
无籽刺梨及其近缘种ITS序列分析
2017-11-15 02:31陈兴银石建明杨鹏张凯凯关萍
江苏农业科学 2017年17期
陈兴银+石建明+杨鹏+张凯凯+关萍


摘要:为探讨无籽刺梨及其近缘种的关系,对采自贵州兴仁的无籽刺梨(Rosa sterilis S. D. Shi)、贵州安顺的光枝无籽刺梨(Rosa sterilis S.D.Shi var.leioclada M.T.An,Y.Z.Cheng et M.Zhong)及采自贵州遵义、兴仁的缫丝花(Rosa roxburghii Tratt.)以及从美国国立生物技术信息中心(NCBI)网站上下载的长尖叶蔷薇(Rosa longicuspis Bertal.)进行内转录间隔区(ITS)序列分析。利用MEGA6.0软件对ITS序列进行分析,ITS序列总长为629 bp,其中ITS1序列长度为253 bp,5.8S长度为164 bp,ITS2长度为212 bp;对无籽刺梨、缫丝花、长尖叶蔷薇进行组合分析,组合1(无籽刺梨、长尖叶蔷薇、兴仁缫丝花)和组合2(无籽刺梨、长尖叶蔷薇、遵义缫丝花)中整个ITS序列共有26个变异位点,占序列总长度的4.10%,组合3(光枝无籽刺梨、长尖叶蔷薇、兴仁缫丝花)、组合4(光枝无籽刺梨、长尖叶蔷薇、遵义缫丝花)中共有30个变异位点,占序列总长度的4.76%。4个组合中ITS1变异位点数都为7个,占ITS1序列长度的2.76%,占序列总长度的1.11%;5.8S序列均没有变异位点;组合1、2中ITS2的变异位点数为19个,占ITS2序列长度的896%,占序列总长度的3.02%;组合3、4中ITS2的变异位点数为23个,占ITS2序列长度的10.8%,占ITS序列总长度的3.65%。4个组合中无籽刺梨与光枝无籽刺梨都含有2种碱基叠加的变异位点,叠加的2种碱基中1个来自长尖叶蔷薇,1个来自缫丝花,无籽刺梨同时具有缫丝花、长尖叶蔷薇的2种ITS序列。通过最大简约法(MP)进行聚类分析,结果显示,无籽刺梨与长尖叶蔷薇聚为1支,通过形态学比较发现,无籽刺梨与贵州缫丝花、缫丝花较为接近。研究结果为进一步探讨无籽刺梨的物种起源提供了一定的理论基础。
关键词:无籽刺梨;ITS序列;变异位点
中图分类号: S685.120.3文献标志码: A文章编号:1002-1302(2017)17-0042-05
通信作者:关萍,博士,教授,研究方向为植物分子生物学。E-mail:guanp508@163.com。无籽刺梨(Rosa sterilis S. D. Shi)别称搭钩刺梨,为蔷薇科(Rosaceae)蔷薇属(Roses)多年生落叶攀援性灌木,为贵州省特有种[1]。其果实清爽可口,酸甜适中,并富含维生素C、氨基酸、超氧化物歧化酶(SOD)、多种微量元素等,无籽刺梨因富含大量维生素C,得名“维C之王”,具有广阔的食品加工、营养保健品等资源开发前景[2-3]。无籽刺梨因不能产生可育种子而得名,无籽刺梨主要通过扦插进行无性繁殖,而多代的无性繁殖可能会导致无籽刺梨品质退化[4]。因此可见,分析无籽刺梨的内转录间隔区(ITS)序列,可為进一步选育无籽刺梨近缘种进行杂交,进而保存无籽刺梨优良特性奠定一定理论基础。
