
陕西子午岭国家级自然保护区鞘翅目昆虫群落的边缘效应
2017-11-10 03:11徐世才王丽娟
生态科学 2017年5期
徐世才, 王丽娟
延安大学生命科学学院, 陕西延安716000
陕西子午岭国家级自然保护区鞘翅目昆虫群落的边缘效应
徐世才*, 王丽娟
延安大学生命科学学院, 陕西延安716000
选取陕西子午岭国家级自然保护区阔叶林—针叶林、农田—灌丛和草地—灌丛3种边缘地带为研究样地, 在科级水平上探讨边缘效应对鞘翅目昆虫群落多样性的影响。共采集鞘翅目昆虫4935只, 隶属于28科, 其中步甲科、叶甲科和瓢虫科的个体数量分别占个体总数的44.19%、21.95%和13.66%, 构成该地区鞘翅目昆虫的优势类群。3种边缘类型中鞘翅目昆虫的科多样性、丰富度和均匀度均沿边缘向相邻生境内部降低,EH´值均大于 1, 呈现边缘正效应。经多元回归分析表明, 植被密度影响阔叶林—针叶林边缘鞘翅目昆虫群落的多样性和丰富度, 植被密度和盖度影响整个群落鞘翅目昆虫的个体数量分布。以上结论可为陕西子午岭自然保护区鞘翅目昆虫生物多样性的保护和合理利用提供科学依据。
鞘翅目; 群落多样性; 边缘效应
1 前言
频繁的人类活动和自然灾害的发生, 加速了生境的破碎化与生物栖息地的丧失。片段化的森林代替了原有大面积的自然生境, 形成不同的斑块与边缘。边缘效应的存在使得森林内部到外部的生态环境产生梯度变化, 包括栖息地、食物、繁殖场所等[1]。生物由于自身及外界环境等因素的影响, 对边缘产生不同的反应, 研究物种对边缘的选择趋势或行为反应对边缘效应的理解与生物多样性的保护具有特定的价值。鞘翅目(Coleoptera)昆虫是昆虫纲中最大的一个目, 占全球已知昆虫总数的25%[2]。鞘翅目昆虫食性多样, 在整个生态系统中扮演着重要的角色[3],且在维持生物多样性等方面具有重要作用, 其中部分类群还常被选作指示性昆虫来反映环境的变化情况[4–5], 对其的研究也逐渐受到国内外学者的重视。
由于人为的破坏, 如乱砍滥伐和过度开垦等,子午岭森林由相对完整逐渐走向破碎化, 形成了片段化的景观和天然次生林[6]。随着人们逐渐认识到子午岭在维持黄土高原地区生态稳定方面的重要性,2006年成立了陕西子午岭国家级自然保护区, 促进了该保护区动、植物多样性的保护[7], 生物多样性也得到逐步恢复, 但生态恢复过程中生物多样性的恢复状况缺乏相应的监测与评价。本文选取子午岭阔叶林—针叶林、农田—灌丛和草地—灌丛3种边缘类型, 以鞘翅目昆虫为研究对象, 通过比较不同边缘鞘翅目昆虫在科级水平上的群落多样性及其边缘效应, 分析影响鞘翅目昆虫分布的植被特征, 研究子午岭在生态恢复过程中生境片段化对鞘翅目昆虫多样性的影响, 以期为子午岭鞘翅目昆虫生物多样性的保护及合理利用, 子午岭保护区植被恢复与重建提供参考。
