
黔东南州篦子三尖杉群落结构特征研究
2017-11-24 08:22冯邦贤韦海霞
湖南林业科技 2017年4期
冯邦贤,韦海霞
(1. 铜仁市林业局,铜仁 554300; 2. 铜仁学生物与农林工程学院,铜仁 554300)
黔东南州篦子三尖杉群落结构特征研究
冯邦贤1,韦海霞2
(1. 铜仁市林业局,铜仁 554300; 2. 铜仁学生物与农林工程学院,铜仁 554300)
篦子三尖杉为国家Ⅱ级保护植物。为探索篦子三尖杉不同群落类型之间物种多样性的规律,比较群落结构和物种多样性的差异性,采用样方调查方法分析了黔东南州凯里市、镇远县、黎平县和台江县的 4 个篦子三尖杉群落的物种组成及群落结构。结果表明:4 个篦子三尖杉群落共有维管植物 54 科 87 属 102 种,其中,蕨类植物 5 科、裸子植物 2 科、被子植物 47 科;4 个群落中高位芽植物所占比例最大,一年生植物所占的比例最小;4 个篦子三尖杉群落的成层现象均非常明显,基本上可以分为乔木层、灌木层和草本层 3 个层次,篦子三尖杉是灌木层中的优势种;受人为活动的影响,4 个群落物种多样性水平表现出明显的差异性,物种多样性水平从高到低的顺序是:台江>凯里>镇远>黎平;篦子三尖杉缺乏成年个体和幼苗,种群属于衰退型。
篦子三尖杉;群落结构;黔东南州
篦子三尖杉(Cephalotaxus oliveri)隶属于三尖杉科(Cephalotaxaceae)三尖杉属(Cephalotaxus)常绿灌木。叶条形,螺旋状着生,平展成两列,中脉稍明显或仅中下部明显,下面具白色气孔带。花期 4 月,种子次年 10 月成熟。分布于四川、云南、湖北、湖南、广东、江西等省份,也分布到越南北部和老挝部分地区,在贵州产梵净山、镇远、黎平、榕江等地[1-5],为国家二级保护植物。有关篦子三尖杉的报道主要包括繁育技术[6-7]、资源与地理分布现状[8]、生物学特征与保护[9-11]、形态解剖[12]、胚胎发育[13]、化学成分与分类学研究[14]、分子生物学与系统进化研究[15-17],植株中所含的三尖杉酯碱的抗氧化活性进研究[18]等,对篦子三尖杉群落物种组成及结构的研究相对较少,仅有艾启芳等对我国近年来在篦子三尖杉的保护生物学、群落结构、遗传多样性、繁殖、保护等方面的研究进行了综述[19],黎桂芳对江西武功山篦子三尖杉野生资源的分布现状、生存条件及群落特性进行了调查研究[20]。贵州省有关篦子三尖杉群落物种组成及结构的研究都出自各个自然保护区的科学考察集中,如黎平太平山自然保护区、南宫自然保护区、老蛇冲自然保护区等考察集中都有对篦子三尖杉的资源分布、群落特征等方面的研究[21-23]。本文研究旨在探索不同群落类型之间物种多样性的规律,比较群落结构和物种多样性的差异,为珍稀植物群落物种多样性保护和持续利用。
1 研究区概况
黔东南苗族侗族自治州地处贵州省东南部,地理坐标为 107°17'20''~109°35'24'' E,25°19'20''~27°31'40'' N,州内东西宽 220 km,南北长 240 km,总面积 30 337 km2。全州地跨扬子准台地与华南褶皱带两个一级大地构造单元,处于云贵高原向湘桂丘陵盆地过渡地带,地形地貌复杂。州境总体地势是北、西、南三面高而东部低。中部雷公山区和南部月亮山为中山地带,西部和西北部为丘陵状低中山区,东部和东南部下低中山、低山、丘陵、盆地。境内大部分地区海拔 500~1 000 m。最高点为雷公山主峰黄羊山,海拔 2 178.8 m,最低点为黎平县地坪乡井郎村水口河出省处,海拔137 m。