季祥彪等对无籽刺梨进行形态解剖研究,认为无籽刺梨有可能是1个自然杂交种[5];在时圣德等报道的蔷薇属新种中,无籽刺梨与缫丝花(Rosa roxburghii Tratt)近缘,其形态主要有小叶数量、花的颜色、果实大小及果实有无籽的差异[6]。邓亨宁等利用核基因和叶绿体基因片段进行建树,得出无籽刺梨为长尖叶蔷薇(Rosa longicuspis Bertal.)和缫丝花的天然杂交种[7]。杂交是指2个在遗传上有明显分化的个体间的交配,交配后一般不能产生有生殖能力的后代[8]。杂交,特别是异源杂交多倍化是新物种形成、多样化的重要机制之一[9]。传统上通常通过中间性状来判断一个种是否为杂交种,但是在一些中间性状不明显或者不清楚亲本的情况下,此方法会判断不准确。因此,正如Soltis等研究表明,即使已经从形态学或细胞遗传学上确定某个物种为杂交种,仍需要其他一些证据进一步验证[10-11]。相较于其他一些新方法,如同工酶、限制性片段长度多态性(RFLP)、PCR-单链构象多态性(PCR-SSCP)、简单重复序列(SSR)等,含有丰富的碱基变化及信息位点的核糖体DNA序列分析更能准确反映杂交种与亲本间的遗传关系[12-14]。本研究以天然杂交种无籽刺梨及其可能的亲本缫丝花、长尖叶蔷薇作为研究对象,以核糖体ITS基因作为分子标记,结合系统发育树分析,探讨无籽刺梨与缫丝花、长尖叶蔷薇在ITS序列上的变化规律和特点。
1材料与方法
1.1材料
内转录间隔区来自14个无籽刺梨的亲缘物种,其中6个种的ITS序列由测序所得,测序样品来自6个种的新鲜叶片,经硅胶干燥后,保存于-20 ℃,材料详细来源见表1。其他8个种的ITS序列从GenBank数据库中下载,长尖叶蔷薇是根据无籽刺梨ITS序列在美国国立生物技术信息中心(NCBI)网站上利用BLAST通过相似搜索法得到的最为相似的序列,详见表2。表1供试试验样品的采集地等信息
样品编号材料名称拉丁学名采集地1无籽刺梨Rosa sterilis S.D.Shi贵州省黔西南州兴仁县巴铃镇2光枝无籽刺梨Rosa sterilis S. D. Shi var. leioclada M.T.An,Y.Z.Cheng et M.Zhong贵州省安顺市3缫丝花Rosa roxburghii Tratt.贵州省黔西南州兴仁县巴铃镇4小果蔷薇Rosa cymosa贵州大学南区后山5缫丝花Rosa roxburghii Tratt.贵州省遵义市道真县上坝乡6金樱子Rosa laevigata贵州梵净山
1.2方法
1.2.1DNA提取称取经硅胶干燥的材料10~20 mg,用植物DNA提取试剂盒提取,试剂盒购自天根生化科技(北京)有限公司。DNA提取后,用1%琼脂糖凝胶与紫外-可见分光光度计检测DNA的质量和浓度,将DNA浓度稀释至 20 ng/μL,保存备用。