2 研究地区与研究方法
2.1 研究区概况
子午岭国家级自然保护区位于陕西省富县境内,南与渭北大平原相接, 西靠子午岭主脊, 北临毛乌素沙漠边缘, 地理坐标在东经 108°29′—108°41′, 北纬 35°45′—36°01′之间, 总面积 40621 hm²。保护区气候属暖温带半湿润气候, 年平均气温7.5—8.9 ℃,昼夜温差大, 年降水量520—610 mm, 南多北少。植被类型是从森林草原—半干早草原的过渡地带, 地带性植被为华北暖温带落叶阔叶林地带的北部落叶阔叶林。依据《中国植被》分类原则, 保护区的植被可分为森林、灌丛和草地3种植被类型, 共20个群系, 主要树种有辽东栎(Quercus wutaishanica)、油松(Pinus tabulaeformis)、山杨(Populus davidiana)、小叶杨(Populus simonii)、白桦(Betula platyphylla)和侧柏(Platycladus orientalis)等。此外还有大果榆(Ulmus macrocarpa)、野杏(Armeniaca vulgaris)、小叶朴(Celtis bungeana)、茶条槭(Acer ginnala)、白蜡树(Fraxinus chinensis)、北京丁香(Syringa pekinensis)等华北树种,常以小片状分布或与主要树种伴生或散生。林下及散生灌木主要有狼牙刺(Sophora viciifolia)、沙棘(Hippophae rhamnoides)、虎榛子(Ostryopsis davidiana)、文冠果(Xanthoceras sorbifolium)、连翘(Forsythia suspense)、胡枝子(Lespedeza bicolor)、土庄绣线菊(Spiraea pubescens)等。草本植物主要有菊科(Compositae)、禾本科(Gramineae)、苔草(Carex)、莎草(Cyperus)等。此外, 保护区内还存有农田和苗圃, 主要种植玉米(Zea mays)和牡丹(Paeonia suffruticosa)。
2.2 调查方法
根据陕西子午岭保护区的自然条件, 以保护区内槐树庄保护站至榆林站为主线, 选取阔叶林—针叶林、农田—灌丛和草地—灌丛3种类型的边缘地带进行研究, 依据采捕的种类及生境的具体情况, 利用网捕法、直接搜捕法和诱捕法等采捕样地内的鞘翅目昆虫并制作标本, 依据相关文献专著鉴定[8–11]并记录鞘翅目昆虫的种类和数量。采集工作在 2016年5—9月进行, 每月中旬采一次样。
