
山西陵川南方红豆杉群落优势种生态位特征
2017-11-10 03:11刘泽宇
生态科学 2017年5期
王 霞, 铁 军 , 刘泽宇
1.山西师范大学生命科学学院, 山西临汾 041004 2.长治学院生物科学与技术系, 山西长治 046011 3.太行山生态与环境研究所, 山西长治 046011
山西陵川南方红豆杉群落优势种生态位特征
王 霞1, 铁 军2,3*, 刘泽宇1
1.山西师范大学生命科学学院, 山西临汾 041004 2.长治学院生物科学与技术系, 山西长治 046011 3.太行山生态与环境研究所, 山西长治 046011
为合理开发和保护山西濒危植物南方红豆杉资源提供依据, 采用Levins生态位宽度、Pianka生态位重叠和生态位相似性指数计算法, 分析了山西陵川南方红豆杉群落25个优势种生态位特征。结果表明: (1)山西陵川南方红豆杉群落25个优势种中, 生态位宽度值相对较大的为鹅耳枥(1.464)、苔草(1.450)和南方红豆杉(1.400), 而淫羊藿(0.049)、陕西荚蒾(0.862)和臭梧桐(0.936)的生态位宽度值相对较小; (2)25个优势种的生态位普遍产生重叠, 重叠程度在0.4以上的种对有 186对, 占总数的 62%, 表明种间竞争激烈; (3)生态位相似性比例在 0.4以上的种对有 156对, 占总数的52%, 表明优势种对资源的利用相似程度较大。(4)生态位宽度大的群落优势种, 其具有较强的竞争力, 对环境以及资源的利用能力较强, 与其它物种间的生态位重叠值较大, 生态位相似性比例也较高。
南方红豆杉; 优势种; 生态位宽度; 生态位重叠; 生态位相似性
1 前言
生态位(niche)是生态学中的一个重要概念, 用于描述某一物种在群落或生境中的地位和作用, 即一个种的生态位由它生存必需的全部环境因素组成, 能反映物种对环境资源的需求[1-2]。生态位相同的物种可能因竞争共同的资源而发生竞争排斥, 不能稳定共存[3]。生态位特征在揭示群落结构和功能、群落演替、群落的物种关系、物种多样性、种群进化、濒危物种评价和森林资源保护与利用等方面有重要作用, 对认识自然群落中物种共存和竞争机制具有重大意义[4]。
南方红豆杉(Taxus chinensisvar.mairei)又称美丽红豆杉, 隶属红豆杉科(Taxaceae)红豆杉属(Taxus),为集观赏、材用、药用于一身的珍贵物种, 是国家一级珍稀濒危保护植物, 其植物体内含有的紫杉醇是一种较好的抗癌药物[5]。在自然状态下, 南方红豆杉野生种群较少、再生能力差、繁殖力低、生长缓慢, 故野生资源非常少; 长期以来, 紫杉醇深受医药界、商业界的高度重视, 一些不法分子为了谋取利益对南方红豆杉乱采滥伐, 使其资源受到严重威胁, 现存物种的数量越来越少, 目前已处于濒危状态[6]。本文通过对山西陵川南方红豆杉群落优势种群的生态位进行研究, 分析南方红豆杉群落各层主要优势种群的生态位关系, 揭示群落内各种群对资源的利用情况, 从而为山西陵川红豆杉自然保护区南方红豆杉天然林的经营管理、开发利用、森林资源以及濒危物种的保护提供理论依据。
2 研究地区与研究方法
2.1 研究地概况
陵川红豆杉自然保护区位于山西省晋城市陵川县东南部的夺火乡、马圪当乡和古郊乡境内(35°34.796′N—35°49.228′N, 113°21.401′E—113°29.401′E), 于 2002年6月被山西省政府批准为省级自然保护区。该保护区离县城30—70 km。该地区山体陡峭, 气候温和,水源充足, 森林茂盛, 海拔628—1469 m之间, 相对高差约841 m, 属暖温带半湿润大陆性季风气候, 年平均气温5—11 ℃, 年最高温度34.5 ℃, 极端最低温度–23.7 ℃, 1 月份均温–7—3.8 ℃, 7 月份均温0.7—24.0 ℃, ≥10 ℃的积温2753—3671 ℃, 无霜期165—200 d, 年降水量503—673 mm。地层结构较为复杂,主要由石灰岩和砂石岩、结晶岩和变质岩及煤系地层组成。地带土类为褐土, 土壤类型有山地褐土、山地淋溶褐土和山地棕壤等[7]。
