021-11-02
空间科学学报(2021年1期)2021-05-22
农业科技通讯(2021年1期)2021-03-06
小学生必读(中年级版)(2021年12期)2021-03-03
小学生必读(中年级版)(2021年11期)2021-02-22
中国农业科技导报(2020年3期)2020-03-15