参考 Meiners和 Pickett[12]及 Heliölä等[13]研究边缘效应的方法, 以距离梯度形式研究边缘及相邻生境内部鞘翅目昆虫群落的差异。设计样点时, 在与边缘垂直方向上设置5条样带, 样带间隔为10 m, 每条样带以边缘为中心, 向相邻生境内部延伸100 m, 以每隔25 m的距离设置样点, 大小为2 m×5 m, 具体取样方法见图1(示两条样带)。
为研究鞘翅目昆虫分布与植被特征之间的关系,统一在调查中期(2016年7月)调查记录各个样带(以样点中央为中心, 直径2 m范围内)的植被种类、密度、盖度和凋落物厚度等特征。
2.3 数据处理与分析
以鞘翅目昆虫在科级水平上的丰富度、个体数量、多样性、均匀度及优势度为数据分析对象, 将每种边缘类型的 45个研究样点依据与边缘的距离聚为3组: 生境1包括A、B和C(分别距边缘100 m、75 m和50 m)3条样地带, 边缘包括D、E和F(分别距边缘25m、0m和25m)3条样地带, 生境2包括余下的G、H和I(分别距边缘50 m、75 m和100 m)3条样地带。
科多样性分析主要选用 4类指数, 即 Shannon-Wiener多样性指数、Margalef丰富度指数、Pielou均匀度指数和Simpson优势度指数, 计算公式如下[14–15]:
Shannon-Wiener多样性指数(H´)H´=-∑PilnPi
Margalef丰富度指数(D)D=(S-1)/lnN
Pielou均匀度指数(J)J=H´/lnS
Simpson优势度指数(C)C=∑(ni/N)2
式中Pi=ni/N,Pi为第i科占总个体数的比率,ni是第i科的个体数,N是个体总数,S表示群落科数。
边缘效应强度(E)根据测度模型:E=mY/∑yi, 令由m个群落组成的交错区的这一指标为Y,m个群落的这一指标为yi(i=1, 2, 3, …,m)。再以 Shannon-Wiener多样性指数(H´)和Simpson优势度指数(C)这两个定量指标去拟合上述公式, 则可得出边缘效应值EH'和EC:
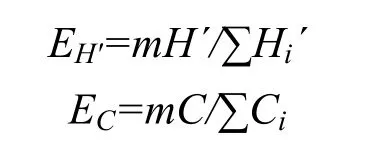
其中H´为群落组成的交错区鞘翅目昆虫科的多样性指数;Hi´为各个群落里鞘翅目昆虫科的多样性指数;C为群落组成的交错区内鞘翅目昆虫科的优势度指数;Ci为各个群落里鞘翅目昆虫科的优势度指数[16]。
利用Excel 2016和SPASS 19.0对实验数据进行分析处理, 通过多元线性逐步回归分析检验鞘翅目昆虫科的多样性、丰富度、均匀度以及优势度与植被特征之间的关系。
3 结果与分析
3.1 研究地鞘翅目昆虫群落组成分析