据报道, 陵川红豆杉自然保护区内具有较丰富的植物种类, 共有维管植物94科, 325属, 525种。以北温带典型科属为主, 就种数而言菊科(Compositae)和禾本科(Gramineae)占优势, 其次是豆科(Leguminosae)、蔷薇科(Rosaeeae)、唇形科(Labiatae)、毛茛科(Ranuneulaceae)、百合科(Liliaceae)和十字花科(Crueiferae)等。该保护区内野生动物资源也比较丰富, 主要有金钱豹、猕猴、蛇类、雕等200余种。区内整体生态环境良好[8-9]。
2.2 研究方法
2015年6—8月和2016年4—5月, 在山西陵川红豆杉自然保护区, 海拔 850—1200 m 区域内, 依据海拔、坡度、坡向等条件选取了30个10 m×10 m的南方红豆杉群落样方进行植被调查; 采用对角线法在每个大样方内设置灌木样方(5 m×5 m)和草本样方(1 m×1 m)各2个; 乔木层样方30个、灌木层和草本层样方各60个。在调查样方内, 记录乔木高度、枝下高、胸径、冠幅和盖度等, 灌木和草本植物的丛数、盖度和高度等; 用GPS测量海拔和经纬度, 同时记录坡度、坡向、坡位、枯枝落叶层厚度、土壤类型及人为干扰状况等环境因子[10]。对于样方内不确认的种类进行科的初步断定, 并采集植物标本、拍照, 带回实验室查阅资料确认。
2.3 数据统计与分析
2.3.1 重要值
重要值(important value)是以综合数值来表示群落中不同植物的相对重要性。由于样方内的灌木层和草本层植物种类较多, 共记录了100多个物种, 依据各物种的重要值大小, 并综合其在样方中的盖度、高度及出现的频率(剔除频度<10%), 得到 25个优势种。乔木、灌木、草本植物的重要值计算依据白玉芳等[11]和陈龙涛等[12]的计算方法, 其计算公式为:

式中: 相对多度为样方内某一物种的个体数占全部物种个体数的百分比; 相对高度为样方内某一物种的高度和占全部物种高度之和的百分比; 相对优势度为样方内某一树种的胸高断面积之和占所有物种胸高断面积之和的百分比; 相对盖度为样方内某一物种的分盖度占所有分盖度之和的百分比。
2.3.2 生态位宽度
生态位宽度(niche breadth)是指某一物种所利用的各种不同资源的总和, 主要反映物种对资源的利用程度。生态位宽度采用Shannon-Wiener指数进行计算[1,13]。其计算公式如下:

式中:B(sw)i为物种i的生态位宽度,Pij是种i在第j个资源的利用占其对全部资源利用的频度, 即,nij为物种i在资源j上的优势度(文中即物种在样方中的重要值),r为资源等级数,上述方程具有值域[0, logr]。B(sw)i值越大, 说明该种对环境的适应能力越强, 对资源利用较为充分, 而且在群落中具有较强的竞争力, 处于优势地位[14-16]。
2.3.3 生态位重叠
生态位重叠(niche overlap)是指一定资源序列上,两个物种利用同等级资源而相互重叠的情况。生态位重叠采用Pianka指数进行计算, 其计算公式为:

式中:NO为生态位重叠值,nij和nkj为种i和种k在资源位j上的优势度(文中为物种重要值)[17-18]。
2.3.4 生态位相似性比例
生态位相似比例(niche similarity)是指两个物种利用资源的相似程度, 其计算公式为:

式中:Cih表示物种i与物种h之间的相似程度, 且有Cih=Chi, 具有域值[0, 1];Pij、Phj分别为物种i和物种h在资源位j上的重要值百分率[14,19]。
3 结果与分析
3.1 南方红豆杉群落主要优势种的重要值

表1 山西陵川南方红豆杉群落25个优势种重要值Tab.1 25 dominant species importance value of Taxus chinensis var.mairei community of Lingchuan in Shanxi