年平均气温 14~18 ℃,降雨量 1 000~1 500 mm,相对湿度为 78%~84%,无霜期 270~330 天。属中亚热带季风湿润气候区,具有冬无严寒、夏无酷暑、雨热同季的特点。独特的地理位置和复杂的自然环境,孕育了丰富的生物多样性,使黔东南州成为贵州生物多样性的关键地区之一。植被类型多样,地带性植被为各类常绿阔叶林,局部有常绿落叶阔叶混交林,非地带性植被以人工马尾松、杉木林为主。组成种类繁多,以壳斗科、樟科、山茶科、木兰科等植物为主,更有不少古老植物和珍稀植物在此得以幸存。篦子三尖杉各样地的基本情况见表 1。

表1 篦子三尖杉不同样方基本情况Tab.1 Environmental conditions of different simples of Cephalotaxus oliveri
2 研究方法
2.1 样地设置
分别于 2016 年 7 月、9 月和 10 月对黔东南州凯里市、镇远县、黎平县和台江县的篦子三尖杉群落进行了调查,均设置了总面积为 1 200 m2的样地。样地由 2 个面积为 20 m×20 m 的样方及 2个面积为 30 m×20 m 的样方组成,并采用相邻格子法,将 20 m×20 m 的样方分割成 8 个 5 m×10 m的小样方,将 30 m×20 m 的样方分割成 12 个 5 m×10 m 的小样方(共计 40 个)。在每个小样方内,测量和记录每株胸径≥5 cm 的乔木种类、胸径、树高及冠幅,所有灌木的种类、株树、平均地径、平均高度以及盖度,所有草本植物的种类和多度。
2.2 多样性指数计算方法
参考《植被生态学》中常用物种多样性指数的公式[24]。
式中:Pi为物种个体属于第 I 种的概率,以其个体占总个体数的分数表示;C 为常数,一般设置C=1。
物种均匀度指数(J),采用 Pielon 均匀度指数:实测多样性(H)与最大多样性(Hmax)之间的比率:

2.3 生活型
根据 C.Raunkiaer 的方法对群落中各物种的生活型及叶特征进行分类[25]。
2.4 数据处理
采用 EstimateS 820 Win 计算物种的多样性指数,Origin7.5 进行作图分析,其它的计算在 Excel工作表中完成。
3 结果与分析
3.1 群落物种组成
通过调查统计,4 个篦子三尖杉群落共有维管植物 54 科 87 属 102 种(含变种,下同)。其中,蕨类植物 5 科 6 属 6 种,裸子植物 2 科 2 属2 种,被子植物 47 科 79 属 94 种。由于经纬度以及海拔高度的变化不大,篦子三尖杉所在群落类型基本相同,群落的种类组成和优势种类也大体一致。群落的乔木层物种组成及重要值见表 2,从乔木层的分析来看,4 个群落的乔木层分别有树种8 种、9 种、18 种和 13 种,枫香是每个群落中都出现的乔木树种,且重要值都排在前 4 位,在凯里和台江的两个群落中,其重要值均居第一位,值分别达到 92.74 和 63.12,在乔木层中占绝对优势。

表2 篦子三尖杉群落乔木层物种重要值Tab.2 Importance values in tree layer of Cephalotaxus oliveri community
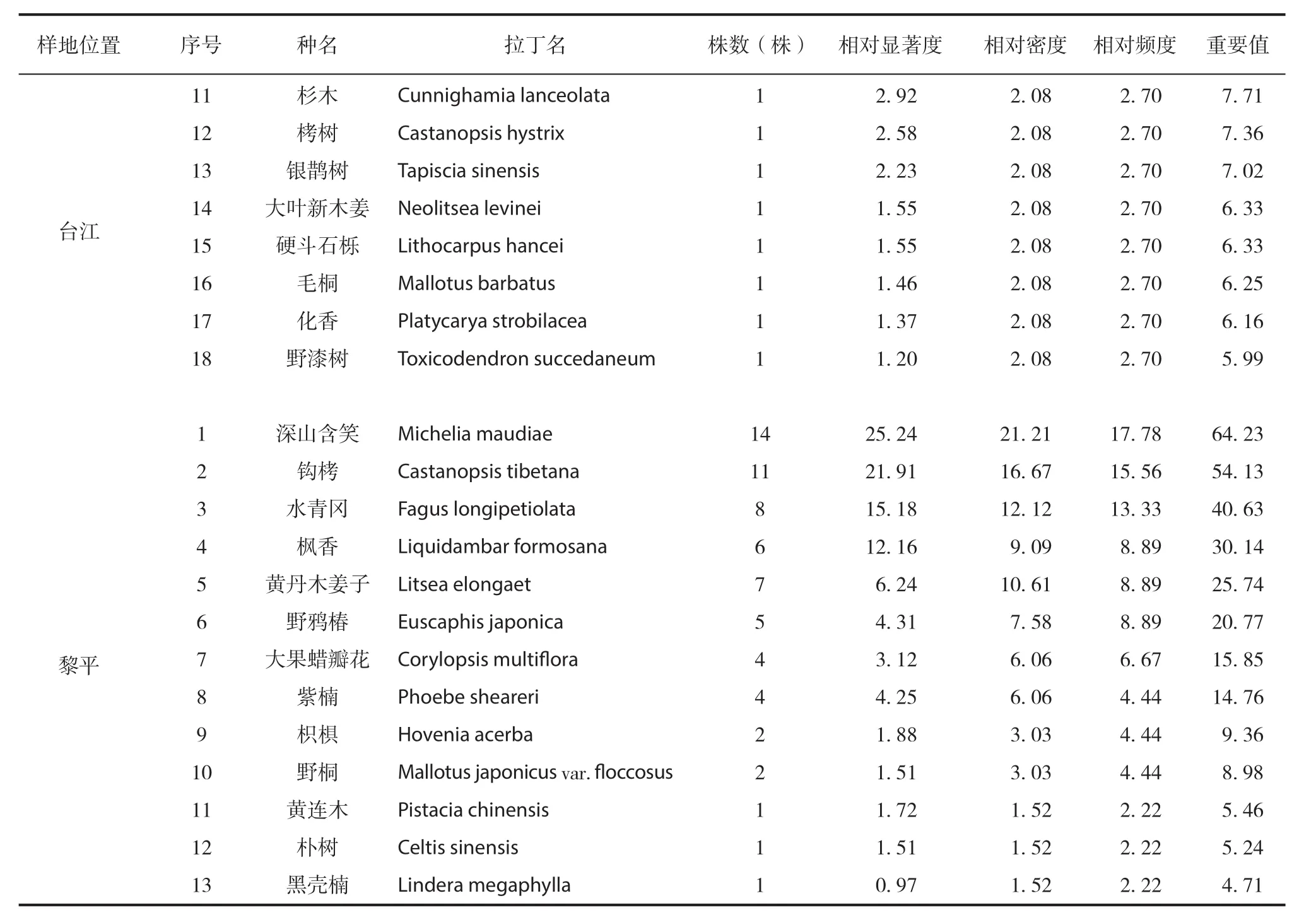
续表2 篦子三尖杉群落乔木层物种重要值Continued Tab.2 Importance values in tree layer of Cephalotaxus oliveri community