调查时间内共捕获鞘翅目昆虫4935号, 分属于24 科(表1)。其中步甲科、叶甲科和瓢虫科的个体数量最多, 分别占个体总数的 44.19%、21.95%和13.66%, 均大于 10%, 为该地区的优势类群; 叩甲科、天牛科、金龟科、拟步甲科和芫菁科的个体数量在 1%—10%之间, 为该地区的常见类群; 虎甲、锹甲、象甲、丽金龟、花金龟、蜣螂、隐翅虫、小蠹虫、豆象、吉丁甲、犀金龟、皮金龟、蜉金龟、葬甲、郭公虫和萤科等 16科个体数量少于1%, 为该地区较不常见类群。
3.2 不同边缘距离鞘翅目昆虫的分布
在科级水平上, 通过比较阔叶林—针叶林边缘、农田—灌丛边缘和草地—灌丛边缘不同边缘距离鞘翅目昆虫的个体数量与丰富度(图 2, 3)发现, 3种边缘类型中鞘翅目昆虫的个体数量均沿边缘分别向相邻生境内部降低, 且整体上鞘翅目昆虫的个体数量以阔叶林—针叶林边缘>草地—灌丛边缘>农田—灌丛边缘, 阔叶林>针叶林>灌丛>草地>农田。阔叶林—针叶林边缘鞘翅目昆虫的丰富度沿边缘向阔叶林内部先升高后降低, 沿边缘向针叶林内部降低; 农田—灌丛边缘鞘翅目昆虫的丰富度变化趋势与阔叶林—针叶林边缘大致相同, 沿边缘向农田内部先升高后降低, 向灌丛内部降低;草地—灌丛边缘鞘翅目昆虫的丰富度沿边缘分别向草地和灌丛内部降低, 且整体上以灌丛内部的丰富度高于草地。
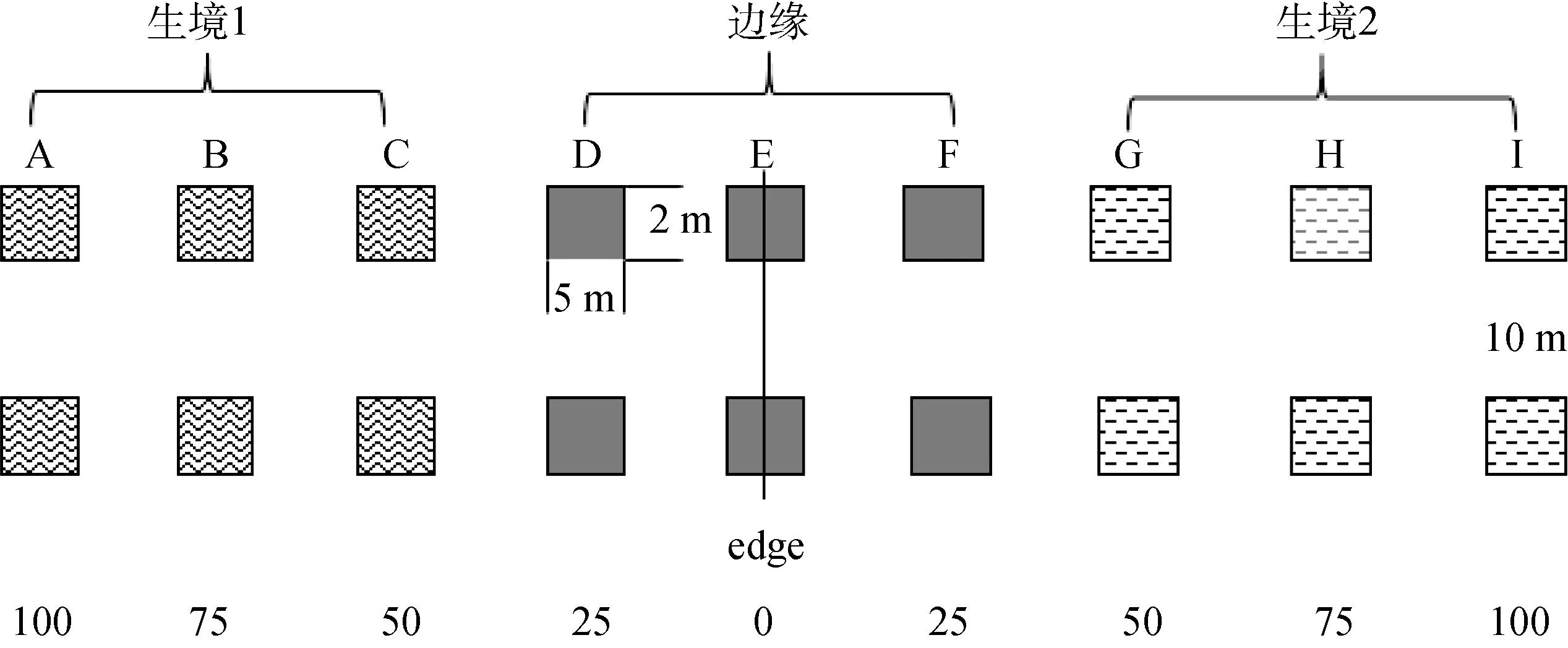
图1 样地设置Fig.1 The sample setting
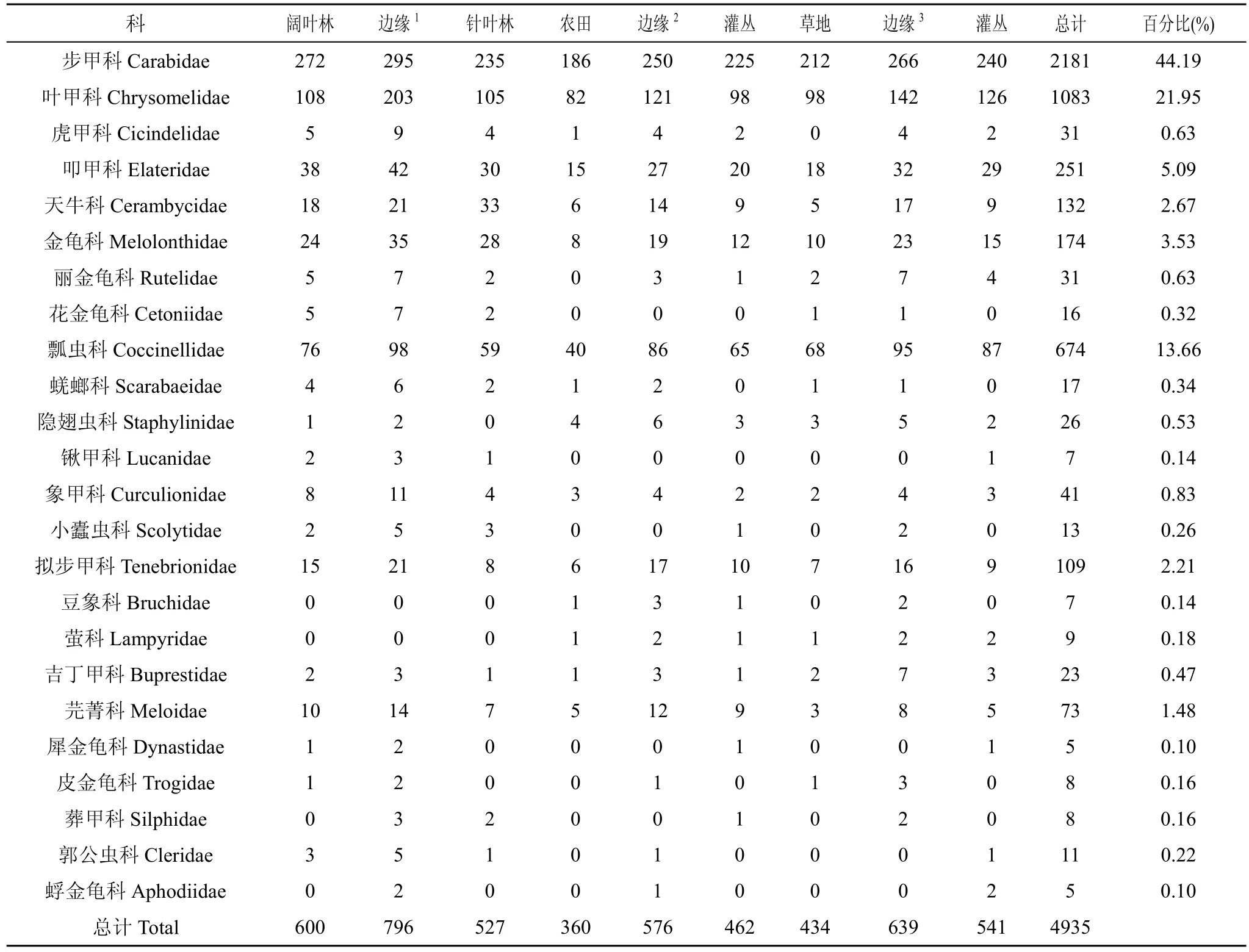
表1 各边缘类型鞘翅目昆虫的组成与数量分布Tab.1 Composition and quantity of Coleoptera insects in different edges
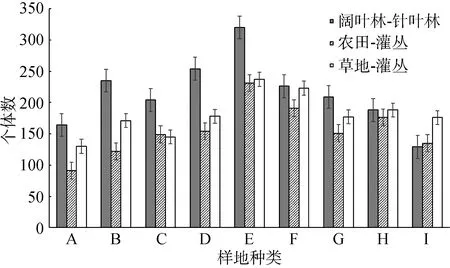
图2 不同边缘距离鞘翅目昆虫个体数量Fig.2 Individuals of Coleoptera insects in different transect
3.3 不同边缘类型鞘翅目昆虫边缘效应分析
本调查涉及的3种边缘及其相邻生境中鞘翅目昆虫群落多样性指数见表2。边缘地带鞘翅目昆虫的Shannon-Wiener多样性、丰富度和均匀度均高于其相邻生境, 总体上各多样性指数的排列顺序为:阔叶林—针叶林边缘>草地—灌丛边缘>农田—灌丛边缘。
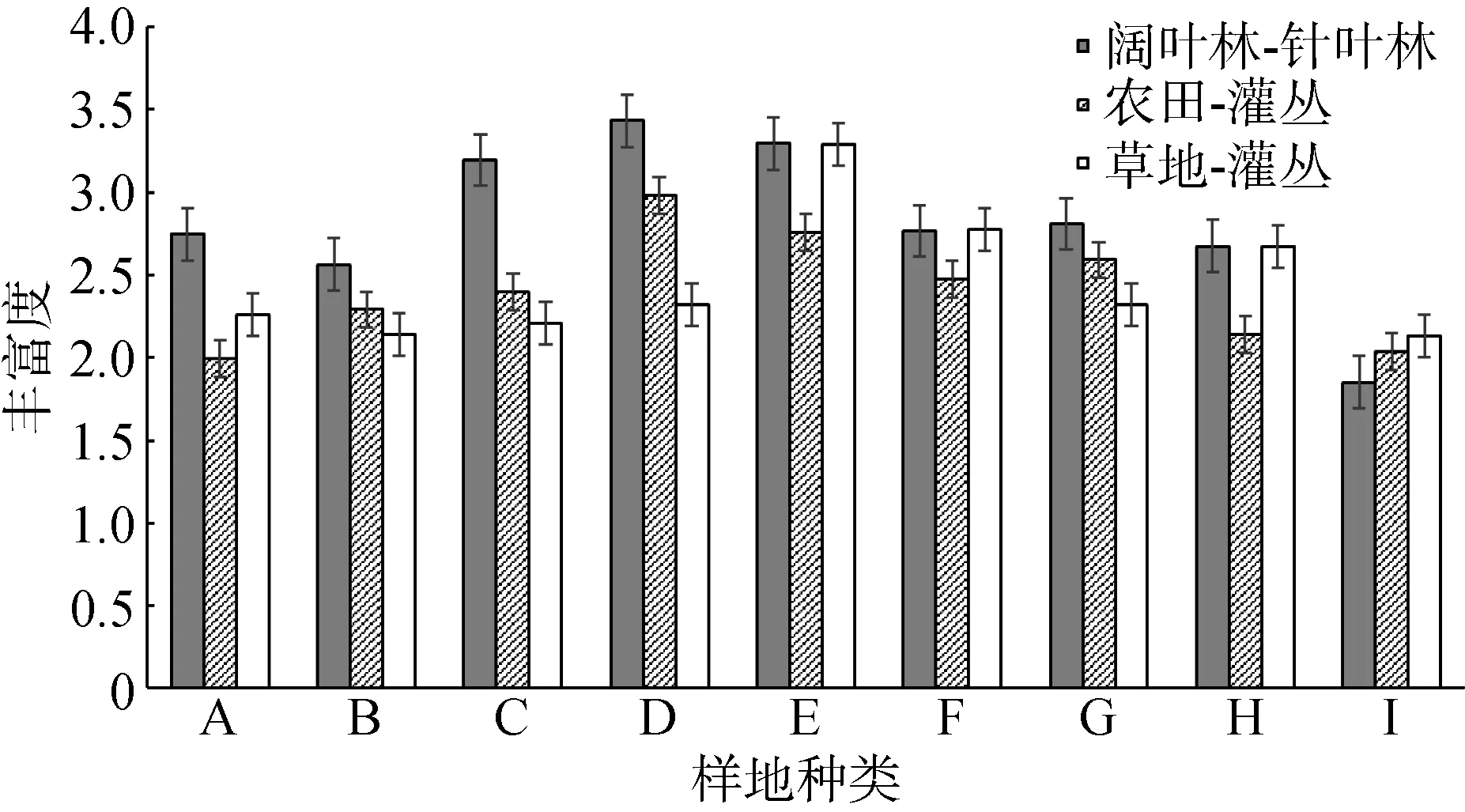
图3 不同边缘距离鞘翅目昆虫科丰富度Fig.3 Family richness of Coleoptera insects in different transect