从表1可看出, 在山西陵川南方红豆杉群落25个优势种中, 乔木层生态位宽度值较大的为鹅耳枥(Carpinus turczaninowii)(53.165)和南方红豆杉(39.085); 灌木层重要值最大的是连翘(Forsythia suspensa)(10.564), 葱皮忍冬(Lonicera ferdinandii)(5.782)、灰栒子(Cotoneaster acutifolius)(5.718)和陕西荚蒾(Viburnum schensianum)(5.304)为次之; 草本层中苔草(Carexspp.)(28.940)的重要值最大, 淫羊藿(Epimedium brevicornu)(10.320)、球果堇菜(Viola collina)(6.534)和荩草(Arthraxon hispidus)(5.971)为次之; 重要值较小的为灌木层的土庄绣线菊(Spiraea pubescens)(2.575), 草本层的叶底珠(Flueggea suffruticosa)(2.390)和狭叶紫竹(Callicarpa jaonicavar.angu)(2.382), 说明它们在样方中出现的频度相对较低, 且分布较为分散。由此表明, 鹅耳枥和南方红豆杉在山西陵川南方红豆杉群落中占有重要地位,是该群落中的建群种。
3.2 南方红豆杉群落主要优势种生态位宽度
生态位宽度是反映物种对环境资源利用状况的尺度, 其大小取决于物种的生态适应性、种间竞争力和分布范围。由表2可看出, 陵川南方红豆杉群落25个优势种中, 乔木层生态位宽度值最大的为鹅耳枥(1.464); 灌木层生态位宽度值最大的为鞘柄菝葜(Smilax stans)(1.354), 其次为陕西荚蒾(1.317)、葱皮忍冬(1.292)和美丽胡枝子(Lespedza formosa)(1.275); 草本层生态位宽度值较大的为苔草(1.450),荩草(1.346)、一叶兰(Aspidistra elatior)(1.329)次之。生态位宽度较高的鹅耳枥、苔草、南方红豆杉和鞘柄菝葜为该群落的建群种, 在群落中的地位较高,对环境的适应能力以及对资源的利用能力都较强,而且分布较为广泛, 在多数样方中均出现, 对群落的组成和结构起着决定性作用, 故其生态位宽度较大; 生态位宽度较小的树种为草本层的叶底珠(0.940)和狭叶紫竹(0.862), 淫羊藿(0.049)的生态位宽度最小, 由于生态位宽度较小的物种对资源的利用能力和环境的适应能力都较弱, 在林内多数以散生的形式存在, 分布范围较为狭窄, 故其生态位宽度相应也较小。
3.3 南方红豆杉群落主要优势种生态位重叠

表2 山西陵川南方红豆杉群落25个优势种生态位宽度Tab.2 25 dominant species niche breadth of Taxus chinensis var.mairei community of Lingchuan in Shanxi
从表3可知, 南方红豆杉群落中25个优势种构成300个种对, 生态位重叠值范围介于0.021—0.928之间。生态位重叠值在0.9以上的只有一对, 为鹅耳枥与苔草(0.928); 在0.8—0.9之间的有6对, 分别是鹅耳枥与连翘(0.853)和荩草(0.815)、南方红豆杉与土庄绣线菊(0.821)、苔草与连翘(0.827)、荩草(0.818)和粘须菝葜(Smilax stans)(0.803)。鹅耳枥和苔草的生态位宽度较大, 说明其对环境适应能力以及对资源利用能力都比较强, 所以与其它物种的生态位重叠也较大, 重叠的物种不同, 生态位重叠值也存在一定的差异。生态位重叠值在0.7—0.8之间的有31对,如苔草与葱皮忍冬(0.733)、连翘与淫羊藿(0.753)、灰栒子与美丽胡枝子(0.716)等; 生态位重叠值在0.6—0.7之间的有 39个种对, 如粘须菝葜与荩草(0.610)、一叶兰与黄精(Polygonatum sibiricum)(0.636)、玉竹(Polygonatum odoratum)与鞘柄菝葜(0.634); 生态位重叠值在 0.5—0.6之间的有 51对,如美丽胡枝子与唐松草(Thalictrum aquilegiifoliumvar.sibiricum)(0.527)、鸡腿堇菜(Viola acuminata)与土庄绣线菊(0.597)等; 生态位重叠值在 0.4—0.5、0.3—0.4、0.1—0.3之间的分别有58、57、56对; 生态位重叠值在 0.1以下的只有一对, 为土庄绣线菊与狭叶紫竹(0.021), 说明它们在群落中数量较少, 在样方中出现的频度相对较低, 对资源利用能力较弱,对生境的需求不同, 生态适应范围较窄, 生物学和生态学特性存在差异, 故它们之间几乎不存在生态位重叠; 生态位重叠值在0.4—0.8之间的有179对,占总数的 59.67%, 表明优势种群间一半以上的植物种群在资源利用上存在相似性。

?
