
沙棘雌雄株叶片解剖结构比较研究
2017-11-10 02:23:44郭学民
植物研究 2017年1期
李 娜 郭学民 孟 阳
(河北科技师范学院生命科技学院,昌黎 066600)
沙棘雌雄株叶片解剖结构比较研究
李 娜 郭学民*孟 阳
(河北科技师范学院生命科技学院,昌黎 066600)
为了研究沙棘雌、雄株叶片的第二性征,本文采用石蜡切片法观察了沙棘雌、雄株叶片结构的差异。结果表明:(1)沙棘雌、雄株叶片均由表皮、叶肉和叶脉3部分组成,表皮均由1层细胞构成,表皮毛发达,上表皮有拟泡状细胞;叶肉栅栏组织与海绵组织分化明显。(2)雌株上表皮具更多的拟泡状细胞,其主脉韧皮部薄壁细胞及其下方的一些薄壁细胞含较多的后含物,下表皮的表皮毛更浓密;而雄株的叶片厚度、叶片上表皮厚度、栅栏组织厚度、栅栏组织厚度/海绵组织厚度均显著大于雌株,且其主脉维管束更发达。结果表明,沙棘雌雄株叶片解剖结构存在明显差异,这些差异是第二性征的表现,也是沙棘长期进化中形成的稳健的适应策略,可能有利于该物种的繁衍。
沙棘;雌雄异株;叶片;比较解剖
高等动物普遍为雌雄异体,而高等植物则多为雌雄同株。雌雄异株植物是指雌花与雄花分别位于不同植株上的单性花植物种类。植物性多态的现象,反映的是不同植物在快速繁育和遗传选择间的平衡以及对环境的适应[1]。据Renner等[2]的研究表明,在自然界的被子植物中仅约7%的属和6%的种具有雌雄异株现象。尽管雌雄异株植物所占比例不多,但其物种资源丰富。据调查,其种类数占被子植物总种数的4%~10%。在热带森林中,雌雄异株植物占木本植物的比例可高达36%[3~4]。雌雄异株植物对稳定维持生态系统和保护物种多样性有着积极的作用,同时也是陆地生态系统的重要成员之一[5]。
人们对动物雌雄两性间在行为、生理和生态活动上存在的差异已做了大量研究[6],但对植物雌雄两性的形态、生理、生长及两性关系却知之甚少。首先意识到雌雄异株植物之间在繁殖和资源需求上具有性别差异的,是英国生物学家达尔文[1]。此后雌雄异株间的差异引起了国内外学者的广泛关注。早期人们已经开展了关于雌雄异株植物的进化、种群结构与动态、生长、空间分布等方面的研究[3]。近20年来,各国学者对雌雄异株植物的兴趣有增无减,有关雌雄异株植物的差异性研究呈现快速增长的势头。目前,国内外有关植物雌雄异株的研究,主要集中在雌雄性别的决定机制[7]、雌雄个体形态和生理差异[8~9]、雌雄个体对环境因子的响应差异[10]、繁殖代价与防御策略差异[11]等方面。