1.2.2PCR扩增与测序引物选用ITS1(5′-TCCGTAGG TGAACCTGCGG-3′)和ITS4(5′-TCCTCCGCTTATT GATATGC-3′)[15],引物序列由昆明硕擎有限公司合成。25 μL PCR反应体系:1 μL DNA模板,各1 μL正反向引物,125 μL Mix,用ddH2O补齐剩余体积。扩增程序:94 ℃ 2 min;94 ℃ 10 s,57 ℃ 45 s,72 ℃ 1 min,共计38个循环;72 ℃ 延伸 10 min。纯化后的PCR产物送至昆明硕擎有限公司进行双向测序。
1.2.3序列分析测序得到的序列利用DNAStar中的SeqmanⅡ软件进行编辑和拼接,并进行人工矫正。ITS序列的范围参照GenBank上缫丝花(AB038459.1)序列,确定样品ITS1、5.8S和ITS2的边界,拼接好的序列用MEGA6.0软件进行序列分析和系统发育分析,以自展法进行检测,共循环 1 000 次,采用最大简约法建树。
2结果与分析
2.1PCR产物的电泳检测
6个样品的DNA扩增产物经2%琼脂糖凝胶电泳分析,发现均有1条明显的750 bp左右的条带,与ITS序列大小相似,证明扩增程序可行,且扩增产物可用于测序。
2.2ITS序列分析
参照GenBank上缫丝花(AB038459.1)ITS序列的范围排序,数据库上的缫丝花序列排序后缺失3个碱基,2种来自不同地方的缫丝花及无籽刺梨、光枝无籽刺梨、下载的长尖叶蔷薇(FJ416657.1)ITS序列(包括ITS1、5.8S、ITS2)长度范围为604~626 bp。遵义缫丝花缺失25个碱基,无籽刺梨缺失18个碱基,光枝无籽刺梨缺失5个碱基。长尖叶蔷薇与兴仁缫丝花一样缺失3个碱基。ITS1序列长度范围为231~251 bp,其中兴仁缫丝花、长尖叶蔷薇缺失2个碱基,光枝无籽刺梨缺失3个碱基,无籽刺梨缺失6个碱基,遵义缫丝花缺失22个碱基。5.8S序列长度均为164 bp,没有变异位点。ITS2序列长度为200~211 bp,其中来自兴仁缫丝花、长尖叶蔷薇花的缺失1个碱基,光枝无籽刺梨、长尖叶蔷薇序列长度为208 bp,遵义缫丝花缺失3个碱基,无籽刺梨缺失12个碱基。排序后ITS全长为629 bp,ITS1序列长度为253 bp,5.8S序列长度为164 bp,ITS2序列长度为212 bp。
2.3形态学分析
通过观察无籽刺梨、刺梨新鲜标本与贵州缫丝花花期腊月标本,同时参考《贵州植物志》[16]上长尖叶蔷薇的形态进行比较分析,从形态上来看,无籽刺梨的形态特征与贵州缫丝花、缫丝花较为接近(表3、图1)。表3无籽刺梨及其近缘种形态学比较分析[6,15]
植物种名叶花果缫丝花椭圆形,长1.0~2.2 cm,宽0.6~1.2 cm,托叶大部分与叶柄贴生,离生部分展开花单生,稀,1~2朵簇生,花径5~6 cm,花瓣重瓣至半重瓣,粉红色至深红色果实扁球形无籽刺梨椭圆形或长卵形,长1.3~4.5 cm,宽0.5~2.2 cm,托叶附生于叶柄上,披针形花簇生,5~10朵组成伞房花序,花径 5 cm,花瓣白色果实球形,外形略像金樱子,直径约2 cm贵州缫丝花椭圆形或长卵形,长1.0~3.5 cm,宽0.7~1.8 cm,托叶下部与叶柄贴生,上部离生,呈披刺形花簇生,7~10朵组成伞房花序,花径 3 cm长叶尖蔷薇卵形,卵状长圆形,椭圆形,长3~7 cm,宽1.0~3.5 cm,托叶大部分与叶柄贴生,离生部分披针形花数朵至10余朵组成伞房花序,花径3~4 cm,白色果实倒卵球形,直径1.0~1.2 cm
