
笼养川金丝猴个体间行为时间分配及空间利用差异
2017-11-17 10:06张明睿滕丽微刘振生王爱善
野生动物学报 2017年3期
张明睿 滕丽微,2 刘振生,2* 王爱善
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.国家林业局野生动物保护学重点开放实验室,哈尔滨,150040;3.上海动物园,上海,200335)
笼养川金丝猴个体间行为时间分配及空间利用差异
张明睿1滕丽微1,2刘振生1,2*王爱善3
(1.东北林业大学野生动物资源学院,哈尔滨,150040;2.国家林业局野生动物保护学重点开放实验室,哈尔滨,150040;3.上海动物园,上海,200335)
2012年11月~2013年5月,采用瞬时扫描取样法,对上海动物园内4只笼养川金丝猴(Rhinopithecusroxellana)的活动时间和笼舍空间利用进行比较研究。不同于其他行为学研究,本文利用环境优势将个体区分研究,统计个体对不同类型空间的利用情况来补充并解释川金丝猴的行为选择。结果表明,笼养川金丝猴休息行为最多,占56.80%。个体间在运动、理毛、摄食、自我梳理行为上差异极显著(P<0.01),亚成体运动时间与雌2自我梳理时间明显高于其他个体。社会单元中雄性个体与雌1个体进行的关联行为最多,其次为亚成体与雌2个体。雄性个体与雌1个体的行为具有排他性。空间利用率上,假山区利用最多(48.20%),主要用来休息。其次为丰容区(32.81%),是个体运动与采食的主要区域。前笼区占12.00%,与进行较多的乞食有关。个体间对不同区域的利用差异显著(P<0.05)。雄性个体和雌1个体对假山区域利用较多,雌2个体和亚成体对丰容区利用较多,亚成体对前笼区利用明显高于其他个体,雌2个体对草地区利用明显高于其他个体。
川金丝猴;时间分配;空间利用率;瞬时扫描取样法;上海动物园
川金丝猴(Rhinopithecusroxellana)又名仰鼻猴,隶属灵长目(Primates)猴科(Cercopithecidae)疣猴亚科(Colobinae)金丝猴属[1],是我国特产种类,被列为国家I级重点保护野生动物,世界自然保护联盟(IUCN)所列濒危(EU)种[2]、濒危野生动植物国际贸易公约(CITES)附录Ⅰ物种。川金丝猴是典型的森林树栖动物,栖息海拔为1 500~3 300 m。在我国的分布主要限于北亚热带,西起横断山北部,东连秦岭南北坡,秦岭为其分布的最北限[3],神农架为其分布的最东限[4]。
将川金丝猴圈养属于迁地保护(off site conservation)的一种形式。迁地保护是相对于就地保护而言的对珍稀动物的保护方法[5]。将濒危动物迁移到人工环境中可以为其提供优越的生存条件从而保存物种的基因库并增加其数量[6-7]。据统计,圈养川金丝猴数量在2002年底为185只,但7 a之后存活数量上涨到227只[8]。
相对于野外观察,对动物园的圈养动物进行行为的观察与记录更加容易且数据详细准确。同时动物园对圈养个体的挑选和笼舍丰容的工作都具有代表性,相当于对野外环境的简化,可作为研究物种的较理想的模型。然而人工提供的圈养条件相对于野外栖息地缺乏复杂性[9],灵长动物行为时间的分配受到环境条件[10]、食物资源的分布与数量[11-12]、种内及种间关系[13-15]和人为活动[16]等的影响。在圈养条件下,这些因子均与自然环境中不同。研究圈养下的川金丝猴行为可以与野生状态下的川金丝猴行为规律形成对照,发现其行为的异常[17],从而更好地改善笼舍环境设计,使其更接近自然状态,促进个体正常行为的表达和繁殖成功[18]。
