
陕西省鼠疫耶尔森菌差异区段分型及其流行病学特征分析
2017-11-21 03:23:53安翠红陈宝宝聂守民范锁平孙养信
中国人兽共患病学报 2017年10期
安翠红,陈宝宝,吕 文,聂守民,范锁平,孙养信
陕西省鼠疫耶尔森菌差异区段分型及其流行病学特征分析
安翠红,陈宝宝,吕 文,聂守民,范锁平,孙养信
目的分析陕西省鼠疫耶尔森菌差异区段分型及流行病学特征。方法对分离自陕西省鼠疫疫区获得的48株鼠疫菌应用23个DFR(差异区段)和PMT1(质粒验证)PCR扩增,进行鼠疫菌DFR基因组分型及流行病学特征分析。结果相同年份,不同染疫动物、媒介蚤分离的鼠疫菌基因组型基本一致。陕西鼠疫耶尔森菌DFR基因组型有Genomovar11、17、20共3种。1987-1988年份、2000-2001年份DFR基因组型以Genomovar17为主,2006年份以Genomovar20为主。结论不同年份疫情主导的基因组型不同,随着时间的变化,鼠疫菌DFR基因组型从Genomovar17基因缺失微进化到Genomovar20。
陕西;鼠疫菌;DFR分型;流行病学
鼠疫是一种自然疫源性疾病,其病原体为鼠疫耶尔森氏菌(以下简称鼠疫菌)。根据对甘油、亚硝酸盐和阿拉伯糖的代谢能力,将鼠疫菌分为古典、中世纪、东方和田鼠4个生物变种[1]。1987年纪树立等将鼠疫菌分为17个生态型[2]。1997年和2005年青藏高原青海田鼠和准格尔盆地大沙鼠疫源地的发现,丰富了我国鼠疫菌生态型,其数目增加到21个[1]。21个生态型鼠疫菌各有特定的地理位置,对人的侵袭力、致病力各不相同。差异区段分析(Different region,DFR)是指在一些菌株基因组中存在,而在另外一些菌株的基因组中缺失的基因区[3],DFR的缺失和获得反映了鼠疫菌在自然界适应性演化的遗传基础[4]。周冬生和Li 等[5-6]将DFR应用于鼠疫菌的分子分型,提出了主要基因组型和次要基因组型的概念,主要基因组型与鼠疫菌在自然疫源地中的演化密切相关。
陕西省榆林市定边县1987年5月被判定为鼠疫疫源地,为鄂尔多斯高原长爪沙鼠鼠疫疫源地的一部分。1987-1988年、2000-2001年、2006年发生3次动物鼠疫流行,鼠疫菌生物变种为中世纪变种,生态型为鄂尔多斯高原型。为探究鼠疫菌在陕西省鼠疫疫源地的进化及其与生态适应的关系,本文运用23个差异区段和PMT1对陕西鼠疫疫源地定边县三起动物鼠疫流行期间分离到的48株鼠疫菌进行DFR基因组分型,并分析流行特征。
1 菌株来源
三起鼠间鼠疫流行期间分离到48株菌。其中1987-1988年8株,2000-2001年30株,2006年10株。菌株分离于自毙动物及自毙动物寄生蚤,其中长爪沙鼠35株,黑线仓鼠1株,荒漠毛足鼠1株,五趾跳鼠1株,秃病蚤5株,同型客蚤4株,二齿新蚤1株。
2 方 法
2.1鼠疫菌DNA的提取 DNA提取按照文献[7]进行。
2.2DFR分型 用23对分型引物DFR01~DFR23和质粒验证引物PMT1对被试菌株DNA进行分型,实验设置阴性、阳性对照,以91001、620024菌株DNA的等量混合物为阳性对照,结果分析按照文献[6]进行。
3 结 果
采用PCR技术对分离48株鼠疫菌23个DFR和PMT1进行了验证,获得了每株菌的DFR图谱,共发现DFR基因组型有3种,分别为11、17、20。见表1。1987-1988年份、2000-2001年份DFR基因组型以17为主,2006年份以20为主。相同年份,不同染疫动物、媒介蚤分离的鼠疫菌基因组型基本一致。不同年份、不同宿主、媒介来源菌株基因组型见表2、表3。疫源乡镇分布见图1。
表1 48株鼠疫菌的DFR基因组型
Tab.1 Genomovars of 48 Yersinia pestis based on DFR profiling

基因组型Genomovars菌株数numbersofstrainsDFR位点Locus1234567891011121314151617181920212223PMT1Genomovar114011110011111010001111101Genomovar1733011110011111010000111101Genomovar2011011110011110010000111101
注:表头中1~23代表DFR1~DFR23位点.“1”表示基因组组中存在该DFR;“0”表示该DFR在基因组组中缺失。
Note: the 1~23 in the header represents the DFR1~DFR23 locus. The number 1 and 0 indicate the presence and absence of the DFR locus in the genome.