灌木层的物种组成及数量等特征见表 3,从表 3 中可知,篦子三尖杉在每个群落中数量均较多,是灌木层的优势种。

表3 篦子三尖杉群落灌木层种类组成Tab.3 Composition of species in shrub layer of Cephalotaxus oliveri community

续表3 篦子三尖杉群落灌木层种类组成Continued Tab.3 Composition of species in shrub layer of Cephalotaxus oliveri community
草本层及层外植物层的物种组成见表 4,相对于乔木层和灌木层,草本层物种多样性更简单,层外植物层物种数则更少。

续表4 篦子三尖杉群落藤本植物和草本植物组成Continued Tab.4 Composition of liana and herbaceous plant of Cephalotaxus oliveri community
3.2 群落外貌特征及生活型
植被生活型是植物群落对其所处环境适应状况的综合反映,是植物与环境长期相互作用的结果,植物在适应环境过程中,不仅形成了与环境条件相适应的生活史对策,而且也表现出特定的生活型外貌。群落的外貌是认识植物群落的基础,也是区分不同植被类型的主要标志。黔东南州 4 个篦子三尖杉群落外貌呈绿色,较整齐,乔木树种落叶成分和常绿成分所占比例相差不大。按照 C.Raunkiaer 对植物生活型的分类系统,对 4个群落组成种类综合划分生活型类型,即:高位芽植物(Ph.)、地上芽植物(Ch.)、地面芽植物(H.)、隐芽植物(Cr.)、一年生植物(Th.)。群落生活型谱见图 1。各个生活型所占比例依次为 64.7%、13.7%、6.9%、11.8%、2.9%。从图 1 中可以看出,高位芽植物所占的比例最大,一年生植物所占的比例最小。群落物种以高位芽植物为主,说明篦子三尖杉群落的性质和我国典型亚热带常绿阔叶林相似。

图1 篦子三尖杉群落物种生活型谱Fig.1 Life form spectrum of Cephalotaxus oliveri community
注:Ph.为空位芽植物;Ch.为地上芽植物;H.为地面芽植物;Cr.为隐芽植物;Th.为一年生植物。
3.3 群落物种多样性
物种多样性是反映群落物种数量及分布均匀程度的综合指标。通过对 4 个篦子三尖杉群落物种多样性指标的计算,得到各个群落中的物种丰富度指数、Simpson 指数、Shannon-Wienner 指数和 Pielou 均匀度指数(见表 5),从表 5 中可以看出,4 个篦子三尖杉群落的物种丰富度均较低,三个样地的值在 30 以下。Simpson 指数的值为 0.825 5~0.902 3,Shannon-Wienner 指数的值为 2.266 0~2.729 5,Pielou 均匀度指数的值为0.344 9~0.413 6,所有物种多样性指数均是台江的篦子三尖杉群落最高,物种丰富度最为明显,值达到 40。
3.4 群落结构
