
45份菜用大黄的SRAP标记引物筛选及亲缘关系分析
2017-11-15 02:28邵珠田姜立娜郭小菲蔡祖国赵一鹏
江苏农业科学 2017年17期
邵珠田+姜立娜+郭小菲+蔡祖国+赵一鹏

摘要:以45份引种的菜用大黄遗传背景未知为出发点,利用已报道的相关序列扩增多态性(sequence-related amplified polymorphism,简称SRAP)引物,对这些引种材料进行遗传多样性分析,旨在阐明这些材料的亲缘关系。结果表明:在选用的182对引物组合中有42对引物扩增产物稳定、条带清晰、多态性较好,可以用作菜用大黄遗传多样性分析;利用42对引物对45份材料扩增后共获得230个等位基因位点,其中多态性位点202个,平均多态性位点比例达到878%。基于非加权组平均法(unweighted pair-group method with arithmetic means,简称UPGMA)的聚类分析结果表明:45份菜用大黄材料聚为7类,虽然来源地相同的大部分种群可以聚在同一个类群内,但种群间存在遗传交叉现象,揭示了种群间的亲缘关系,为这些引入材料的进一步利用提供了参考。
关键词:菜用大黄;SRAP分子标记;引物筛选;亲缘关系
中图分类号: S644.903文献标志码: A文章编号:1002-1302(2017)17-0039-03
通信作者:赵一鹏,博士,教授,主要从事园艺植物种质资源与育种研究。E-mail:yzhao36@163.com。菜用大黄(Rheum rhaponticum L.)别称食用大黄、圆叶大黄、酸菜等,为蓼科(Polygonaceae)大黄属(Rheum)中以叶柄为蔬菜的栽培种,英文统称为Rhubarb。菜用大黄属于多年生草本植物,其叶柄具有粗大多汁、营养丰富、气味清新芬芳、风味独特、口感微酸等特点,是理想的芳香保健类蔬菜。在国外,菜用大黄被广泛用于制作馅饼、果酱、果冻、面包、蛋糕、沙拉、大黄果酒等各类甜品及加工成高级果蔬汁饮料[1-4]。
目前,国内对大黄属植物的研究和开发利用仅限于药用大黄相关资源,对菜用大黄的系统研究较少。2004年以来,河南科技学院菜用大黄引种及种质资源利用课题组赵一鹏博士在与国外合作的基础上,首次开展了菜用大黄优良种质资源的引种、组织培养技术[3-6]、栽培学特性及营养成分分析[7]、根尖染色体观察及核型分析[8]等研究,但关于菜用大黄起源、资源亲缘关系的研究尚未开展。本研究在进行了相关序列扩增多态性(sequence-related amplified polymorphism,简称SRAP)体系优化的基础上[9],利用多态性高的42对引物组合对从国内外收集到的45份菜用大黄材料进行亲缘关系分析,旨在了解这些菜用大黄材料的遗传背景,为今后菜用大黄优良品种选育和分子生物学研究提供一定的技术支持。
1材料与方法
1.1 材料