表2 不同年份鼠疫菌分离株的DFR基因组型
Tab.2 Genomovars of Yersinia pestis isolates from different years
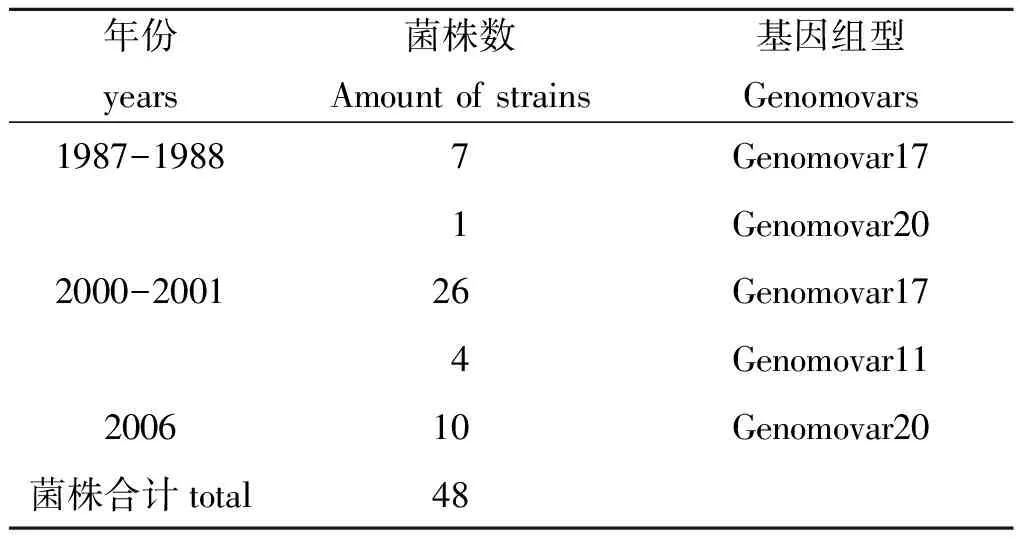
年份years菌株数Amountofstrains基因组型Genomovars1987-19887Genomovar171Genomovar202000-200126Genomovar174Genomovar11200610Genomovar20菌株合计total48

图1 定边县疫源乡镇分布Fig.1 Epidemic villages and towns in Dingbian County
表3 不同宿主、媒介来源菌株的DFR基因组型
Tab.3 DFR genomovar of Yersinia pestis isolates from different host and media

年份years宿主动物hosts媒介vector菌株数No.ofstrains基因组型Genomovars1987-1988长爪沙鼠Merionesunguiculatus4Genomovar17秃病蚤蒙冀亚种Nosopsylluslaevicepskuzenkovi1Genomovar20秃病蚤蒙冀亚种N.laevicepskuzenkovi1Genomovar17同形客蚤Xenopsyllaconformisconformis2Genomovar172000-2001长爪沙鼠M.unguiculatus21Genomovar17长爪沙鼠M.unguiculatus4Genomovar11荒漠毛足鼠Phodopusroborovskii1Genomovar17黑线仓鼠Cricetulusbarabensis1Genomovar17五趾跳鼠Allactagasibirica1Genomovar17秃病蚤蒙冀亚种N.laevicepskuzenkovi2Genomovar172006长爪沙鼠M.unguiculatus7Genomovar20同形客蚤X.conformisconformis2Genomovar20秃病蚤蒙冀亚种N.laevicepskuzenkovi1Genomovar20菌株合计total48
4 讨 论
陕西省疫源地仅分布于定边县。该县隶属于榆林市,地处陕西省西北角、榆林市的最西端,是黄土高原与内蒙古鄂尔多斯荒漠草原过渡地带,位于东经107°15′至108°22′,北纬36°49′至37°53 ′。东至东南与本省靖边县、吴起县相连;南至西南与甘肃省华池县、环县相接;西与宁夏回族自治区盐池县毗邻,北至东北与内蒙古鄂托克前旗、乌审旗相邻,系陕、甘、宁、蒙四省区交界地。1987-1988年份、2000-2001年份鼠间疫情是在获悉内蒙宁夏发生鼠疫后,扩大检索,发现疫情的。疫情是否受到两地的波及,从DFR基因组型分型结果来看,陕西发现的3个基因组型,为内蒙古高原长爪沙鼠鼠疫疫源地的主要基因组型[8],且Genomovar20型仅分布于长爪沙鼠鼠疫疫源地,从分子分型角度证实了陕西鼠疫疫源地为鄂尔多斯高原长爪沙鼠鼠疫疫源地的一部分,据此推测陕西疫情与两地疫情关联,因内蒙古、宁夏接壤区域对应流行年代菌株尚无文献可查,这一点还有待于以后验证。
陕西自1987年被判定为鼠疫疫源地以来,共发生了3起鼠间疫情,从DFR基因组分型看,1987-1988年份、2000-2001年份前流行主要基因组型一致,三起疫情流行的主导组型不一致,2006年份流行的主导组型在2000-2001年份尚未检测到,说明鼠间疫情不受前两次的影响,有自身的规律。从表3可以看出,同一块鼠疫疫源地主要基因组型与菌株来源宿主、媒介无关联。