对灵长类动物行为学研究通常由于野外个体较多、难以准确辨别而将群体的性别组与年龄组作为研究对象,本次研究利用动物园有利条件将每个个体进行区分,研究个体间的差异。另一方面,在野外,以社群形式存在的非人灵长类种群的空间结构是物种在进化过程中对其生存环境适应的结果[19]。对物种具体空间的利用情况的研究可以更好地解释其行为的成因,也可以与行为相结合,提供物种某一阶段在生存环境中的具体特点。本文对笼养川金丝猴的空间利用情况进行研究,找出每个个体对空间利用的差异,并与行为相结合来更好地描述笼养川金丝猴在一段时间内的行为学特点。
1 研究对象与笼舍环境
1.1 研究对象
2012年11月~2013年5月,选取上海动物园的1个笼舍内的川金丝猴群体作为研究对象。包括1只成年雄性,2只成年雌性和1只亚成体雌性(表1)。亚成体雌性为此雄性与雌性之一所生。
表1 上海动物园圈养川金丝猴谱系

Tab.1 The captive Rhinopithecus roxellana group in Shanghai Zoo
1.2 笼舍环境
笼舍环境为笼网式笼舍,面积约为190 m2,为了便于记录与研究,将外笼舍简化成5个区域(图1)。

图1 川金丝猴笼区空间划分Fig.1 Space utilization distribution of the cage
假山区(A:rockery area)有低矮灌木及为川金丝猴提供休息和乘凉避雨空间的松木小房;高空区(B:ceiling area)为笼舍的拱形顶部,有供川金丝猴攀爬的笼网和吊绳;丰容区(C:enrichment area)占笼舍的面积较大,分布着假树、栖架、挂网、铁网球、吊绳及轮胎等设施;前笼区(D:visit area)与游客游览通道相邻;草地区(E:grass area)位于丰容区与外笼区之间。
2 研究方法
2.1 数据的采集
在研究期间于外笼区不同角度安设4台摄像头(可以覆盖绝大部分川金丝猴活动区域)记录笼内川金丝猴个体行为。每个月记录10~17 d不等。
本研究需要对川金丝猴群体中的每一只个体进行取样,故采用行为学研究中的扫描法(Scan samping)进行取样[20]。在笼养条件下影响川金丝猴个体行为的随机外界因素(如食物分布的不确定性、天气的突变、其他种群及同物种其他社会单元的干扰等)极少,其行为单一且持续时间较长。故采用行为学研究中的瞬时记录法(Instantaneous sampling)。该记录法所得到的数据偏差极小,准确可靠,并可以节省时间来充分多次利用设备而延长整体数据采集的持续时间。瞬时扫描法[21]是研究川金丝猴行为的常用方法,在视觉良好的情况下效果极佳[22],本研究条件适于该方法。
根据川金丝猴实际活动情况选取每隔10 min记录川金丝猴的行为及其所在区域,区分记录每个个体。由于川金丝猴是典型的昼行性动物,且夜间主要在较狭小隐蔽的内笼区休息,行为与空间利用情况单一,故记录时间为每天昼间7:00~17:00。