用于引物筛选的菜用大黄材料为样本4号,种子来源于美国。用于亲缘关系分析的45份材料,采自河南科技学院菜用大黄资源圃,其中从美国引种材料13份,分别编号为1~13;从英国引种材料20份,分别编号为14~28、41~45;从北京引种材料12份,分别编号为29~40。于2016年4月上旬采集新鲜、无病虫害的叶片,用清水冲洗干净,置于-80 ℃冰箱中保存备用。
1.2方法
1.2.1模板DNA的提取及质量检测基因组DNA提取参照Liu等改良十六烷基三甲基溴化铵(CTAB)法[10],用Eppendorf核酸蛋白测定仪(德国)检测所提DNA的纯度及浓度,并用无菌ddH2O稀释至10 ng/μL,-20 ℃保存备用。
1.2.2SRAP-PCR引物的筛选SRAP-PCR扩增程序参照陈万胜等的方法[11],即94 ℃ 5 min;94 ℃ 45 s,35 ℃ 45 s,72 ℃ 1 min,5个循环;94 ℃ 45 s,52 ℃ 45 s,72 ℃ 1 min,35个循环;72 ℃ 10 min。PCR产物于4 ℃保存备用。菜用大黄SRAP-PCR反应体系为笔者所在课题组郭小菲等已优化的体系[9],即25μL体系中包含2.50 μL 10×PCR Buffer、30 ng模板DNA、2.50 mmol/L Mg2+、0.25 mmol/L dNTPs、0.04 U/μL Taq DNA聚合酶,各0.50 μmol/L上下游引物,剩余体积用双蒸水补足。以上试剂均购自TaKaRa公司。引物序列参考Li等的报道[12-15],共选用14条正向引物、13条反向引物(表1),可组合成182对引物。利用4号材料对上述引物进行筛选。依据电泳结果,选择扩增产物多态性丰富、条带清晰、重复性好的引物组合进行菜用大黄的亲缘关系研究。
1.2.3PCR扩增产物的检测扩增反应结束后,在产物中加入5 μL 6×Loading Buffer,混匀,取2.5 μL混合液上样于6%非变性聚丙烯酰胺凝胶中。预电泳电压90 V,时间 30 min;电泳电压120 V,时间3 h。电泳结束后的胶板采用袁菊红等的银染法[16]进行染色。
1.2.4数据统计与分析根据电泳结果,在凝胶相同迁移位置上,扩增有条带记为“1”,无条带记为“0”,最终组成1个由45份材料和所有扩增位点等位基因组成的“1,0”矩阵。利用NTsys-pc Version 2.10e 软件,采用非加权组平均法(unweighted pair-group method with arithmetic means,简称UPGMA)
2结果与分析
2.1SRAP-PCR引物的筛选及扩增产物的多态性分析
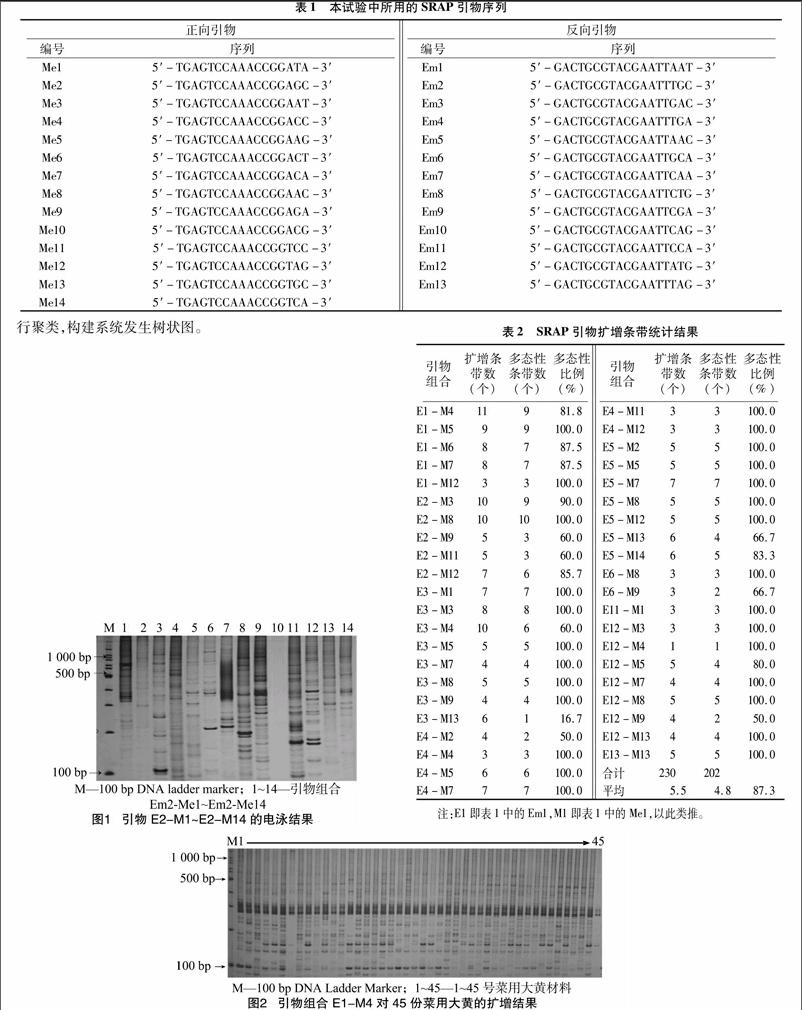
从182对引物中共筛选出扩增產物重复性好、多态性高和条带清晰的引物组合42对,图1为部分引物筛选扩增结果。应用筛选的42对多态性引物组合对45份材料模板DNA样品进行PCR扩增,结果共获得230个等位基因位点,其中202个为多态性位点。每个引物组合的多态性比率在16.7%~1000%之间,平均多态性比例达到87.8%;平均每对引物扩增条带数5.5个,平均多态性条带数4.8个;在42对引物中,引物组合E1-M4扩增条带数最多,共11个,E12-M4组合扩增条带数最少,只有1个(表2)。引物组合E1-M4的扩增结果如图2所示。