在阔叶林—针叶林边缘中, 鞘翅目昆虫的Shannon-Wiener多样性与丰富度变化趋势相同, 总体上以阔叶林—针叶林边缘>阔叶林>针叶林, 但均匀度与优势度在阔叶林与针叶林中基本相等。农田—灌丛边缘鞘翅目昆虫的Shannon-Wiener多样性与丰富度以边缘处最高, 且灌丛内部大于农田, 均匀度农田内部稍大于灌丛, 优势度的变化趋势与Shannon-Wiener多样性和丰富度相反。草地—灌丛边缘, 鞘翅目昆虫群落的Shannon-Wiener多样性、丰富度和均匀度的变化趋势为: 草地—灌丛边缘>灌丛>草地, 与优势度的变化趋势相反。各生境鞘翅目昆虫群落的 Shannon-Wiener多样性排列顺序为:阔叶林>针叶林>灌丛>农田>草地, 林地因其拥有丰富的植被类型与较高的生产力, 同时受人为干扰少, 所以鞘翅目昆虫群落 Shannon-Wiener多样性与丰富度高; 农田由于其特殊的生境, 种植类型单一, 且受到的人为干扰较大, 具有较低的群落丰富度, 但由于其内分布的鞘翅目昆虫种类虽少但个体数量较多, 且分布均匀, 因此其多样性指数H´并不低。
边缘效应强度可对边缘效应的强弱进行定量测量, 据王伯荪等人所组建的测度模型[16]的生态学意义, 较大的EH´值表示具有较高的边缘效应强度, 当EH´>1时, 可认为呈正的边缘效应, 当EH´<1时则呈负的边缘效应。对于EC值, 则较高的边缘效应强度具有较低的EC值, 故在测度时EC值可变换为倒数。由边缘效应强度分析结果(表3)得出, 阔叶林—针叶林边缘、农田—灌丛边缘和草地—灌丛边缘的EH´值均大于1, 表明这3种边缘类型均呈正的边缘效应作用。阔叶林—针叶林边缘地带由于两相邻生境植被类型丰富, 各环境因子的分配差异较小, 鞘翅目昆虫的种类和数量分布均匀, 因此具有较高的群落多样性, 导致其EH´值低于农田—灌丛边缘和草地—灌丛边缘。用EC值来测定也能得到相同的结果, 即 3种边缘类型均呈正的边缘效应作用。
3.4 鞘翅目昆虫群落多样性与植被特征的相关性
对鞘翅目昆虫的科多样性、丰富度、均匀度、优势度和个体数量与植物群落多样性、植被密度、植被盖度和凋落物厚度这4种植被特征进行多元线性回归检验, 只有阔叶林—针叶林边缘鞘翅目昆虫群落的多样性指数与植被特征有显着的回归关系(表4)。