叶片是植物进行光合作用和代谢功能的主要器官,直接暴露在空气中,对生态条件的反应最为明显,其生长发育和形态特征直接影响到植物的基本行为和功能。尤其是,由于长期的适应性进化和雌雄性别的分株表达,雌雄异株植物叶片在结构上可能出现一定的性别差异。因此,可根据其叶片的形态结构特征、生长情况等,判断雌雄异株植物对环境的适应性以及生态对策的不同。另外,由于植物生理生化指标易受环境的改变而表现出不同的变化,而植物的形态和结构,是在长期特定生态条件下形成的结果,不会随环境因子的暂时改变而发生较大的变化,因而在植物诸多特性研究中,形态解剖结构特征可作为研究雌雄异株植物适应性一个重要的参考指标[12~13]。
沙棘(HippophaerhamnoidesL.)又名醋柳、酸刺、酸溜溜等,属胡颓子科(Elaeagnaceae)沙棘属(Hippophae),为雌雄异株的多年生落叶直立灌木或小乔木。我国是世界上少数拥有丰富天然沙棘种质资源的国家,沙棘的生命力顽强,抗风耐沙、耐干旱、耐盐碱瘠薄、耐酷暑、耐低温,具有广泛的适应性,在我国西北、华北、东北、西南各省(区)均有分布,因此是改善土壤肥力、促进生态平衡、防风固沙、护岸固堤的先锋树种。其果实富含有机酸、蛋白质、黄酮类化合物、脂溶性维生素等多种营养物质和生物活性物质,是天然的药用资源,具有药用价值的同时,还可作为食品、染料、香料、饲料等原料。
目前,沙棘已成为一种兼具经济价值、生物科学与社会效益的树种,各国对于沙棘的开发利用日益广泛,研究也越来越深入。迄今,在叶片解剖方面,已有人比较了沙棘不同品种[14~17]、不同种类[18]、同一性别不同品种[19]以及各地区沙棘与不同种植物[20]叶片的形态解剖结构和抗旱性,这些研究均未进行性别区分。Li等[21]研究了不同海拔沙棘叶片形态特性的性别差异,认为胁迫环境以各种方式对雌株的负面影响大于雌株。
本文以沙棘雌、雄株叶片为研究对象,对比探讨沙棘雌、雄株叶片解剖结构的差异,以期为探讨沙棘雌、雄株对环境适应性差异和生产实践的选育提供基础资料,丰富人们对植物第二性征的认识。
2 材料与方法
2.1 采样地概况
试材采自小五台自然保护区金河口保护区山基部,该区位于河北省境西部偏北,北纬39°50′~40°07′,东经114°47′,气候类型属暖带季风区,气温低、昼夜温差大,降水集中于夏季,年降水420~700 mm,无霜期短60~140 d,年均温2.5~6℃,冬季台顶夜间温度低过-40℃,土壤类型主要为褐土、棕色森林土及亚高山草甸土,阳坡土壤贫瘠,阴坡则较肥沃[22]。
2.2 方法
选取生长健壮、生长势一致、健康、同一方位的沙棘雌、雄株枝条中部新鲜叶片,取叶脉中段及叶中约0.5 cm2的组织,以FAA固定液(70%乙醇90 mL、38%甲醛5 mL、冰醋酸5 mL)固定,依次经过乙醇梯度脱水、透明、浸蜡并用石蜡包埋,制成连续石蜡切片,厚度为8 μm,番红—固绿双重染色,中性树胶封片。