2.2菜用大黄遗传相似性与聚类分析
45份样品经NTSYS软件聚类分析结果显示,在遗传相似系数0.68处可将45份样品聚为7类,第1类是34号、35号,种子来源于北京;第2类是25号、26号、27号、28号,种子来源于英国;第3类为36号、37号,种子同样来源于北京;第4类有13号、15号、38号、39号、40号、42号6个样品;第5类为2号、3号、4号、8号、10号、11号、12号和24号共8个样品,除24号来源于英国外,其余均引种于美国;1号单独聚为1类,引种地为美国,其余22个样品聚为一大类;45份样品间的遗传相似系数在0.514 9~0.811 9之间,遗传距离在0.117 1~0.703 0之间(图3)。
3讨论
形态标记是植物分类学上最古老、最简便易行的方法,但是由于数量性状的遗传机制复杂、遗传力低、容易受到环境因素的影响、工作量大等因素,该方法不能完全满足亲缘关系研究的需要。SRAP标记是一种基于PCR的DNA分子标记技术,具有操作简单、引物通用性与多态性高、重复性好、标记位点在基因组中分布均匀等优点[18]。近年来利用SRAP标记技术进行品种鉴定,分析种质间的遗传多样性、品种间的亲缘关系等已经有了较多报道[19-21]。但在关于大黄属植物的研究中, 仅见陈大霞等利用SRAP标记对正品大黄的遗传关系进行了分析,其研究结果表明,3种正品大黄间的SRAP的多态性比例为73.2%[22]。而本研究中菜用大黄多态性比例为87.8%,多态性相对较高,笔者认为与本研究的供试材料来源地有关,因为45份菜用大黄材料来源于北京、美国、英国等不同地区,遗传基础相差较远,导致变异位点较多。从本试验结果来看,SRAP分子标记可以成为一种评价种质资源亲缘关系的新技术。
运用SRAP分子标记技术对45份菜用大黄材料进行亲缘关系研究,从遗传相似系数和聚类树状图来看,个体间存在较大的差异。在遗传相似系数0.68处可将供试的45份菜用大黄样品聚为7类,第1类是34号、35号,种子来源于北京,结果期种子均呈现黄绿色,叶柄表面有短毛、粗糙且斑点密布;第2类是25号、26号、27号、28号,种子来源于英国,叶柄基部均呈现深粉色,表面较为光滑;第3类为36号、37号,引种于北京,结果期种子翅缘均呈现粉红色。虽然来源地相同的大部分样品可以聚在同一个类群内,但仍出现了交叉聚类现象。24号种子来源于英国,却与来源于美国的2号、3号、4号、8号、10号、11号、12号种子聚在了第5类;13号、15号种子又与来源于北京的38号、39号、40号种子聚在了第4类。出现这种情况的原因可能是由于长期的选择进化及不断的相互引种,导致各生态地理群之间种质的相互渗透。本研究通过SRAP标记技术对收集到的45份国内外菜用大黄材料进行了初步的亲缘关系分析,为今后进行菜用大黄遗传育种和品种鉴定提供了一定的技术支持。
参考文献:
[1]卢莉,赵一鹏. 菜用大黄的研究进展[J]. 广东农业科学,2008(2):19-21,27.