3.4.1 群落的垂直结构 垂直结构是反映群落结构重要的因素之一,群落的垂直结构决定于各种植物种的高度及在不同高度植物的数量。植株的高度级概率分布反映了树种在森林中的各种特征(如耐荫性、更新类型等)以及群落的分层结构。黔东南州 4 个篦子三尖杉群落的层次结构均较为简单,可分为乔木层、灌木层、草本层,凯里、台江、黎平的 3 个群落还存在层间植物层,但种类和数量均不多,高度主要集中在 5~7 m。乔木层高度集中的 9~18 m,主要由枫香、构树、海通、深山含笑、南酸枣、钩栲等组成。灌木层高度集中在 1~3 m,种类和数量相对来说均较多,其中一些种类是乔木层的幼苗,出现株树较多的植物有篦子三尖杉、山胡椒、檵木、细枝柃、木姜子等。篦子三尖杉是灌木层中的优势种,平均高度在 1.2~1.6 m,冠幅 0.2~0.3 m,地径 1.5~1.8 cm。草本植物高度多在 0.5 m 以下,主要有异叶楼梯草、序叶苎麻、翠云草、五节芒等组成。层间植物主要为木本植物,以爬藤榕、钩藤、冷饭团为主,多攀附在灌木植物上,个别植物可达到乔木层。
根据树高和植株数量作出了4个篦子三尖杉群落木本植物的高度结构图,由图 2 得:4 个群落的高度结构大致相同,除黎平的群落外,1≤h≤5 m,5 m≤h≤10 m,10 m≤h≤15 m,15 m≤h≤20 m,h>20 m 高度级的个体数依此减少。1≤h≤5 m 高度级的个体数目在 4 个群落中都占有很大的比例。
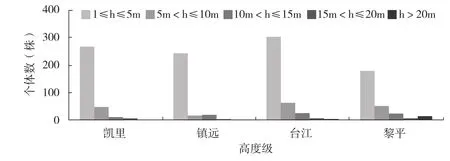
图2 篦子三尖杉群落木本植物高度结构Fig.2 The height structure of woody plants of 4 Cephalotaxus oliveri communities
3.4.2 群落中篦子三尖杉种群年龄结构 分析种群的年龄结构是探索种群动态的有效方法,在研究中常采用径级或高度近似代替年龄。本文采用高度代替年龄对篦子三尖杉种群的年龄结构进行分析。图 3 为 4 个样地中篦子三尖杉种群年龄结构图,从图 3 中可以看出,4 个篦子三尖杉种群年龄结构相似,按年龄的分布均没有没有明显的规律,从低年龄往高年龄不呈金字塔形。h≤0.5 m 的幼苗稀少,分布在凯里的群落中的数量更是为 0,主要集中在 1~2 m,数量达到 70%以上。

表5 各样地物种多样性指数分析Tab.5 The analysis of species diversity indexes in different simple
从整体调查的结果来看,4 个样方中的篦子三尖杉在一段时间内仍然是群落灌木层中的优势种,但由于缺乏幼苗的更新,群落中的成年个体也较少,从结实这一表现看,结实植株也比较少,地上也没有发现种子,随时间的推移篦子三尖杉种群会趋向衰退阶段。

图3 篦子三尖杉种群的高度结构Fig.3 The height structure of 4 Cephalotaxus oliveri communities
4 结论与讨论
黔东南州 4 个篦子三尖杉群落共有维管植物 54 科 87 属 102 种。其中,蕨类植物 5 科 6 属6 种,裸子植物 2 科 2 属 2 种,被子植物 47 科 79属 94 种,4 各群落的群落类型基本相同,群落的种类组成和优势种类也大体一致。篦子三尖杉在每个群落中数量均较多,是灌木层的优势种,这与郎学东等对云南、贵州、江西分布的 8 个篦子三尖杉群落物种组成相似[26]。4 个篦子三尖杉群落外貌呈绿色,较整齐,乔木树种落叶成分和常绿成分所占比例相差不大。生活型中高位芽植物所占的比例最大,一年生植物所占的比例最小。群落物种以高位芽植物为主,说明篦子三尖杉群落的性质和我国典型亚热带常绿阔叶林相似。对4个篦子三尖杉群落物种多样性指标的计算,得到各个群落中的物种丰富度指数、Simpson 指数、Shannon-Wienner 指数和 Pielou 均匀度指数。各指数计算结果表明,4 个篦子三尖杉群落的物种丰富度均较低,三个样地的值在 30 以下。