在光学显微镜Olympus BX51下观察,以照相机DP72拍照,每个样品观察30个视野,并用PhotoshopCS 6.0软件测定叶片结构参数:叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度、主脉维管束木质部高度、主脉维管束高度,计算组织结构致密度和组织结构疏松度、主脉木质部面积占维管束百分比,所有数据以平均值±标准差表示。并作如下计算:
组织结构致密度=栅栏组织厚度/叶片厚度×100
(1)
组织结构疏松度=海绵组织厚度/叶片厚度×100
(2)
主脉木质部面积占维管束百分比=主脉木质部面积/主脉维管束面积×100
(3)
2.3 数据处理
利用软件DPS 7.05,对雌、雄株叶片厚度、上表皮厚度、下表皮厚度、栅栏组织厚度、栅栏组织占叶肉百分比、海绵组织厚度、栅栏组织厚度/海绵组织厚度、细胞结构致密度、细胞结构疏松度、主脉维管束高度、主脉维管束木质部高度、主脉木质部面积占维管束百分比等各项指标进行t-检验。
3 结果分析
3.1 雌雄株叶片解剖结构观察
雌、雄株叶片属于典型异面叶,其解剖结构均由表皮、叶肉、叶脉3部分组成,其形态解剖结构特征基本一致(图1:A~B)。
3.1.1 叶片表皮组织结构特征
由图1:G~H可看出,雌、雄株叶片上、下表皮均由1层无叶绿体的细胞构成,上表皮细胞近似矩形或矩圆形,多数细胞的细胞质浓稠;下表皮细胞略小于上表皮,为椭圆形或圆形,呈不规则起伏。个别上表皮细胞显著增大,胞腔透明,呈泡状,类似于禾本科或莎草科植物叶表皮上的泡状细胞,为拟泡状细胞。雌、雄株叶片上、下表皮均具表皮毛,其中上表皮毛星散分布,下表皮毛层层紧密遮盖。
3.1.2 叶肉组织结构特征
雌、雄株叶肉组织均有明显的栅栏组织与海绵组织的分化(图1:E~F)。栅栏组织发达,位于叶近轴面表皮下面,排列紧密,细胞长短不一,多为2层,偶有3层。海绵组织细胞疏松、不规则,分布在叶远轴面的下表皮上方,数量少、不发达,有不规则的胞间隙相连,其中有叶脉穿过。
3.1.3 主脉维管束结构特征
由图1:C~D可看出,雌、雄株叶片主脉维管束单一,均由木质部、少量形成层和韧皮部构成,其中木质部导管呈径向排列,倒伞状分布于韧皮部上方,构成维管束的主要部分。束内形成层活性较强。韧皮部位于木质部下方,细胞较小,其中部分薄壁细胞及其下方的一些薄壁组织细胞中可见许多呈红褐色颗粒状后含物。
3.2 雌雄株叶片结构的比较分析
沙棘雌雄株叶片解剖结构组成一致,但各类型组织在细胞组成、数量、大小、厚度方面存在差异。
3.2.1 雌雄株叶表皮结构比较分析