2.4聚类分析
基于ITS序列,利用最大简约法构建系统树,从图2可看出,来自贵州遵义和兴仁的缫丝花与GenBank上下载的缫丝花序列聚为1支的支持率为98%,贵州铜仁金樱子与GenBank上下载的金樱子序列聚单聚为1支的支持率为75%,金樱子单聚为1支,形态学上将金樱子分在金樱子组中。无籽刺梨和光枝无籽刺梨聚为1支,支持率为99%,无籽刺梨与光枝无籽刺梨为同一种,无籽刺梨和光枝无籽刺梨与长尖叶蔷薇构成姐妹群。硕苞蔷薇单聚为1支,根据形态学分类,硕苞组中只含有硕苞蔷薇。小果蔷薇和木香花聚为1支,形态学上将小果蔷薇与木香花归为木香组。腺齿蔷薇、尾萼蔷薇和玫瑰聚为1支,形态学分类中腺齿蔷薇、尾萼蔷薇和玫瑰为桂味组,苹果为外类群,其系统发育树与形态学上分类基本一致。
2.5变异位点及碱基类型
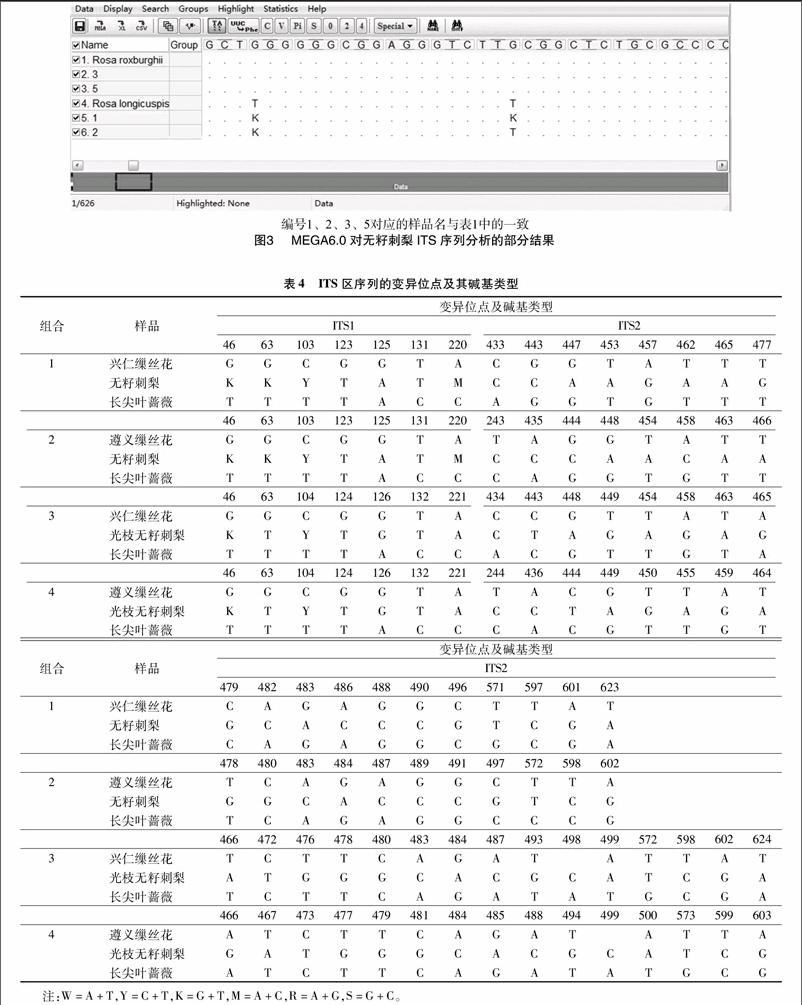
通过MEGA 6.0对序列进行分析,以无籽刺梨和光枝无籽刺梨为中心,进行组合分析变异位点、碱基类型。兴仁缫丝花、无籽刺梨、长尖叶蔷薇为第1个组合,遵义缫丝花、无籽刺梨、长尖叶蔷薇为第2个组合,贵州兴仁缫丝花、光枝无籽刺梨、长尖叶蔷薇为第3个组合,遵义缫丝花、光枝无籽刺梨、长尖叶蔷薇为第4个组合。组合1中共有26个变异位点,占ITS序列总长度的 4.10%,其中ITS1有7个变异位点,占ITS1序列长度的 2.76%,占ITS序列总长度的1.11%,5.8S中没有變异位点,ITS2中含有19个变异位点,占ITS2序列长度的8.96%,占ITS序列总长度的3.02%。组合2、组合1的变异位点数与变异比例一样。组合3中总变异位点为30个,占ITS序列总长度的4.76%,其中ITS1有7个变异位点,占ITS1序列长度的2.76%, 占ITS序列总长度的1.11%,ITS2
中有变异位点23个,占ITS2序列长度的10.8%,占ITS序列总长度的3.65%。组合4与组合3变异位点数与变异比例一样。每个组合具体的碱基变异类型见表4,其中4个组合为不同个体间的4个重复,4个组合中的ITS序列中,无籽刺梨第46个位点上发生了碱基的叠加,在这个位点上这2个碱基1个来自缫丝花,1个来自长尖叶蔷薇。由图3、表4可以看出,所有组合上的64位点,组合1、组合2上的第63、103、220位点,组合3、组合4上的第104位点都发生了碱基叠加。
3讨论与结论
光枝无籽刺梨为安明态等于2009年发表的新变种[17],同年后几个月,邓朝义等在对无籽刺梨形态分类修订中,对光
枝无籽刺梨进行归并[18]。从形态学上无籽刺梨的分类存在一定争议,基于ITS序列,从构建系统树上可见,无籽刺梨和光枝无籽刺梨聚为1支,支持率为99%,即光枝刺梨和无籽刺梨为同一个种,其结果与李旦等运用扩增片段长度多态性(AFLP)分子标记和DNA条形码对无籽刺梨的鉴定结果一致[19]。