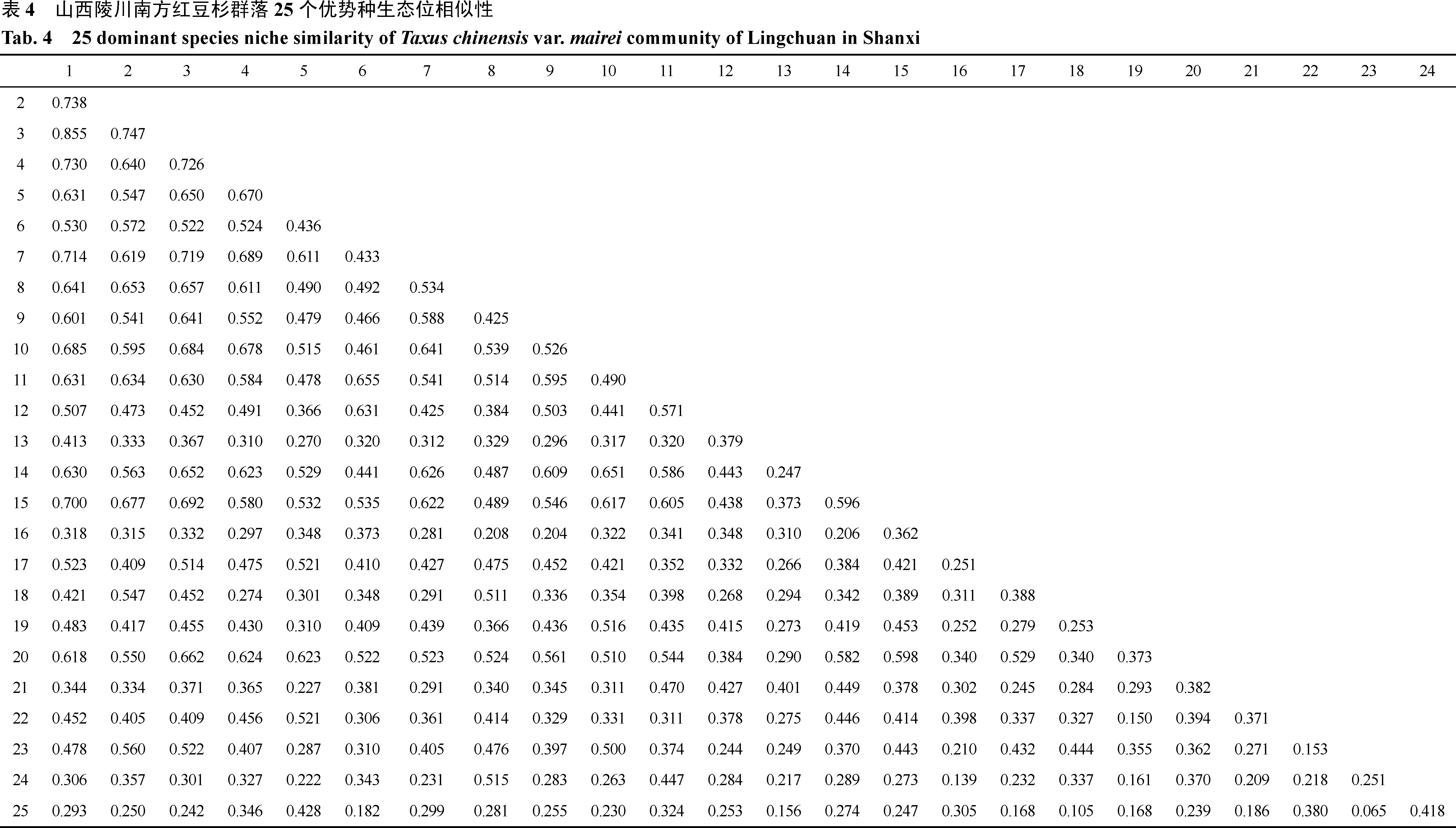
3.4 南方红豆杉群落主要优势种生态位相似性
从表4可看出, 陵川南方红豆杉群落25个优势种300个种对间的生态位相似性比例范围介于 0.065—0.855之间。生态位相似性比例最大的是鹅耳枥与苔草(0.855), 说明它们的生态需求较为一致; 生态位相似性比例在0.6—0.8之间的有44个种对, 如鹅耳枥与南方红豆杉(0.738)、苔草与连翘(0.726)、球果堇菜与一叶兰(0.655); 生态位相似性比例在 0.4—0.6之间的有112个种对, 如连翘与灰栒子(0.552)、南方红豆杉与淫羊藿(0.547)、陕西荚蒾与溲疏(Deutzia scabra)(0.441); 生态位相似性比例在 0.2—0.4之间的有132对, 生态位相似性比例在0.1—0.2的有10对, 如狭叶紫竹与球果堇菜(0.182)、糙苏(0.156)、二花六道木(Abelia biflora)(0.168)、鸡腿堇菜(0.105)、玉竹(0.168)和黄精(0.186), 叶底珠与臭梧桐(Clerodendrum trichotomum)(0.139)、玉竹(0.161), 唐松草与玉竹(0.150)、土庄绣线菊(0.153), 这10个种对生态位相似性比例都较低, 说明它们对资源的利用状况存在差异; 土庄绣线菊与狭叶紫竹的生态位相似比例最小(0.065), 说明它们的生物学特性和生态学特性差异较大, 对生境的需求以及对资源的利用有明显的差异。
一般来说, 生态位宽度较高的物种, 它们的生态位相似性比例也较大, 反之亦然。如生态位宽度大的鹅耳枥与南方红豆杉(0.738)、南方红豆杉与苔草(0.747)、苔草与荩草(0.719)等种对之间皆产生较大的生态位相似性。生态位宽度较小的糙苏(Phlomis umbrosa)与狭叶紫竹(0.156)、叶底珠与臭梧桐(0.139)等物种对之间的生态位相似性比例也较小。由此看来, 生态位宽度较大的物种间生态位相似性也高,原因可能是生态位宽度较大, 所占据的资源位也较多, 从而导致对资源的利用较为相似。但有时生态位宽度低的物种对会比生态位宽度高的物种对相似性比例要大, 例如连翘与淫羊藿和葱皮忍冬与溲疏,前一种对的生态位宽度较后者小, 但前者的生态位相似性比例要比后者高, 也就是说两者有可能同时高, 有可能一高一低。生态位重叠与生态位相似性比例, 它们的变化趋势是一致的, 生态位重叠值越大, 生态位相似性比例越大; 反之则小。