Simpson指数的值为 0.825 5~0.902 3,Shannon-Wienner 指数的值为 2.266 0~2.729 5,Pielou 均匀度指数的值为 0.344 9~0.413 6,所有物种多样性指数均是台江的篦子三尖杉群落最高,物种丰富度最为明显,值达到 40。4 个群落的层次结构均较为简单,可分为乔木层、灌木层、草本层,凯里、台江、黎平的 3 个群落还存在层间植物层,但种类和数量均不多,高度主要集中在 5~7 m。乔木层高度集中的 9~18 m,灌木层高度集中在 1~3 m,篦子三尖杉是灌木层中的优势种,草本植物高度多在 0.5 m 以下。4 个篦子三尖杉种群年龄结构相似,按年龄的分布均没有没有明显的规律,从低年龄往高年龄不呈金字塔形。4 个样方中的篦子三尖杉在一段时间内仍然是群落灌木层中的优势种,但由于缺乏幼苗的更新,群落中的成年个体也较少,从结实这一表现看,结实植株也比较少,地上也没有发现种子,随时间的推移篦子三尖杉种群会趋向衰退阶段,由于受人为活动频繁,这与黎平太平山自然保护区、南宫自然保护区、老蛇冲自然保护区等考察集中篦子三尖杉种群不同[21-23]。
[1]郑万钧,傅立国.中国植物志:第七卷[M].北京:科学出版社,1978:434-436.
[2]陈谦海.贵州植物志:第一卷[M].贵阳:贵州人民出版社,1982:32-33.
[3]Fu Liguo,Li Nan,Robert R Mill.Flora of China[M].Beijing:Science Press,1999:85-88.
[4]Farjon,Aljos.World Checklist and Bibliography of Conifers[M].Richmond.UK.:Royal Botanical Gardens.Kew.1998.
[5]Mark Newman,Sounthone Ketphanh,Bouakhaykhone Svengsuksa,et al.A Checklist of the Vascular Plants of Lao PDR[M].UK:Royal Botanic Garden Edinburgh:2007,34.
[6]吴朝斌,伍铭凯,杨汉远,等.篦子三尖杉育苗技术[J].育苗技术,2007(8):23.
[7]吕 梅,王瑞辉,曹基武,等.篦子三尖杉愈伤组织的诱导[J].湖南林业科技,2010,37(1):7-9.
[8]司马永康,于 鸿,杨桂英,等.云南省三尖杉属植物的地理分布与环境因子的关系[J].林业调查规划,2004,29(1):83-87.
[9]Mitsuo Suzuki.The Course of Resin Canals in the Shoots of ConifersⅠ.Taxaceae,Cephalotaxaceae and Podocarpaceae[J].Bot Mag Tokyo:1979(92):235-251.
[10]顾志建,周其兴,岳中枢.三尖杉科的核形态学研究[J].植物分类学报,1998,36(1):47-52.
[11]郭泉水,包奋强,王祥福,等.三尖杉属所属群落优势乔木树种种间关系[J].林业科学研究,2008,21(5):662-668.
[12]席以珍.三尖杉科的花粉形态及其外壁超微结构研究[J].植物分类学报,1993,31(5):925-931.
[13]李 荣,土伏雄,陈祖铿.篦子三尖杉的胚胎学研究及其系统位置探讨[J].植物分类学报,1986,24(6):411-422.