参考苗秀莲对笼养川金丝猴建立的行为谱并结合实际观察的情况制定本研究的行为谱[23]。
休息:个体没有明显的位置移动变化,身体为坐、卧或者趴的状态,包括头部转动等微小的动作。
运动:个体从甲地行走或攀爬到乙地,全身性活动行为。
摄食:指用手将食物送入嘴内并伴有咀嚼动作。
采食:指个体在笼舍内主动寻找并接近管理员投喂以及笼内生长的食物,将食物就地或者带到其他地方的过程。与摄食行为不同,采食行为体现了川金丝猴个体自主寻找食物的过程,与野外的采食行为相似,并与不正常的乞食行为形成对比。
乞食:攀爬在笼壁上或者坐于笼舍边缘,上肢伸出笼外,手掌摊开或者半握,向游人索要食物。
拥坐:个体拥坐时面对面腹部相对并紧挨,一方或双方伸出双臂抱住对方,一方的头部放在另一方的胸前或肩上。
理毛:指某一个体给另一个体清理皮毛的行为。
自我梳理:自我理毛者采取坐姿,自我梳理的部位一般为大腿、小腿及尾部等手容易接触到的地方。可看成个体以自己作为对象进行理毛行为的过程。
搔痒:个体用上肢快速抓挠自己身上的某个部位,与自我梳理的区别是个体眼睛并不注视梳理部位,也不认真、慢速地整理。
交配:由于发生频率较少,可以视作个体所有的繁殖过程,主要部分为爬跨行为。
由于丰容工作较好,观察过程中未发现刻板行为。由于空间及个体数的限制,其他复杂行为发生极少,即将其归入运动与休息两大行为中。
2.2 数据的处理与分析
将每次对每个个体的行为扫描视为一个独立样本,以每个个体每小时活动时间分配作基本单元,求出需要的每天行为时间分配、每月行为时间分配等数据。用同样的方式得出需要的空间利用数据。
数据的处理与分析利用Microsoft Excel 2007与SPSS 22软件完成。
3 结果与分析
3.1 笼养川金丝猴行为时间分配
3.1.1 川金丝猴总行为时间分配
将所有个体行为时间分配合并,得到川金丝猴总行为时间分配,各行为所占比如图2所示。

图2 笼养川金丝猴行为总时间分配Fig.2 The time budgets for the behaviours of the whole captive Rhinopithecus roxellana individuals
3.1.2 川金丝猴个体行为时间分配差异
如表2所示,用非参数性检验(Kruakal-WallisH test,n=280)对金丝猴不同个体间行为进行检验。个体间在采食行为上差异性显著(P<0.05),雄与亚的采食行为明显较多。个体间在运动、摄食、理毛、自我梳理行为上有极其显著的差异(P<0.01)。
表2 笼养川金丝猴不同个体间昼间活动时间分配(%)
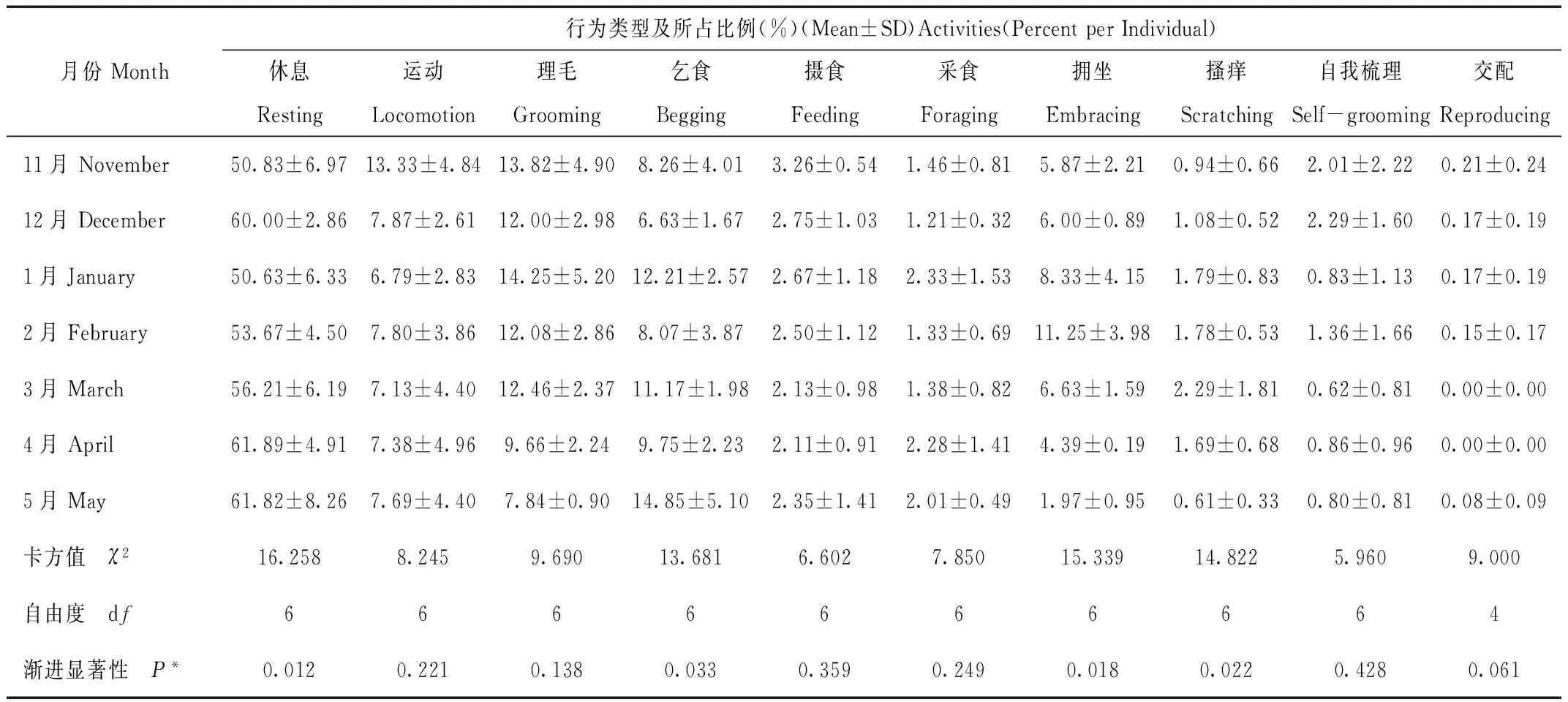
Tab.2 The diurnal activity budgets of each captive Rhinopithecus roxellana individual
3.2 关联行为统计
在群居性动物中,个体间通过行为上的相互作用来显现个体间与社群内的联属关系[24]。为了通过行为来更深探讨这种联属关系,在1次扫描记录中将2只或以上个体在距离小于1 m的情况下进行同一行为的情况记录下来,称为关联行为。关联行为分为社会行为与普通行为。社会行为又分为理毛和拥坐,都反映了个体间的亲密程度。而理毛与拥坐不同,反映个体间社会地位的差别。剩下的为普通行为(行为谱中除理毛与拥坐2种行为外其他所有行为,由于重要性没有此2种行为高,即在本研究中合并在一起进行统计),发生频率较大,但体现联属关系的程度不如社会行为。