查阅资料1987-1988年资料,参考马国柱[9]、岳永杰[10]对2000-2001、2006年菌株分析,三起鼠间疫情鼠疫耶尔森菌脱氮阴性,甘油+,鼠李糖-,麦芽糖-,阿胶糖+(1987年有2株不发酵),根据纪树立分型标准属于鄂尔多斯高原生态型[2]。尤其是2000-2001、2006年份,菌株生化特性完全一致。但本研究发现这两次疫情发生期间,菌株DFR基因组型却不一致。说明传统的生态分型不够细致,而分子分型不仅可以把不同的菌株分开,还能把相似的菌株分开。
鼠疫耶尔森菌的基因组型均局限于特定的地理区域, 通常是完整的鼠疫自然疫源地或是部分地域, 每个地理区域都有其独特的环境、宿主、鼠疫耶尔森菌组合[11]。决定鼠疫耶尔森菌存在及其型别的因素可看作是一个由自然环境、贮存宿主、传播媒介、病原体相互作用的复合体(宿主生态位),在不同的地理区域中,起主要作用的因素各有不同。本研究发现陕西鼠疫菌DFR基因组型为Genomovar 11、17、20,1987-1988年份、2000-2001年份DFR基因组型以17为主,2006年份以20为主,表明引起陕西省鼠疫疫源地鼠间鼠疫流行的菌株已经发生了变化。基因型G20与G11相比,缺失了DFR12和18 位点,与G17相比,缺失了DFR12位点,说明鼠疫菌种内正经历着典型的基因组平行微进化过程,基因缺失不断发生,推动了鼠疫菌基因组的简约进化[12]。这与其地理生态环境密切相关,一定程度上反应了鼠疫菌适应自然选择的结果。
陕西鼠疫耶尔森菌DFR基因组型有Genomovar11、17、20共3种。17型多分布在定边的周台子乡、定边镇疫点及盐场堡狼塘疫点,20型分布在红柳沟镇黄沙窝疫点及盐场堡的马圈村。这3个基因组型为内蒙古高原长爪沙鼠鼠疫自然疫源地主要基因组型,文献[8]将3044株鼠疫菌分为52个基因组型,其中Genomovar20有73株,全分布于该疫源地。据了解,所有疫点生境植被差别不是太大,从图1可以看出,4个疫源乡镇连成一片,中间没有特殊屏障隔开,分析不同疫点基因组型不同是鼠疫菌适应疫源地大环境改变的结果。这一说法有待以后验证。
[1] Cong XB,Yin WW. Emergency Handbook of plague control and prevention[M] .Beijing:Peking University Medical Press.2009:13.(in Chinese)
丛显斌, 殷文武.北京:鼠疫防控应急手册[M].北京大学医学出版社,2009:13.
[2] Ji SL,Zhang HJ,Liu YP,et al.The typing and ecology,epidemioiogical significance of Chinese Yersinia pestis [J].Chin J Endemiol,1987,6(5):257-263.(in Chinese)
纪树立,张海峻,刘云鹏,等.中国鼠疫菌分型及其生态学流行病学意义[J].中国地方病学杂志,1987,6(5):257-263.
[3] Zhou DS,Yang RF. Strategies on the evolutionary for Bacterial Genomes[J].J Micro, 2004,24(1):34-41. (in Chinese)
周冬生,杨瑞馥.细菌基因组进化的分子策略[J].微生物学杂志,2004,24(1):34-41.
[4] Li YJ.Study of genomic polymorphism inYersiniapestisand construction of the rapid source—tracking pipeline[D].Beijing: Academy of Military Medical Sciences,2009.(in Chinese)
李艳君.鼠疫耶尔森氏菌基因组多态性研究及快速鉴定溯源系统的建立[D].北京:中国人民解放军军事医学科学院,2009.