与雄株相比,雌株上表皮分散有较多的大型的拟泡状细胞,这些细胞的细胞质较稀薄,另外,雌株下表皮细胞后含物丰富,其表皮毛更浓密(图1:E~G)。雄株上表皮厚度比雌株大10.2%(P<0.05),而雌雄株下表皮厚度差异不显著(P>0.05)(表1)。
表1沙棘雌、雄株叶片解剖结构特征
Table1LeafanatomicalstructurecharacteristicsinfemaleandmaleofH.rhamnoides
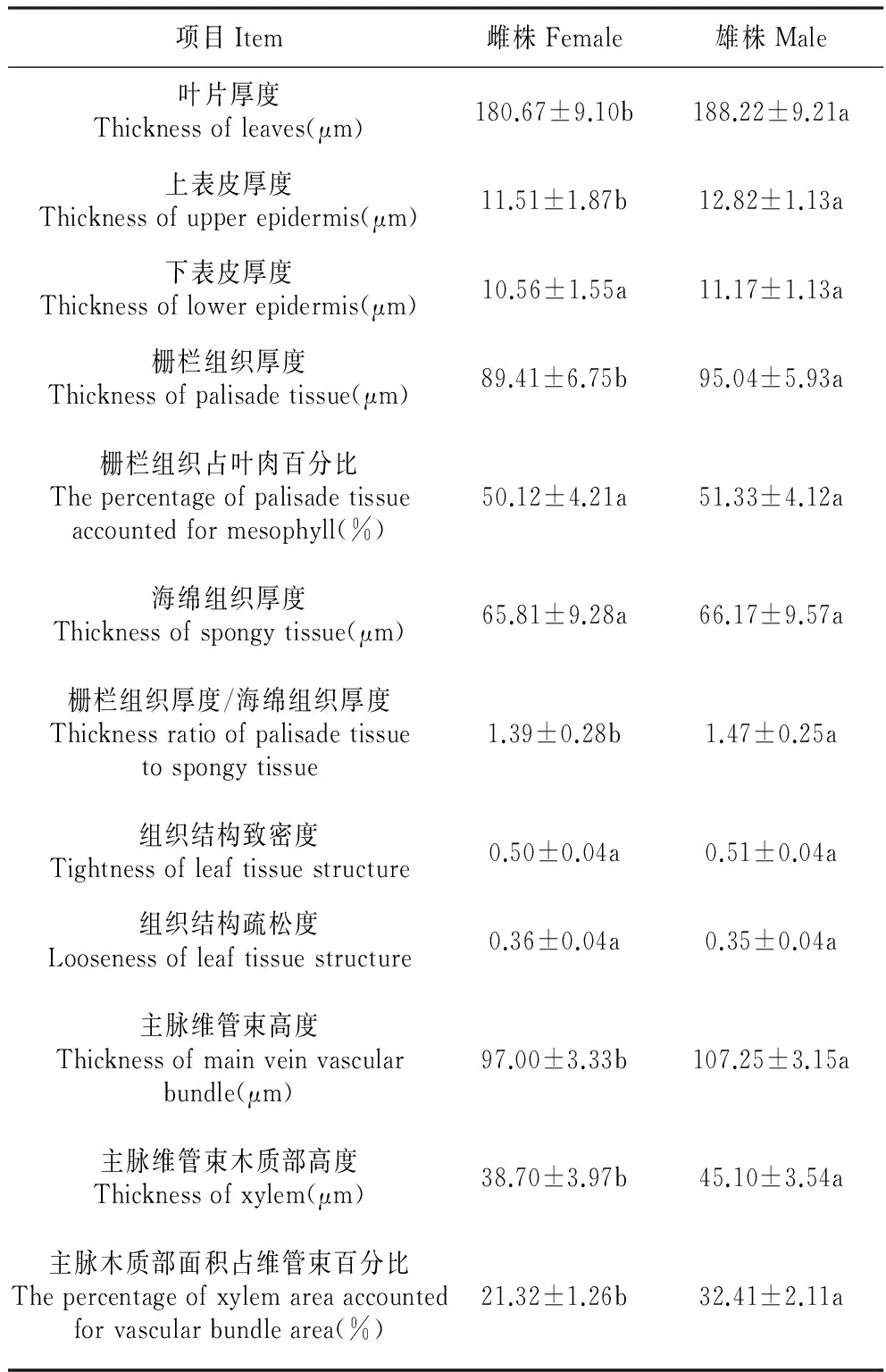
项目Item雌株Female雄株Male叶片厚度Thicknessofleaves(μm)180.67±9.10b188.22±9.21a上表皮厚度Thicknessofupperepidermis(μm)11.51±1.87b12.82±1.13a下表皮厚度Thicknessoflowerepidermis(μm)10.56±1.55a11.17±1.13a栅栏组织厚度Thicknessofpalisadetissue(μm)89.41±6.75b95.04±5.93a栅栏组织占叶肉百分比Thepercentageofpalisadetissueaccountedformesophyll(%)50.12±4.21a51.33±4.12a海绵组织厚度Thicknessofspongytissue(μm)65.81±9.28a66.17±9.57a栅栏组织厚度/海绵组织厚度Thicknessratioofpalisadetissuetospongytissue1.39±0.28b1.47±0.25a组织结构致密度Tightnessofleaftissuestructure0.50±0.04a0.51±0.04a组织结构疏松度Loosenessofleaftissuestructure0.36±0.04a0.35±0.04a主脉维管束高度Thicknessofmainveinvascularbundle(μm)97.00±3.33b107.25±3.15a主脉维管束木质部高度Thicknessofxylem(μm)38.70±3.97b45.10±3.54a主脉木质部面积占维管束百分比Thepercentageofxylemareaaccountedforvascularbundlearea(%)21.32±1.26b32.41±2.11a
注:表中小写英文字母代表5%水平的差异显著性。
Note:The small letters shown in the table indicate significant difference at the 0.05 level.