在总行为时间中,关联行为占53.20%。其中理毛占25.04%,拥坐占11.68%,普通行为占63.28%。
3.2.1 个体理毛行为时间统计
本研究将理毛行为定为个体间互相的梳理,理毛存在行为的发出者与接受者,所以将不同的理毛行为区分记录(表3),当理毛行为的发出者和接受者是同一个体时,表中显示的时间为自我梳理行为的时间[25]。
表3 笼养川金丝猴理毛行为时间统计

Tab.3 The time for different kinds of grooming behavior of captive Rhinopithecus roxellana
注:A表示行为发出者,R表示行为接受者
Note:A represents the individual which gave the grooming behavior,R represents the individual which received the grooming behavior
需要说明的是,由于本次研究采用的瞬时扫描法是每10 min进行1次扫描,表3的10 min是扫描的一个时间单位,也可以看作是每次扫描中不同形式的理毛行为所出现的频次。
3.2.2 拥坐行为中个体组合比例
拥坐行为中,雄与雌1拥坐占比例最大,为61.5%,其次为亚与雌2占38.27%。亚、雌1与雄拥坐及亚与雄拥坐各占1.00%与0.17%。可以看出在拥坐行为中雄与雌,亚与雌2关系亲密且与此2种组合外的个体拥坐概率极小。亚与其父母拥坐行为仅为1.00%,说明其和雌2更亲密,也说明雄与雌1在拥坐行为上具有排他性。
3.2.3 普通行为中个体组合比例
普通行为中,比率依次为:雄雌160.56%、亚雌222.22%、亚雄雌111.92%、亚雌12.75%、亚雄1.42%、雌1雌20.90%、其他0.23%。可看出,雄雌1之间关系依然是最亲密的,其次是亚雌2。说明亚有机会与其父母互动,而雌2则是有意与雄和雌1避开活动。
3.3 笼养川金丝猴空间利用
3.3.1 调查期间川金丝猴总空间利用情况
川金丝猴活动最多的区域是A区,占48.20%,因为假山区上用于运动的设施较少且隐蔽地点较多,川金丝猴在A休息时间较长。其次是C区,占32.81%,因为丰容区是川金丝猴昼间运动与采食的主要区域。D区占12.00%,与个体在外笼区进行较多的乞食活动有关。在B高空区和E草地区活动分别占5.35%与1.64%。
3.3.2 川金丝猴个体空间利用差异
对川金丝猴不同个体对空间利用的情况进行非参数性检验(Kruakal-WallisH test,n=140)。结果(表4)显示个体间对不同空间的利用情况存在着明显的不同。雄与雌1对A假山区利用较多,而亚与雌2对C丰容区和B高空区利用较多。这说明雄与雌1更多的是在假山区休息,而亚与雌2则更偏向于在丰容区运动和休息,由于在B区不易停留,可以看出亚与雌2更倾向于运动。亚利用D区域明显高于其他个体,说明其乞食行为多于其他个体。雌2在E草地区停留时间较长,说明与其他个体较疏远。

表4 笼养川金丝猴不同个体间昼间活动区域分配(%)
4 讨论