4 结论与讨论
4.1 南方红豆杉群落主要优势种生态位宽度分析
生态位宽度反映了物种对环境的适应状况和对资源的利用程度, 体现了物种的生态适应性以及在生境中的分布状况, 衡量它对资源多样性的利用水平。通常, 生态位宽度的大小取决于物种对环境的适应能力和对资源的利用能力, 生态位宽度会随环境的变化而变化, 环境的改变会引起物种对资源的利用和对环境适应性的变化[20]。一般来说, 物种的生态位宽度越大, 表明其分布较为广泛, 对资源的利用能力较强, 环境的适应能力较强, 同时也表明该物种倾向于泛化种, 在资源竞争中处于优势[21-22]。在山西陵川南方红豆杉群落中, 鹅耳枥、南方红豆杉、苔草、鞘柄菝葜、荩草、一叶兰和陕西荚蒾等物种的生态位宽度值也较大, 对资源利用较为充分, 适应多样化的生境, 分布范围较广且分布均匀。
4.2 南方红豆杉群落主要优势种生态位重叠分析
当两个物种利用同一资源或共同占有某一资源(营养成分和空间等)时, 就会出现生态位重叠现象,生态位重叠较大的物种要么有相近的资源利用方式,要么对生境因子有互补性的要求, 即生态位重叠是两个种在其与生态因子联系上的相似性[23-24]。生态位重叠更多的是用于研究种间竞争, 通常两个物种的生态位重叠值越大, 两个种之间存在的竞争越激烈。这与Levins(1968)的结论相悖, 他认为: 生态位重叠不一定就发生竞争, 竞争是在资源供应不足且生态位重叠条件下形成的, 如果资源很丰富, 生态位重叠不一定伴随有竞争排斥的现象发生。在陵川南方红豆杉群落中, 鹅耳枥与苔草(0.928)的生态位重叠值最大, 这可能是由二者的生态学特性相似,在研究样地中分布较广泛而导致的。值得一提的是,灌木层中的陕西荚蒾与草本层中的狭叶紫竹生态位重叠较低(0.188), 这可能由于不同的生态学特性以及占有不同的资源位导致的。鹅耳枥与南方红豆杉的生态位重叠值较大(0.719), 可能由于它们对空间的占据以及对资源(光照)的利用较为相似的结果。
?