图1 沙棘雌株(A,C,E,G)与雄株(B,D,F,H)叶片解剖结构 A.雌株叶片主脉横切,示表皮、厚角组织、维管束、形成层、薄壁组织和表皮毛;B.雄株叶片主脉横切,所示同A;C.雌株叶片主脉维管束放大图,示维管束结构、韧皮部薄壁细胞及其下方薄壁细胞的后含物;D.雄株叶片主脉维管束放大图,所示同C;E.雌株叶片中部横切,示栅栏组织、海绵组织、上表皮中的拟泡状细胞(箭头)、下表皮中的细胞后含物;F.雄株叶片中部横切,所示同E;G.雌株叶缘横切,示上表皮中的拟泡状细胞(箭头)、上下表皮细胞后含物、表皮毛;H.雄株叶缘横切,所示同GFig.1 Leaf anatomical structure in the female(A,C,E,G)and the male(B,D,F,H) of H.rhamnoides A. Cross section of leaf main veins in female plant,showing epidermis,collenchyma,vascular cambium,parenchyma and trichomes; B. Cross section of leaf main veins in male plant,showing the same as A; C. Enlarged view of leaf main vein vascular bundles in female plant,showing vascular bundle structure and ergastic substance in some parenchymal cells of phloem and below the phloem; D. Enlarged view of leaf main vein vascular bundles in male plant,showing the same sa C; E. Cross section of central leaf in female plant,showing palisade tissue,sponge tissue,similar bulliform cell in upper epidermis(arrow) and ergastic substance in the lower epidermis cell; F. Cross section of central leaf in male plant,showing the same as E; G. Cross section of leaf margin in female plant,showing many similar bulliform cell in upper epidermis(arrow) and plenty of ergastic substance in upper and lower epidermis cell and trichomes; H. Cross section of leaf margin in male plant,showing the same as G
3.2.2 雌、雄株叶肉组成比较分析
由表1可以看出,雄株叶片厚度比雌株大4.0%(P<0.05),栅栏组织厚度比雄株大5.9%(P<0.05),栅栏组织厚度/海绵组织厚度比雌株大5.4%(P<0.05)。雌雄株栅栏组织占叶片厚度的百分比、海绵组织厚度、组织结构致密度和组织结构疏松度差异不显著(P>0.05)(表1)。
3.2.3 雌、雄株主脉维管束比较分析
由表1可看出,雄株主脉维管束高于雌株9.6%(P<0.05),木质部高于雌株14.2%(P<0.05),木质部面积占维管束面积比例比雌株大34.2%(P<0.05),这说明沙棘雄株主脉维管束较雌株更发达。
4 讨论
本研究发现,虽然沙棘雌雄株叶片解剖结构组成相似,下表皮密被表皮毛,栅栏组织与海绵组织分化程度明显,主脉韧皮部部分及其下方一些薄壁细胞中含有大量后含物,具有明显的旱生特性,但是,雌株上表皮含有较多的拟泡状细胞,下表皮细胞后含物丰富;雄株叶片厚度、上表皮厚度、栅栏组织厚度/海绵组织厚度均大于雌株,且其主脉维管束更发达,雌雄株叶片解剖结构表现出了显著的分化。
植物器官的形态结构是对其生理功能和生长环境长期适应的一种遗传特性,叶片长期暴露在外,是进行光合作用的主要器官,也是生态系统中的初级生产者,其形态结构上的变异性和敏感性是最大的,即对环境因子的反映是最为明显的,因此可从一定程度上反映植物的抗逆性[16]。