植被密度是影鞘翅目昆虫 Shannon-Wiener多样性(r2=0.511,F=7.311,P=0.030)和丰富度(r2=0.710,F=17.169,P=0.004)的决定因素, 但植被特征对鞘翅目昆虫群落的科均匀度和优势度影响不大, 没有显着的回归关系; 植被密度和盖度是影响鞘翅目昆虫总个体数量分布的重要因素(r2=0.845,F=16.387,P=0.004); 对3个优势科进行的回归分析表明, 叩甲的个体数量主要受植被密度的影响(r2=0.566,F=9.129,P=0.019), 瓢虫的个体数量主要由植被密度与盖度决定(r2=0.817,F=13.423,P=0.006), 而布甲的个体数量与植被特征之间无显着的回归关系, 反映出不同类群对植被特征的反应存在差异。
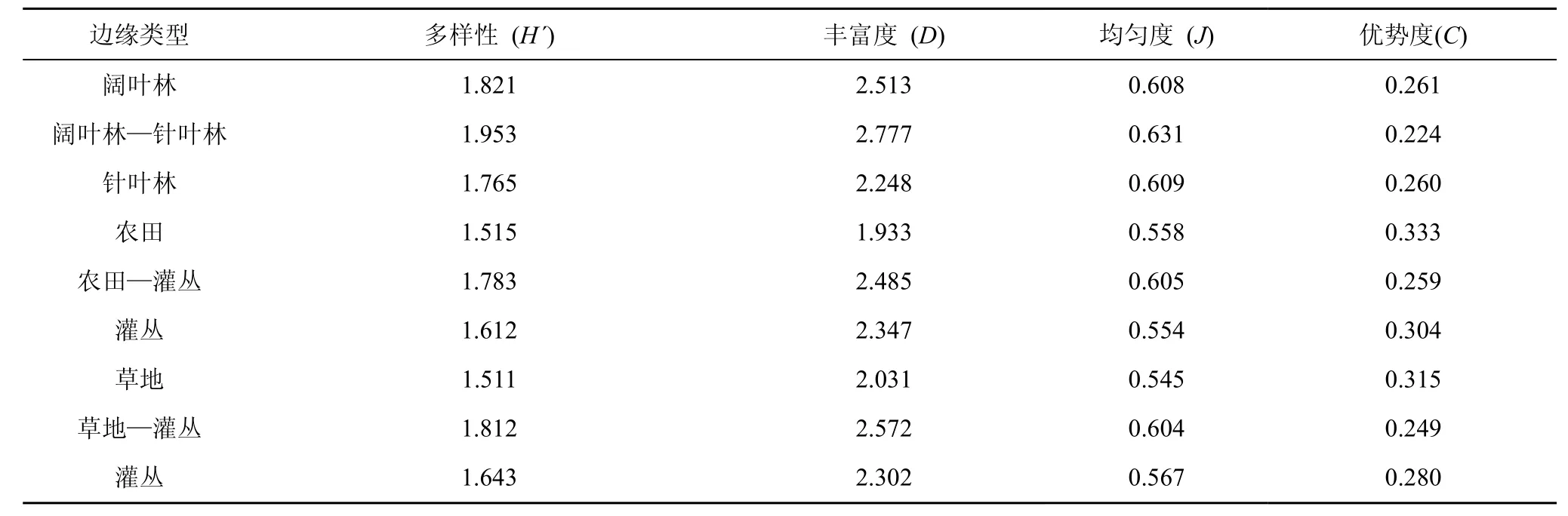
表2 不同边缘类型鞘翅目昆虫群落多样性Tab.2 Comparison of Coleoptera insects community diversity indices among different type edges
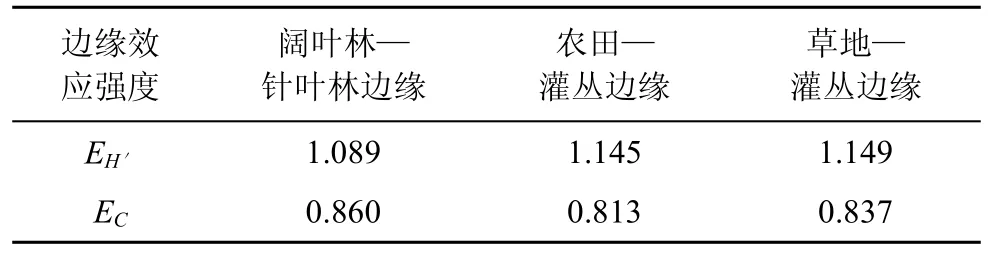
表3 不同边缘的边缘效应强度比较Tab.3 Comparison with the edge effect strength of different edges
4 讨论
在科级水平上, 阔叶林—针叶林、农田—灌丛和草地—灌丛3种边缘类型中鞘翅目昆虫群落的丰富度与个体数量整体上表现为沿边缘分别向相邻生境内部降低的趋势, 则对大部分鞘翅目昆虫来说,能够适应边缘复杂的环境, 在边缘很好地生存和繁殖, 但各边缘的微环境与资源状况不同, 使得鞘翅目昆虫的种类和数量分布存在差异。
本文调查的 3种边缘地带鞘翅目昆虫群落的Shannon-Wiener多样性、丰富度和均匀度均高于其相邻生境, 总体上各生境内鞘翅目昆虫群落的多样性指数排列顺序为: 阔叶林>针叶林>灌丛>草地>农田。对比刘云慧等人研究坝上地区农田和恢复生境地表甲虫多样性的结果, 农田较禁牧草地和再造林地其鞘翅目科数及步甲物种数都更多, 表现出草地禁牧和再造林的生境修复活动在当前状态下对生物多样性恢复的作用尚不明显[17], 此差异可能说明子午岭保护区在其生境恢复过程中生物多样性得到较好的恢复,但对于评估生境恢复在多样性恢复中所起的作用,需要建立在长期监测的基础上[18], 对此我们还需要进行长期深入地调查研究, 以期为子午岭保护区的生态恢复提供更合理有效的措施, 提高生物多样性恢复的效率。
边缘效应强度EH´值均大于1, 说明3种边缘类型均呈正的边缘效应作用。这与王巍巍等人[4]研究荒漠景观地表甲虫群落的边缘效应有正效应和负效应之分不一致, 然而实验结果的多元化与研究对象所处的阶元、边缘类型及实验设计等诸多因素皆有关[19]。通常在栖息地边缘会有不同的植被结构与微环境, 如植被类型、温度、光照、水分和土壤等生物与非生物因子[20], 这些因子将直接影响生物的取食行为与分布格局, 可能导致边缘存在高的生物多样性。当然任何边缘都处于动态变化之中, 这样的实验结果不具有普遍代表性, 只能作为该调查区特定的生态系统管理的理论指导。因此, 对边缘效应的动态演替进行长期监测与预报, 对边缘效应与生物多样性的内在关系进行本质上的探索, 是我们应该努力的方向。