4.1 笼养川金丝猴行为时间分配特点与个体间差异
研究表明笼养川金丝猴的大部分行为时间分配在休息上。一方面,这符合灵长类行为分配中休息占比很大的普遍规律,对此现象的解释一种为,在摄食大量植物后,灵长类需要较长时间进行发酵、分解以及吸收食物中的营养[26-27],另一种解释是因为觅食树叶所获得的能量有限,为了减少能量消耗,灵长类动物必须减少过多的活动[28-29];另一方面,笼养情况下休息时间分配明显多于野生状态下(26.83%)[30],其原因是笼养条件下食物充足,川金丝猴分配在采食上的时间极少。加上笼舍提供的环境不是典型的川金丝猴所适应的高山森林环境[31],所以休息行为占川金丝猴行为时间较多。除休息之外,理毛与乞食占比较大。理毛作为在野生川金丝猴种群中最常见的社会行为[32-33],在本研究中与在野外研究中所占时间几乎相同(11.56%与11.58%),说明在笼养环境下川金丝猴个体之间存在基本的社会关系。在野外并不会发生的乞食行为在本研究中占较大的比例(10.09%),而正常的摄食与采食行为占比极低(4.27%)。一方面是由于个体在动物园环境下不受食物条件约束,获得食物难度极低;另一方面是因为动物园所提供的食物营养较丰富,减少了摄食量。乞食行为较多说明笼养川金丝猴对人类的依赖较高,这对迁地保护是不利的。
作为研究对象的4只川金丝猴个体,在各自行为分配上存在着明显的差异。亚成体的运动行为明显高于其他成年个体,学者对此的解释为较年轻的个体会通过进行更多的社会性玩耍学习和掌握其物种所特有的全部行为技能,能够更好地生存下来,是一种适应[34]。而雌2个体进行的运动时间比另外2只多,这可能是因为它与亚成体比较亲密,且有意避免与雄和雌1接触。在理毛行为上,雄与雌1进行的最多且占比接近,说明它们关系亲密。而雌2的自我梳理行为最多,这是作为缺少理毛对象而进行的一种补偿。
从行为时间的分配情况我们可以看出,动物园的饲养工作虽然在饲料搭配与时间安排上已经非常合理,但是饲喂形式比较单一,因为在野外条件下,食物因素往往影响到整个川金丝猴种群的个体及社会行为时间分配,所以为了调动笼养川金丝猴个体的积极性,使其行为时间分配更接近于野外条件,应针对不同个体进行区分饲喂,并将食物放置在不同区域内供个体寻找。这样可以有效降低休息及乞食行为所占比例,增加采食行为占比。
4.2 笼养川金丝猴个体关系讨论
绝大多数野生川金丝猴的社会单元可以分为一雄多雌以及子女单元(OMU)和全雄单元(AMU)[35]。作为本研究对象的金丝猴群体可以看成是最简单的OMU。
在理毛行为上,雄性接受的理毛行为最多,这符合在野外社会等级最高的个体会得到最多的理毛的规律。然而进一步的整理发现,雄与雌1,亚成体与雌2互相之间的理毛占个体间此行为的绝大多数,亚成体会偶尔为其父母或接受其父母(雄与雌1)的理毛,而雌2个体不存在与雄和雌2理毛的情况。而在对拥坐与其他行为的研究上更证明了这一点,即雄与雌1总是一同行动,亚成体与雌2总是一同行动,亚成体并不会排斥与其父母的互动,而雌2个体会有意地避免与雄1和雌2的互动。