根据孟德尔遗传学定律和植物有性生殖的特点可知,当2个亲本进行杂交时,如果2个亲本在相同位点上的等位基因序列不同,那么在子代,尤其是F1代中,将会含有这2种不同序列的等位基因。基于DNA测序仪的工作原理和测序反应的特点,当1个杂交发生后,如果2个亲本在同一基因的相同位点上的碱基不同,那么,在其杂交后代的相同基因的相同位点上将检测出2种不同的碱基,其中1个来自父本,1个来自母本,因而在原始测序图上,将呈现出这2种碱基峰的叠加[20]。由本研究可知,所有组合上的第64位点,组合1、组合2上的第63、103、220位点均发生了碱基峰的叠加,此外还有组合3、组合4上的第104位点。从本研究还可以看出,以无籽刺梨为重复,来自兴仁、遵义不同个体缫丝花为对照的组合1和组合2的变异位点及变异碱基类型基本一致,以光枝无籽刺梨为重复,来自兴仁和遵义不同个体缫丝花为对照的组合3、组合4的变异位点及变异碱基类型也基本一致。其中第46位点上4个组合中的无籽刺梨碱基都为K,均发生了碱基叠加,且叠加的碱基中1个来自缫丝花,另1个来自长尖叶蔷薇。组合1、组合2的第103位和组合3、组合4的第104位也发生相同碱基叠加,其位点不一样的原因为遵义和兴仁缫丝花不同个体间有碱基缺失,导致排序后碱基位点发生变化,无籽刺梨和光枝无籽刺梨为同一种,其中组合1和组合2中第63位上无籽刺梨为碱基叠加,而组合3、组合4中的第63位上光枝无籽刺梨碱基为T,与长尖叶蔷薇碱基一样,4个组合中缫丝花和长尖叶碱基一样。组合1、组合2上第220位点,以及组合3、组合4上的第221位点情况与第63位点一样。组合3、组合4在第63、221位点上出现此种情况的原因可能是经历了较长时间的繁衍和生境的不同,而导致光枝无籽刺梨第63位点上的G被替换成了T,第221位上的C被替换成A,ITS2中大多出现缫丝花与长尖叶碱基一致,无籽刺梨碱基不一致,如组合1中的第443位点,缫丝花和长尖叶碱基都为G,无籽刺梨却为C,其原因可能是无籽刺梨上碱基突变,G被C替换,其他变异位点也发生了替换。即无籽刺梨与缫丝花、长尖叶蔷薇ITS上的基因变化部分符合孟德尔遗传学定律。由本研究还看出,组合1和组合2的叠加碱基比组合3和组合4的多,其原因是组合1和组合2中的来自兴仁的野生无籽刺梨为原始种,采自时圣德等在贵州蔷薇属新分类群中发布的新种无籽刺梨本种采集地,位于海拔1 500 m兴仁县巴铃[6],安顺光枝无籽刺梨则经过大规模人工无性栽培繁殖,通过多次无性繁殖栽培及环境改变等都可能导致其碱基发生变化。
综上所述,无籽刺梨核基因序列与缫丝花和长尖叶蔷薇的核基因序列上部分碱基符合孟德尔遗传学定律,从用最大简约法构建的进化树来看,无籽刺梨与长尖叶蔷薇聚为1支,形态上对比可知,无籽刺梨与贵州缫丝花较为接近。文晓鹏等认为,无籽刺梨为贵州缫丝花的高度不育种[21]。贵州缫丝花仅存1份花期标本,存于贵州大学南校区博物馆内,未见果期标本,且笔者于2016年多次在贵州缫丝花采集地花溪进行搜寻采样,未发现贵州缫丝花,并对贵州缫丝花标本叶片进行DNA反复提取,由于年代久远,未提取到贵州缫丝花DNA,所以无法获取有关贵州缫丝花的分子信息,基于上述情况,无籽刺梨与贵州缫丝花的亲缘关系还有待进一步研究。
参考文献:
[1]韦景枫,程友忠,蒙先举,等. 无籽刺梨生物学特征性观察[J]. 中国林副特产,2012,121(6):27-29.