调查结果显示, 3种边缘类型中, 只在阔叶林—针叶林边缘鞘翅目昆虫群落的多样性指数与植被特征有显著的回归关系。植被密度影响鞘翅目昆虫群落的 Shannon-Wiener多样性和丰富度, 植被密度和盖度影响鞘翅目昆虫的个体数量分布。由此看出, 植被特征对鞘翅目昆虫群落多样性的影响存在差异, 不仅表现在不同边缘类型上, 同一边缘类型不同位置植被特征的影响也不同, 这可能与鞘翅目昆虫不同类群对特定环境变量的选择和敏感程度有关[21]。

表4 植被特征与鞘翅目昆虫科多样性、丰富度、均匀度、优势度及个体数量间的回归分析Tab.4 Regression analysis of vegetation characteristics and family diversity, richness, evenness, dominance and abundance of Coleoptera insects
[1]杨贵军, 王新谱, 贾彦霞, 等.人工柠条—荒漠草地交错带拟步甲昆虫群落多样性[J].生态学报, 2016, 36(3):608–619.
[2]FLOHRE A, FISCHER C, AAVIK T, et al.Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids and birds[J].Ecological Applications, 2011, 21(5): 1772–1781.
[3]鲁佳雄, 朱丽得孜·艾山, 伊 龙, 等.阿尔金山自然保护区鞘翅目昆虫区系调查分析[J].干旱区研究, 2015, 32(6):1207–1212.
[4]王巍巍, 贺达汉, 张大治.荒漠景观地表甲虫群落边缘效应研究[J].应用昆虫学报, 2013, 50(5): 1383–1391.
[5]娄巧哲, 徐养诚, 马吉宏, 等.古尔班通古特沙漠南缘地表甲虫物种多样性及其与环境的关系[J].生物多样性,2011, 19 (4): 441–452.
[6]刘立品.子午岭木本植物志[M].兰州: 兰州大学出版社,1998: 4–13.
[7]党坤良, 宋小民.陕西子午岭自然保护区综合科学考察[M].杨凌: 西北农林科技大学出版社, 2004: 1–185.
[8]郑乐怡, 归鸿.昆虫分类[M].南京: 南京师范大学出版社, 2010: 564–652.