研究表明, 生态位重叠与物种生态位宽度有一定关系, 生态位宽度大的物种与其它物种的生态位重叠一般也较高, 如鹅耳枥与南方红豆杉(0.719)、苔草与荩草(0.818)、陕西荚蒾与美丽胡枝子(0.724)等物种, 它们的生态位宽度都较大, 生态位重叠也较高, 说明这些物种的生态学特性和生物学特性相近,对环境具有相似的需求。但有些物种的生态位宽度较大, 而与其它物种的生态位重叠却较小, 如南方红豆杉与糙苏(0.306)、荩草与鸡腿堇菜(0.278)等物种, 这说明了生态位宽度和生态位重叠之间并不存在绝对的正相关关系, 这与柴宗政等[25]和刘晓宁等[26]的结论相吻合。这可能由于其对资源的利用不同以及不同的生物学特性导致。因此, 在一定范围之内生态位宽度值较大的物种之间不一定产生较大的生态位重叠[14]。
4.3 南方红豆杉群落主要优势种生态位相似性分析
生态位相似性比例能够反映两个物种对资源利用的相似程度, 生态位宽度与生态位相似性比例有一定关系[14]。一般来说, 生态位宽度值大的种群与其它种群间的生态位相似性比例也较大,例如鹅耳枥与苔草(0.928)、苔草与荩草(0.818)、南方红豆杉与葱皮忍冬(0.769)等; 生态位宽度值小的种群与其它种群间的生态位相似性比例也较小,例如狭叶紫竹与黄精(0.186)、叶底珠与玉竹(0.161)、唐松草与土庄绣线菊(0.153)等。生态位重叠与生态位相似性比例也存在一定的关系, 生态位重叠较大的种对生态位相似性比例也大, 例如鹅耳枥与南方红豆杉(生态位重叠0.719、生态位相似性比例0.738)、南方红豆杉与苔草(同上, 0.772、0.747)、连翘与淫羊藿(0.753, 0.670); 生态位重叠小的种对生态位相似性比例也小, 例如叶底珠与玉竹(0.144, 0.161)、狭叶紫竹与土庄绣线菊(0.021,0.065)。说明生态位宽度、生态位重叠与生态位相似性比例之间存在正相关关系, 这与哀建国等[27]在石垟林场省级森林公园常绿阔叶林种群生态位特征一文中所得结果一致。此外, 生态位相似性比例的高低与种的生活习性有着密切的关系, 相同生活习性的种具有相同的资源要求, 它们之间的生态位相似比例也高, 反之则较低[28]。如鞘柄菝葜与一叶兰(0.605), 同属于百合科(Liliaceae)的物种, 它们对资源的利用相似性程度就高, 故它们的生态位相似比例值也较大; 如叶底珠与臭梧桐(0.139), 它们分别属于大戟科(Euphorbiaceae)和马鞭草科(Verbenaceae)的植物, 它们的生活习性存在差异; 因此, 它们对资源的利用存在差异, 故生态位相似性比例值也相对偏低。
4.4 南方红豆杉濒危原因以及保护对策的分析
南方红豆杉缺乏较强的竞争力, 竞争力较弱是导致物种濒危的重要因素之一。对光和空间的争夺是南方红豆杉与其他植物竞争的主要表现形式[6]。南方红豆杉为典型的阴性树种, 在郁闭度 0.5—0.6时其幼树和成树长势较好。随着林冠郁闭度的增加,长势会逐渐减弱, 这对于南方红豆杉的生长极其不利。因此在郁闭度较大的森林群落中, 适当的人为降低林冠郁闭度会提高南方红豆杉幼树的成活率,对于南方红豆杉幼树成活率低将会有所改善。在群落生境的构建中, 南方红豆杉适应性较弱, 加之其生长极其缓慢, 在对空间的利用上不占优势, 一旦竞争力较强的物种种群数量增加时, 剧烈的种间竞争, 将导致南方红豆杉种群数量的下降。鉴于此, 可以适当减少与南方红豆杉有较强竞争力的物种, 从而降低种间竞争力, 使南方红豆杉在空间以及资源利用上处于优势地位, 使其更好的生长。通过对南方红豆杉群落的实地考察, 我们发现南方红豆杉砍伐现象较为严重, 因此建议保护区加强对南方红豆杉资源的管理和保护, 积极做好宣传教育工作, 使当地群众以及管理人员充分认识南方红豆杉资源保护的重要性, 自觉爱护这一宝贵资源。
[1]毕润成.生态学[M].北京: 科学出版社, 2012.