在野外环境中,虽然一个OMU的社会单元中的成年雌性存在着等级差别,但基本上雄性都会与其交配并保持较亲密的关系,未在生育期的雌性个体会给雄性更多的理毛来交换更多交配的可能,而在生育期的雌性个体会得到更多其他雌性个体的理毛,幼体会得到其母亲更多的理毛机会[33]。然而在此笼养环境中,情况却不同。作者对其解释为,雄性个体过于对雌1个体进行关注,而对雌2个体进行更多的攻击行为。由于雄性的过分关注,使得雌1个体与其孩子亚成体的互动减少,而由于雌2个体的存在使得亚成体更倾向于选择雌2个体共同活动。而在此单元中,雌2个体得到的来自其他个体的关注最少,这一点可以从它较多的自我梳理行为上看出来,此行为可以视为对理毛行为的一种补偿。
从研究结果可以看出来,如果想要达到个体间正常的野外社群关系,改善雌2与雄、雌1之间的关系是解决的办法。
4.3 笼养川金丝猴空间利用情况
在对笼养川金丝猴个体空间利用的统计中,A假山区占比最大,这与假山区为个体提供的隐蔽物较多更适合个体休息有关,与休息行为在所有行为中占比最大相符合。而C丰容区提供的丰容设施则为个体运动、采食等行为提供很棒的条件,个体也会在C丰容区休息,故C丰容区占比也很高。对B高空区和E草地区的利用极少则与野生川金丝猴生活在丛林环境的习性有关。D外笼区则是川金丝猴与游客进行互动及乞食的主要地点,应尽量减少其在此活动的概率。
不同个体对空间利用情况的差异研究,作者发现雄性个体和雌1个体,与亚成体和雌2个体喜欢利用的区域正好相错,雌2个体对E区的利用明显高于其他个体。这也从空间角度上证明在个体关系讨论中得出的结论,即雄性过于关注雌1个体并排斥雌2个体从而导致此社群单元个体之间的关系与野生环境下不同。
4.4 管理建议
首先,笼养川金丝猴较野外川金丝猴休息时间长,自主采食时间较少,不正常的乞食行为占比很高。对于此问题,一方面要定期改变丰容设施来提高个体对设施的兴趣[36-37],在A区也应该设置一些供个体攀爬的设施来尽量减少个体的休息时间;另一方面应该在1 d内多次投喂食物,将食物放置在个体稍有难度接触到的地方,以激发个体对采食的欲望并根据不同个体的营养需要来区分饲喂,这样在乞食行为中贡献最多的幼体的此行为会明显减少。
第二,过多的乞食行为会对川金丝猴造成不良影响,对此,除了上述对饲养方式的建议外,动物园还应该加强管理,对投喂川金丝猴的游客进行警告与教育。
第三,关于此川金丝猴社会单元的特殊情况,制定出增加雌2与其他个体互动机会的计划是解决问题的关键,而首先要做的就是找出雄与其疏远的原因并进行改善措施。如果有必要,可以考虑引进新的川金丝猴个体或交换已有个体来改善个体之间的关系。
[1] 杨士剑,王政昆,张峻.疣猴亚科及仰鼻猴的分类研究进展[J].云南师范大学学报:自然科学版,2003,23(5):51-54.
[2] Long Y,Richardson.Rhinopithecusroxellana[DB/OL].The IUCN Red List of Threatened Species,2008.Http://www.iucnredlist.org/details/19596/0.
[3] 解文治,陈服官.川金丝猴(Rhinopithecusroxellana)的行为观察和社会结构的空间配置[M].西安:西北大学出版社,1989:243-250.
[4] 张荣祖.中国动物地理[M].北京:科学出版社,1999.
[5] 黄雪蔓,周岐海,江峡,等.笼养松鼠猴的活动时间分配[J].广西师范大学学报:自然科学版,2010,28(2):103-106.