[5] Zhou DS,Han YP,Song YJ,et a1.DNA microarray analysis of genome dynamics inYersiniapestis:insight into bacterial genome microevolution and niche adaptation[J].Med J Chin PLA,2004, 29(3):204-210.(in Chinese)
周冬生, 韩延平, 宋亚军,等. 鼠疫耶尔森氏菌基因组进化与生态位适应研究[J].解放军医学杂志,2004,29(3):204-210.
[6] Li Y, Dai E, Cui Y, et al. Different region analysis for genotypingYersiniapestisisolates from China[J]. PLoS One, 2008, 3(5): e2166. DOI:10.1371/journal.pone.0002166
[7] Achtman M, Zurth K, Morelli G, et al.Yersiniapestis, the cause of plague, is a recently emerged clone ofYersiniapseudotuberculosis[J]. Proc Natl Acad Sci U S A, 1999, 96: 14043-14048.
[8] Yang XY,Wei BQ,Jin J, et al. Regional genotyping and the geographical distribution regardingYersiniapestisisolates in China[J]. Chin J Epidemiol, 2014,35(8),943-948. (in Chinese)
杨晓艳,魏柏青,靳娟,等. 中国鼠疫耶尔森菌差异区段分型及其地理分布特征[J].中华流行病学杂志,2014,35(8),943-948.
[9] Ma GZ,Zhang FL,Wang L,et al, Identification and analysis of the biological characteristics ofY.pestisstrains isolated from Shaanxi Province 2000 to 2001 [J]. China Endem, 2003,18(3):163-164. (in Chinese)
马国柱,张福利,王丽,等.陕西省2000-2001年分离鼠疫菌株特性及分析[J].中国地方病防治杂志,2003,18(3):163-164.
[10] Yue YJ,An CH,Sun YX.Identification and analysis of the biological characteristics ofYersiniapestisstrains isolated in Shaanxi in 2006[J]. China Endem, 2003,18(3):163-164. (in Chinese)
岳永杰,安翠红,孙养信.陕西省2006年鼠疫菌生物学特性及分析[J].中国地方病防治杂志,2009,24(3):204-206.
[11] Rang RF,Huang PT.Comparative and evolutionary genomics of Yersinia pestis[J].Med J Chin PLA,2004,29(3):189-191.(in Chinese)
杨瑞馥,黄培堂. 鼠疫耶尔森菌比较和进化基因组学研究[J] .解放军医学杂志,2004,29(3):189-191.
RegionalgenotypingandepidemiologicalcharacteristicsregardingYersiniapestisisolatesinShaanxiProvince,China
AN Cui-hong, CHEN Bao-bao, LYU Wen, NIE Shou-min, FAN Suo-ping, SUN Yang-xin
(ShaanxiCenterforDiseaseControlandPrevention,Xi’an710054,China)
We typedYersiniapestisisolated from plague foci of Shaanxi Province using different region (DFR) and analyzed epidemiological characteristics. Twenty-three DFRs primers and PMT1 (plasmid) primer were used to verify the DFR genomovars and 48Yersiniapestiswere involved to analyze DFR profiles and epidemiological characteristics. In the same year, the genotypes ofYersiniapestisisolated from different infected vector and animals were basically the same. Three genomovars named Genomovar 11, 17, and 20 were verified in 48Yersiniapestisstrains in Shaanxi Province. The main genotypes were different in different epidemic years. In 1987-1988 and 2000-2001 years, genomovar 17 was major genomovar and genomovar 20 in 2006 year. In conclusion, the dominant genotypes were different in different epidemic years. As time goes on, DFR genomovars ofYersiniapestisundergone the evolution of gene deletion, which changes genomovar 17 into genomovar 20.
Shaanxi Province;Yersiniapestis; DFR genotyping; epidemiological characteristics
Sun Yang-xin,Email: sxpco@126.com
10.3969/j.issn.1002-2694.2017.10.013
孙养信, Email: sxpco@126.com
陕西省疾病预防控制中心,西安 710054
378
A
1002-2694(2017)10-0916-04
2017-05-10编辑李友松
猜你喜欢
中国人兽共患病学报(2023年8期)2023-09-18 02:53:38
中国药学药品知识仓库(2021年12期)2021-12-07 00:43:43
意林·少年版(2021年13期)2021-08-30 16:26:41
中国人兽共患病学报(2019年6期)2019-07-16 08:24:40
饲料博览(2019年1期)2019-02-14 21:44:22
中国学术期刊文摘(2019年23期)2019-01-29 11:49:47
浙江医学(2018年16期)2018-09-08 05:58:00
兽医导刊(2016年12期)2016-05-17 03:51:53
中国人兽共患病学报(2015年12期)2015-01-25 03:21:47
实验动物与比较医学(2014年5期)2014-02-28 14:53:08