[2]韦景枫,钟漫,程友忠,等. 无籽刺梨试管苗移栽及其影响因素的探讨[J]. 中国林副特产,2010,104(1):30-31.
[3]郑元,辛培尧,高健,等. 无籽刺梨的研究与应用现状及展望[J]. 贵州林业科技,2013,41(2):62-64.
[4]林源,唐军荣,田斌,等. 无籽刺梨的研究现状及发展建议[J]. 江苏农业科学,2014,42(4):122-123,124.
[5]季祥彪,李淑久. 贵州4种刺梨的比较形态解剖学研究[J]. 山地农业生物学报,1998,1(1):28-33.
[6]时圣德. 贵州蔷薇属新分类群[J]. 贵州科学,1985(1):11-12.
[7]邓亨宁,高信芬,李先源,等. 无籽刺梨杂交起源:来自分子数据的证据[J]. 植物资源与环境学报,2015,24(4):10-17.
[8]徐炳声,顾德兴. 杂交在进化中的作用及杂种的识别和分类处理[J]. 武漢植物学研究,1986,4(4):385-397.
[9]Arnold M L,Bouck A C,Cornman R S. Verne Grant and Louisiana Irises:is there anything new under the sun[J]. New Phytol,2003,161(1):143-149.
[10]Soltis P S,Doyle J J,Soltis D E. Molecular data and polyploid evolution in plants [M]. US:Springer,1992:177-201.
[11]罗瑜萍,龚维,邱英雄,等. 羊蹄甲属3种园艺树种分子鉴定及亲缘关系的ISSR分析[J]. 园艺学报,2006,33(6):433-436.
[12]Baldwin B G. Phylogenetic utility of the internal transcribed spacers of nuclear ribosomal DNA in plants:an example from the compositae[J]. Molecular Phylogenetics and Evolution,1992,1(1):3-16.
[13]Baklwin B G,Sanderson M J. Porter J M,et al. The ITS region of nuclear ribosomal DNA:a valusble source of evidence on angiosprem phyloeny[J]. Annals of the Missouri Botanical Garden,1995,82(2):247-277.
[14]盧松茂,陈振东,林秀香,等. 基于rDNA-ITS序列的天门冬拟茎点霉与相似种的系统发育关系[J]. 江苏农业学报,2015,31(1):62-67.
[15]Gurushidze M,Mashayekhi S,Blattner F R,et al. Phylogenetic relationships of wild and cultivated species of Allium section Cepa inferred by nuclear rDNA ITS sequence analysis[J]. Plant Systematics and Evolution,2007,269(3/4):259-269.
[16]李永康,黄威廉,王兴国,等. 贵州植物志[M]. 成都:四川民族出版社,1984:235-247.
[17]安明态,程友忠,钟漫,等. 贵州蔷薇属一新变种——光枝无籽刺梨[J]. 资源与利用,2009,1(42):63.
[18]邓朝义,方仕能,黄勇. 贵州特有种子植物无子刺梨形态特征研究及分类学订正[J]. 种子,2009,28(9):62,68.
[19]李旦,周安佩,张德国,等. 基于AFLP分子标记和DNA条形码对无籽刺梨的鉴定[J]. 林业科学研究,2015,28(1):116-121.
[20]袁长春,黎培新,王燕芳,等. 用核糖体ITS区序列验证自然杂交种Meconopsis× cookei G. Taylor[J]. 遗传学报,2004,31(9):901-907.
[21]文晓鹏,庞晓明,邓秀新. 刺梨及部分近缘种形态学性状及RAPD标记分析[J]. 园艺学报,2003,30(2):204-206.伍革民,徐龙鑫,朱丽莉,等. 瑶山鸡TRHR基因编码区序列SNP检测及其与繁殖性状的相关性[J]. 江苏农业科学,2017,45(17):47-49.