[2]张忠华, 梁士楚, 胡刚.桂林岩溶石山阴香群落主要种群生态位研究[J].林业科学研究, 2009, 22(1): 63–68.
[3]VANDERMEER J H.Niche theory[J].Annual Review of Ecology and Systematics, 1972, 3: 107–132.
[4]陈旭, 卜兆君, 王升忠, 等.长白山哈泥泥炭地七种苔藓植物生态位[J].应用生态学报, 2009, 20(3): 574–578.
[5]茹文明, 张金屯, 张桂萍, 等.濒危植物南方红豆杉的研究进展[J].长治学院学报, 2005, 22(5): 16–20.
[6]茹文明, 张金屯, 张峰, 等.濒危植物南方红豆杉濒危原因分析[J].植物研究, 2006, 26(5): 624–628.
[7]华振铃, 刘芳, 王志雄, 等.山西陵川的南方红豆杉(Taxus chinensisvar.mairei)年轮特征的初步研究[J].首都师范大学学报(自然科学版), 2010, 31(6): 44–49.
[8]张桂萍.陵川山区野生植物资源研究[J].河南科学,1996, 14(增刊): 104–106.
[9]李岩.陵川县磨河水库工程建设对山西南方红豆杉自然保护区的影响[J].山西水利科技, 2015, 45(2): 123–125.
[10]李燕芬, 铁军, 张桂萍, 等.山西蟒河国家级自然保护区人工油松林生态位特征[J].生态学杂志, 2014, 33(11):2905–2912.
[11]白玉芳, 毕润成, 白玉宏, 等.山西太岳山稀有濒危植物脱皮榆群落种间关联[J].广西植物, 2014, 34(1): 56–61.
[12]陈龙涛, 高润梅, 石晓东.山西蟒河自然保护区南方红豆杉群落生态位研究[J].林业资源管理, 2016, 45(2):68–73.
[13]茹文明, 铁军.南方红豆杉生态学研究[M].北京: 中国林业出版社, 2012.
[14]铁军, 张晶, 彭林鹏, 等.神农架川金丝猴栖息地优势树种生态位及食源植物[J].植物生态学报, 2009, 33(3):482–491.
[15]毛空, 张殷波, 张峰.关帝山森林植被优势种群生态位[J].生态学杂志, 2013, 32(11): 2920–2925.
[16]LEVINS R.Evolution in Changing environments: Some theoretical explorations[M]. Princeton: Princeton University Press, 1968: 158–160.
[17]谢春平.Microsoft Excel 2003在群落生态位参数计算中的应用[J].安徽农业科学, 2008, 36(11): 4791–4792.
[18]PIANKA, E R.The Structure of Lizard Communities[J].Annual Review of Ecology and Systematics, 1973, 4:53–74.
[19]刘金福, 洪伟.格氏栲群落生态学研究——格氏栲林主要种群生态位的研究[J].生态学报, 1999, 19(3):347–352.
[20]崔丽娟, 李伟, 赵欣胜, 等.采砂迹地型湿地恢复过程中优势种群生态位研究[J].生态科学, 2013, 32(1): 73–77.
[21]刘巍, 曹伟.长白山云冷杉群落主要种群生态位特征[J].生态学杂志, 2011, 30(8): 1766–1774.
[22]李箐, 骆有庆, 石娟, 等.阿尔山地区兴安落叶松林下植物种群生态位[J].林业科学研究, 2011, 24(5): 651–658.