[14]马忠武,保关福.三尖杉属(科)植物的双黄酮成分及其化学分类的研究[J].植物分类学报,1990,28(2):48-53.[15]Yuchang Cheng,Robert G,Nicolson,et al.Phylogeny of Taxaceae and Cephalotaxaceae Genera Inferred from Chloroplast matK Gene and Nuclear rDNA ITS Region[J].Molecular Phylogenetics and Evolution,2000,14(3):353-365.
[16]Da Cheng Hao,Pei Gen Xiao,BeiLi Huang,et al.Interspecific relationships and Origins of Taxaceae and Cephalotaxaceae revealed by partitioned Bayesian analyses of Chloroplast and nuclear DNA sequences[J].Plant Syst Evol,2008(276):89-104.
[17]应站明,杨明照,苏应鹃.篦子三尖杉 IAAR-PCR 系统优化[J].南昌大学学报:理科版,2009,33(3):298-301.
[18]Zeng Lingbin,Lilian,L.P.Vrijmoed.Antioxidant activities and phenolic constituents of Cephalotaxus oliveri Mast.Aerial parts[J].Journal of the Serbian Chemical Society,2012,77(4):437-451.
[19]艾启芳,陈明慧,梁 娴.篦子三尖杉的研究进展[J].贵州农业科学,2010,38(3):181-183.
[20]黎桂芳.江西武功山篦子三尖杉资源的调查研究[J].萍乡高等专科学校学报,2005(4):48-50.
[21]张华海,李明晶,邓锦光.黎平太平山自然保护区综合科学考察集[M].贵阳:贵州科技出版社,2006.
[22]张华海.老蛇冲自然保护区科学考察集[M].贵阳:贵州科技出版社,2003.
[23]张华海.南宫自然保护区科学考察集[M].贵阳:贵州科技出版社,2003.
[24]宋永昌.植被生态学[M] .上海:华东师范大学出版社,2001.
[25]曲仲湘,吴玉树,王焕校,等.植物生态学(第二版)[M].北京:高等教育出版社,1984:2-155.
[26]郎学东,苏建荣,张志钧,等.濒危植物篦子三尖杉的群落特征[J].林业科学研究,2011,24(6):727-735.
(文字编校:杨 骏)
Community structure characteristics of Cephalotaxus oliveri in the Qiandongnan Miao and Dong Autonomous Prefecture
FENG Bangxian1,WEI Haixia2
(1. Forestry Bureau of Tongren City,Tongren 554300,China;
2. College of Biology and Engineering,Tongren University,Tongren 554300,China)
Cephalotaxus oliveri is the second class conservative plant of China,and it belongs to Cephalotaxus of Cephalotaxaceae.The species composition and structural characteristics of C.oliveri grows in Kaili,Zhenyuan,Liping and Taijiang County of the Qiandongnan Miao and Dong Autonomous Prefecture were studied with plot investigation.The results showed that:Totally,102 species of plants were recorded in the 4 C.oliveri communities,and they were distributed into 54 families and 87 genera.That concluded 5 family pteridophyte,2 families gymnosperms and 47 families angiosperms.In the 4 C.oliveri communities,the proportion of phanerophytes is the largest,and the proportion of therophytes is the least.The community structure of the 4 C.oliveri communities has a characteristic of obvious stratification phenomenon and simple structure,the community is basically divided into tree layer,shrub layer and herb layer three levels,and C.oliveri is the dominant species in the shrub layer.The complexity of the diversity from high to low is Taijiang>Kaili>Zhenyuan>Liping.4 C.oliveri communities lacks of adult and seeding, the population belongs to declined type.
Cephalotaxus oliveri;community structure;Qiandongnan Miao and Dong Autonomous Prefecture
2017-03-21
“黔东南森林植物种质资源调查”项目
冯邦贤(1984-),男,贵州省织金县人,工程师,主要从事种质资源、种子植物分类及森林生态学研究;E-mail:bangxianfeng@126.com
S 718.54
A
1003-5710(2017)04-0034 -09
10.3969 / j.issn. 1003-5710.2017.04.008
猜你喜欢
建筑与预算(2022年6期)2022-07-08
祝您健康·文摘版(2022年4期)2022-04-02
水利水运工程学报(2021年5期)2021-11-10
农村实用技术(2020年1期)2020-12-16
山西建筑(2019年10期)2019-04-01
乡村科技(2018年8期)2018-02-13
现代园艺(2018年14期)2018-01-18
小猕猴智力画刊(2017年11期)2017-12-07
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