[9]张巍巍, 李元胜.中国昆虫生态大图鉴[M].重庆: 重庆大学出版社, 2011: 239–378.
[10]乔治·C.麦加文.昆虫[M].北京: 中国友谊出版公司,2007: 109–131.
[11]杨星科.中国动物志·昆虫纲(第 61卷): 鞘翅目·叶甲科·叶甲亚科[M].北京: 科学出版社, 2014: 1–641.
[12]MEINERS S J, PICKETT S T A.Changes in community and population responses across a forest-field gradient[J].Ecography, 1999, 22(3): 261–267.
[13]HELIOLA J, KOIVULA M, NIEMELA J.Distribution of carabid beetles (Coleoptera, Carabidae) across a boreal forest–clearcut ecotone[J].Conservation Biology, 2001,15(2): 370–377.
[14]赵志模, 郭依泉.群落生态学原理与方法[M].重庆: 文献和科学技术出版社, 1990: 147–279.
[15]马克平, 刘玉明.生物群落多样性的测度方法: Ⅰα多样性的测度方法(下)[J].生物多样性, 1994, 2(4): 231–239.
[16]王伯荪, 彭少麟.鼎湖山森林群落分析——Ⅹ.边缘效应[J].中山大学学报自然科学版, 1986, (4): 52–56.
[17]刘云慧, 宇振荣, 王长柳, 等.坝上地区农田和恢复生境地表甲虫多样性[J].生态学报, 2011, 31(2): 0465–0473.
[18]DAVIS C A, UTRUP J S.Response of terrestrial invertebrates to high-and low-diversity grassland restorations in south-central Nebraska[J].Restoration Ecology, 2010,18(S2): 479–488.
[19]廉振民, 于广志.边缘效应与生物多样性[J].生物多样性, 2000, 8(1): 120–125.
[20]杨贵军, 贾彦霞, 王新谱.苜蓿—荒漠草地交错带步甲昆虫多样性[J].环境昆虫学报, 2015, 37(3): 483–491.
[21]HUMPHREY J W, HAWES C, PEACE A J, et al.Relationships between insect diversity and habitat characteristics in plantation forests[J].Forest Ecology & Management,1999, 113(1): 11–21.
Edge effects of Coleoptera insect communities in Ziwuling National Nature Reserve, Shaanxi
XU Shicai*, WANG Lijuan
College of Life Sciences,Yan’an University,Yan’an Shaanxi716000,China
Three edge zones, broad-leaved-coniferous forest, farmland-shrub and grassland-shrub of Ziwuling National Nature Reserve in Shaanxi Province were selected as the sample plots, and the impact of the edge effect on the diversity of Coleopteran insect community was discussed at family level.A total of 4935 Coleopteran insects were collected, belonging to 28 families, among which the individual amount of Carabidae, Chrysomelidae and Coccinellidae accounted for 44.19%,21.95% and 13.66% of the total number of individuals respectively, which constituted the dominant group of Coleopteran insects in this area.The family diversity, richness and evenness of Coleopteran insects in the three edge types decreased from the edge to the adjacent habitats, with theEH´values greater than 1, showing a positive edge effect.The multiple regression analysis showed that the vegetation density affected the diversity and richness of Coleopteran insect community in the broad-leaved-coniferous forest, and the vegetation density and coverage affected the individual population distribution of the entire Coleopteran insects community.The above conclusions provide scientific evidence for the conservation and rational utilization of the biodiversity of Coleopteran insects in Shaanxi Ziwuling National Nature Reserve.
Coleoptera; community diversity; edge effect
10.14108/j.cnki.1008-8873.2017.05.026
Q968.1
A
1008-8873(2017)05-204-07
徐世才, 王丽娟.陕西子午岭国家级自然保护区鞘翅目昆虫群落的边缘效应[J].生态科学, 2017, 36(5): 204-210.
XU Shicai, WANG Lijuan.Edge effects of Coleoptera insect communities in Ziwuling National Nature Reserve, Shaanxi[J].Ecological Science, 2017, 36(5): 204-210.
2017-04-14;
2017-06-12
陕西省高水平大学建设专项资金项目“生态学”(2012SXTS03); 延安大学引导项目“陕北烟区烟粉虱的发生规律及其有机防控”(YD2015-11)
徐世才(1973—), 男, 陕西延安人, 硕士, 副教授, 硕士生导师, 主要从事昆虫生态与综合防治研究, E-mail: shicaixu@163.com
*通信作者:徐世才
猜你喜欢
水土保持学报(2022年5期)2022-10-10
西北农林科技大学学报(自然科学版)(2022年3期)2022-04-01
南方农业·下旬(2021年3期)2021-07-28
农家科技中旬版(2020年4期)2020-05-25
草业科学(2019年5期)2019-06-04
现代农业科技(2018年14期)2018-10-31
中国测试(2018年5期)2018-05-14
湖北农业科学(2016年21期)2017-03-18
鄱阳湖学刊(2017年1期)2017-02-16
天津师范大学学报(自然科学版)(2016年4期)2016-12-14