[23]史作民, 程瑞梅, 刘世荣.宝天曼落叶阔叶林种群生态位特征[J].应用生态学报, 1999, 10(3): 265–269.
[24]叶铎, 温远光, 邓荣艳, 等.大明山常绿阔叶林演替序列种群生态位动态特征[J].生态学杂志, 2009, 28(3):417–423.
[25]柴宗政, 王得祥, 张丽楠, 等.秦岭山地天然油松群落主要植物种群生态位特征[J].生态学杂志, 2012, 31(8):1917–1923.
[26]刘晓宁, 张钦弟, 闫明, 等.五鹿山国家级自然保护区油松群落优势种生态位研究[J].植物研究, 2012, 32(6):712–716.
[27]哀建国, 吴谷汉, 陈锦宇, 等.石垟林场省级森林公园常绿阔叶林种群生态位特征[J].浙江大学学报(农业与生命科学版), 2006, 32(5): 576–584.
[28]王念奎, 李海燕, 荣俊冬, 等.突脉青冈群落乔木层优势种群生态位研究[J].福建林学院学报, 2010, 30(2):128–132.
Niche characteristics of dominant plant species in Taxus chinensis var.mairei community of Lingchuan, in Shanxi
WANG Xia1, TIE Jun2,3*, LIU Zeyu1
1.College of Life Science,Shanxi Normal University,Linfen041004,Shanxi,China2.Department of Biological Sciences and Technology,Changzhi College,Changzhi046011,Shanxi,China3.Ecological and Environmental Research Institute of Taihang Mountain,Changzhi046011,Shanxi,China
To provide data for the effective utilization and protection of endangered plantTaxus chinensisvar.maireiresources in Shanxi, we analyzed the niche characteristics of 25 dominant species ofT.var.maireicommunity of the Lingchuan, in Shanxi by using the formulas of Levins niche breadth, Pianka niche overlap and the niche similarity.The results showed are as follows.(1) The 25 dominant species ofT.var.maireicommunity of the Lingchuan in Shanxi, the niche breadth ofCarpinus turczaninowii(1.464),Carexspp.(1.450) andT.var.mairei(1.400) were much greater; those ofEpimedium brevicornu(0.049),Viburnum schensianum(0.862) andClerodendrum trichotomum(0.862) were much smaller.(2) The niche overlap among the 25 dominant species occurred generally, and the species (186) had overlap degree being higher than 0.4, accounting for 62% of total pairs, suggesting the strong competition between species.(3) The species with niche similarity proportion above 0.4 reached to 156, accounting for 52 % in total, indicating that dominant species had a higher similarity proportion in resource utilization.(4) The dominant species in the community with larger niche breadth had stronger competitiveness, and the environment and resource utilization ability was stronger, and had greater niche overlap and niche similarity proportion with other populations.
Taxus chinensisvar.mairei; dominant species; niche breadth; niche overlap; niche similarity
10.14108/j.cnki.1008-8873.2017.05.009
Q948.1
A
1008-8873(2017)05-064-09
王 霞, 铁 军, 刘泽宇.山西陵川南方红豆杉群落优势种生态位特征[J].生态科学, 2017, 36(5): 64-72.
WANG Xia, TIE Jun, LIU Zeyu.Niche characteristics of dominant plant species inTaxus chinensisvar.maireicommunity of Lingchuan, in Shanxi[J].Ecological Science, 2017, 36(5): 64-72.
2016-07-21;
2016-09-10
山西省自然科学基金项目(2012011034-6; 2010011041-2); 山西省重点扶持学科项目和长治学院博士启动基金项目
王霞(1990—), 女, 山西省朔州市人, 硕士研究生, 生态学专业, 主要从事植物生态学和保护生物学研究, E-mail: 572738849@qq.com
*通信作者:铁军(1974—), 男, 内蒙古通辽市人, 教授, 博士, 主要从事植物生态学和保护生物学研究, E-mail: tiejun74@163.com
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
草地学报(2021年6期)2021-07-05
林业调查规划(2020年3期)2020-06-03
西江月(2018年5期)2018-06-08
广西林业科学(2016年4期)2016-03-16
党员生活(2015年6期)2015-07-16
肝胆胰外科杂志(2015年4期)2015-02-27
草业科学(2012年5期)2012-03-13