[2]Zhao Y P. Somaclonal variation and crop failure of micro-propagated rhubarb[D]. Colchester,UK:University of Essex,2004.
[3]卢莉,赵一鹏,蔡祖国,等. 欧洲大黄种子发芽特性研究[J]. 种子,2007,26(3):35-37.
[4]赵一鹏,Grout B W,周岩. 欧洲大黄茎尖组织培养与快速繁殖[J]. 河南职业技术师范学院学报,2004,32(3):24-25,28.
[5]卢莉. 菜用大黄的种子萌发特性与离体快繁技术研究[D]. 新乡:河南师范大学,2008.
[6]张有铎,蔡祖国,赵一鹏. 菜用大黄组培苗生根特性研究[J]. 湖北农业科学,2009,48(11):2759-2761.
[7]吴亚蓓. 菜用大黄栽培学特性与营养成分分析[D]. 新乡:河南科技学院,2012.
[8]任文娟,郭小菲,姜立娜,等. 菜用大黄染色体制片优化及核型分析[J]. 华北农学报,2013,28(5):128-132.
[9]郭小菲,姜立娜,蔡祖国,等. 菜用大黄 SRAP-PCR 反应体系的
优化及验证[J]. 华北农学报,2014,29(4):105-110.
[10]Liu L,Guo W,Zhu X,et al. Inheritance and fine mapping of fertility restoration for cytoplasmic male sterility in Gossypium hirsutum L.[J]. Theoretical and Applied Genetics,2003,106(3):461-469.
[11]陈万胜,王元英,罗成刚,等. 利用正交设计优化烟草SRAP反应体系[J]. 分子植物育种,2008,6(1):177-182.
[12]Li G,Quiros C F. Sequence-related amplified polymorphism (SRAP),a new marker system based on a simple PCR reaction:its application to mapping and gene tagging in Brassica[J]. Theoretical and Applied Genetics,2001,103(2/3):455-461.
[13]Ferriol M,Picó B,Nuez F. Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers[J]. Theoretical and Applied Genetics,2003,107(2):271-282.
[14]林忠旭,张献龙,聂以春. 新型标记SRAP在棉花F2分离群体及遗传多样性评价中的适用性分析[J]. 遗传学报,2004,31(6):622-626.
[15]王和勇,乔爱民,陈敏. 植物遗传标记的发展及应用[J]. 仲恺农业技术学院学报,2000,13(4):58-64.
[16]袁菊红,权俊萍,胡绵好,等. 石蒜SRAP-PCR扩增体系的建立与优化[J]. 植物资源与环境学报,2007,16(4):1-6.
[17]尹跃. 枸杞品种(系)分子鉴定及亲缘关系分析[D]. 福州:福建农林大学,2013.
[18]刘雅辉,王秀萍,鲁雪林,等. 棉花耐盐相关序列扩增多态性(SRAP)分子标记筛选[J]. 江苏农业学报,2015,31(3):484-488.
[19]沈国正,李春楠,傅巧娟,等. 一串红SRAP标记的建立與品种鉴定[J]. 浙江农业学报,2011,23(1):84-89.
[20]颜璐茜,李佳蔓,员涛,等. 滇杨遗传多样性的SRAP分析[J]. 生物技术通报,2016,32(4):159-167.
[21]肖政,苏家乐,刘晓青,等. 杜鹃花种质资源遗传多样性的SRAP分析[J]. 江苏农业学报,2016,32(2):442-447.
[22]陈大霞,李隆云,钟国跃,等. 用SRAP标记分析正品大黄的遗传关系[J]. 中国中药杂志,2008,33(20):2309-2312.陈兴银,石建明,杨鹏,等. 无籽刺梨及其近缘种ITS序列分析[J]. 江苏农业科学,2017,45(17):42-46.