[6] 李俊清,李景文,崔国发.保护生物学[M].北京:科学出版社,2012:188-194.
[7] 陆建身.中国生物资源[M].上海:上海科技教育出版社,1997:398-401.
[8] 于泽英.川金丝猴圈养种群繁殖健康度的比较分析[J].野生动物,2011,32(2):69-72.
[9] Britt A.Environmental enrichment for apes:a literature review[C]//Field D A.Guidelines for environmental enrichment.London:Association of British Wild Animal Keepers,1998:233-247.
[10] Hanya G.Seasonal variations in the activity budget of Japanese macaques in the coniferous forest of Yakushima:effects of food and temperature[J].American Journal of Primatology,2004,63(3):165-177.
[11] Umapathy G,Kumar A.Impacts of the habitat fragmentation on time budget and feeding ecology of lion-tailed macaque(Macacasilenus)in rain forest fragments of Anamalai Hills,South India[J].Primate Report,2000,58:67-82.
[12] Pavelka M S M,Knopff K H.Diet and activity in black howler monkeys(Alouattapigra)in southern Belize:does degree of frugivory influence activity level?[J].Primates,2004,45(2):105-111.
[13] Teichroeb J A,Saj T L,Paterson J D,et al.Effect of group size on activity budgets of colobus vellerosus in Ghana[J].International Journal of Primatology,2003,24(4):743-758.
[14] Hemingway C A.Time budgets and foraging in a Malagasy primate:do sex differences reflect reproductive condition and female dominance?[J].Behavioral Ecology and Sociobiology,1999,45(3/4):311-322.
[15] Tsuji Y,Takatsuki S.Food habits and home range use of Japanese macaques on an island inhabited by deer[J].Ecological Research,2004,19(4):381-388.
[16] 李义明,廖明尧,喻杰,等.社群大小的年变化、气候和人类活动对神农架自然保护区川金丝猴日移动距离的影响[J].生物多样性,2005,13(5):432-438.
[17] 余建秋,刘选珍,王强,等.环境丰容对圈养川金丝猴行为影响的初步研究[J].四川动物,2015,34(2):245-250.
[18] Traylor-Holzer K,Fritz P.Utilization of space by adult and juvienile groups of captive chimpanzees(Pantroglodytes)[J].Zoo Biology,1985,4(2):115-127.
[19] 郭程,向左甫,任保平,等.滇金丝猴群中全雄单元在移动时的空间结构与功能[J].中南林业科技大学学报,2011,31(7):136-139.
[20] Martin P,Bateson P.Measuring behaviour:an introduetory guide [M].2nd ed.Cambridge:Cambridge University Press,1994:84-88.
[21] Altmann J.Observational study of behavior:sampling methods[J].Behaviour,1974,49(3):227-266.
[22] Harcourt A H,Stewart K J.Gorillas’ time feeding:aspects of methodology,body size,competition and diet[J].African Journal of Ecology,1984,22(3):207-215.
[23] 苗秀莲.笼养川金丝猴的活动规律和时间分配及社会行为的研究[D].济南:山东师范大学,2007.
[24] 李娟娟.秦岭川金丝猴(Rhinopithecusroxellana)OMU间联署关系的研究[D].西安:西北大学,2014.
[25] 张德军,黎大勇,胡杰,等.滇金丝猴雌性个体间的理毛行为[J].兽类学报,2014,34(1):38-45.
[26] Clutton-Brock T H,Harvey P H.Species differences in feeding and ranging behaviour in primates [C]//Clutton-Brock T H.Primate ecology:studies of foraging and ranging behaviour in lemurs monkeys and apes.London:London Academic Press,1977:557-584.
[27] Stanford C B.The capped langur in Bangladesh:behavioral ecology and reproductive tactics[M].Karger Medical and Scientific Publishers,1991.
[28] 黄乘明.中国白头叶猴[M].桂林:广西师范大学出版社,2002:1-17,49-62.
[29] 熊金荣.笼养灰叶猴(Trachypithecusphayrei)的夏季日活动时间分配[J].生物学杂志,2007,24(6):46-48.
[30] 吕九全,李保国.秦岭川金丝猴的昼间活动时间分配[J].兽类学报,2006,26(1):26-32.
[31] Jablonski N G.The natural history of the doucs and snub-nosed monkeys[M].Singapore:World Scientific,1998:1-9.
[32] 禹洋.川金丝猴(Rhinopithecusroxellana)相互理毛行为研究[D].长沙:中南林业科技大学,2012.
[33] 夏东坡.短尾猴理毛行为交换策略与群体稳定性研究[D].合肥:安徽大学,2013.
[34] Poirier F E,Bellisari A,Haines L.Functions of primate play behavior [C]//Smith E O.Social play in primates.New York:Academic Press,1978:143-168.
[35] 任仁眉,严康慧,苏彦捷,等.金丝猴的社会野外研究[M].北京:北京大学出版社,2000:1-222.
[36] 陈炜,申琦,马清义,等.圈养秦岭羚牛夏季昼间活动时间分配研究[J].经济动物学报,2006,10(4):211-214.
[37] 滕丽微,刘旭,刘振生.圈养条件下小熊猫指名亚种雨季的活动规律[J].经济动物学报,2013,17(2):67-70.
Differences of Time Budgets and Space Utilization in CaptiveRhinopithecusroxellana
Zhang Mingrui1Teng Liwei1,2Liu Zhensheng1,2*Wang Aishan3
(1.College of Wildlife Resources,Northeast Forestry University,Harbin,150040, China;2.Key Laboratory of Conservation Biology,State Forestry Administration, Harbin,150040,China;3.Shanghai Zoo,Shanghai,200335,China)
From November 2012 to May 2013,we studied the time budgets and space utilization of 4 Sichuan golden snub-nosed monkeys(Rhinopithecusroxellana)by instantaneous and scan sampling methods in an enclosure at Shanghai Zoo.The differences between individuals were also compared and analyzed.In contrast to previous behavioral studies,we differentiated individuals and introduced the space factor to refine and explain how behaviors affect captive monkeys.Resting was the dominant activity,accounting for 57%.Individual monkeys varied in the amount of time expended in locomotion,grooming,feeding and self-grooming(P﹤0.01).The time spent locomoting by juveniles was much higher than for other age classes.Self-grooming by female 2 also occupied more time than for others.In this tiny social group,male 1 and female 1 moved together most frequently,and the intimacy of juvenile and female 2 was second in this category.Activities of male 1 and female 1 were exclusive.Golden monkeys most often occupied the rockery area(48%)and mainly used this area for resting.This was followed by the enrichment area(33%).Individual diurnal movement and feeding activities were concentrated at the visit area(12%)where begging was recorded.Individual monkeys varied in their use of different areas(P﹤0.05).Male 1 and female 1 used the rockery area more than others,juvenile and female 2 used the enrichment area more than others,female 2 used the grass area far more than others,and juvenile used the visit area far more than others.
Rhinopithecusroxellana;Distribution;Utilization of space;Instantaneous and scan sampling;Shanghai Zoo
稿件运行过程
2017-03-15
修回日期:2017-05-08
发表日期:2017-08-10
Q958.1
A
2310-1490(2017)03-347-08
中央高校基本科研业务费资助项目(2572014CA03、DL13EA01);上海市绿化和市容管理局资助项目(F080401)
张明睿,男,26岁,硕士研究生;主要从事野生动物生态学研究。E-mail:zmr888887@163.com
*通讯作者:刘振生,E-mail:zhenshengliu@163.net
猜你喜欢
云南画报(2021年10期)2021-11-24
东方少年·布老虎画刊(2021年1期)2021-06-22
小哥白尼(野生动物)(2020年4期)2020-07-27
铁道通信信号(2020年9期)2020-02-06
劳动保护(2019年7期)2019-08-27
数学大王·趣味逻辑(2019年5期)2019-06-13
福建基础教育研究(2019年11期)2019-05-28
小学科学(学生版)(2019年5期)2019-05-21
学生天地(2018年36期)2019-01-28
中学科技(2015年1期)2015-04-28