沙棘叶片上的表皮毛,具有反射阳光,减少叶片表面空气的流动,降低蒸腾作用,防止水分过度丧失的功能。下表皮的表皮毛尤为浓密,整个表面犹如被不同高度的伞遮盖着,因而在下表皮表面形成了一个特殊的小空间,称为“微气候区”[23],能有效地防止叶片水分的过度蒸腾,有利于抵抗干旱胁迫。拟泡状细胞是植物局部性演化中特化的结构,此结构能增加植物体含水量,并减弱和抵御强辐射对叶肉的伤害,它是提高植物抗旱性和抵御不良环境的结构,是自然选择中植物表现的有利变异之一。这与高鹤[24]在向海李叶解剖结构中发现的拟泡状细胞相似。本研究中,发现雌株上表皮含有较多的拟泡状细胞,说明雌株抗逆性更强。雄株较大的上表皮厚度,有利于在干旱胁迫下叶片的保水性能的维持。一般认为,栅栏组织及叶片越厚,栅栏组织与海绵组织的比值越大,控水能力越强,其耐旱性越强。本研究发现,沙棘雄株叶片厚度、栅栏组织厚度、栅栏组织与海绵组织比值均高于雌株,这表明,在这个意义上,沙棘雄株的耐旱性更强。
雄株更耐旱的结论在其他木本植物和草本植物中也得到了印证。Dawson等人[25]的研究结果表明,在干旱条件下,梣叶槭雄株的水分利用效率比雌株更高;在干旱条件胁迫下,酒神菊树雄株的水分利用效率明显高于雌株[8];在人工模拟和田间干旱生境下,枸骨叶冬青雄株均比雌株具有更高效的瞬时水分利用率与长期水分利用率[26]。李德颖[27]对野牛草进行干旱处理,利用电导法和压力室法,测定野牛草雌、雄株在干旱胁迫高峰时电导率和水势等指标差异,实验结果均表明其雄株比雌株具有更强的保水能力和更高的水分利用效率。所以,对沙棘而言,在干旱条件下,雄株比雌株具有更高的水分利用效率和更强的保水能力是可能的。
主脉维管束主要是为叶片输导水分、无机盐和有机养料等。沙棘雄株主脉维管束较发达发达,显示了较强的水分和无机盐的输导能力,这与郭学民等[28]对美国红梣雄株和雌株茎的次生木质部导管比较研究结果一致,即美国红梣雄株茎导管分子具长导管性、宽导管性、梯纹、网纹导管和单穿孔等特征,较雌株输导水分和无机盐的效率更高。
雌雄异株植物对光、水分、养分等非生物因素也存在种内竞争,因此雌雄个体在适应自然界的过程中会采取不同的生态对策,其中之一就是形态解剖结构的不同。沙棘雌株叶片主脉韧皮部薄壁细胞及其下方一些薄壁细胞、下表皮细胞含有更为丰富的后含物,其上表皮拟泡状细胞更多,下表皮毛也较雄株浓密,表现出耐旱特征;而雄株叶片厚度、上表皮厚度、栅栏组织厚度/海绵组织厚度均大于雌株,且其主脉维管束更发达,也表现出耐旱的特性。正是由于这种不同,才使沙棘成为对高温、干旱、风沙等各种复杂生境适应力极强的生态果树,这是长期自然选择与自身多方面变异、不断进化的结果。
有研究表明,土壤肥沃的时,雄性植株分配更多的资源到繁殖器官,而在土壤贫瘠时,雄株分配到生殖的资源锐减,但雌株却呈增加的趋势[29]。这种因种内资源竞争而产生的不同分配方式,不仅提高了植物对不同环境的适应度,还使其拥有了更优质的生长特性与繁殖策略。沙棘雌、雄株叶片解剖结构的差异正是其适应各种环境、使种族得以延续的策略的表现。
沙棘雌、雄株叶片解剖结构的差异也反映了沙棘的第二性征。第二性征是指从非生殖器官来分辨一个物种的性别特征,主要为习性、形态学特征、营养生长与繁殖、种群中性比率、生态位及物候期等方面的不同[30]。沙棘雌雄株叶片解剖结构的第二性征主要体现在各项指标大小的差异和结构上的细微差别上。
总之,沙棘叶片的旱生结构可能是沙棘拥有强大适应能力,并被誉为防风固沙、保持水土、改善生态环境优良树种的重要解剖学原因。
1.陈娟,李春阳.漫谈雌雄异株植物[J].生命世界,2015(11):40-45.
Chen J,Li C Y.Talking about dioecious plants[J].Life World,2015(11):40-45.
2.Renner S S,Ricklefs R E.Dioecy and its correlates in the flowering plants[J].American Journal of Botany,1995,82(5):596-606.
3.严欢欢,肖娟,杨永清.基于文献计量的植物雌雄异株研究进展[J/OL].广西植物,http://www.cnki.net/kcms/detail/45.1134.Q.20160328.1624.012.html.
Yan H H,Xiao J,Yang Y Q.Bibliometric analysis of research on dioecious Plant[J/OL].Guihaia,http://www.cnki.net/kcms/detail/45.1134.Q.20160328.1624.012.html.
4.何梅,孟明,施大伟,等.雌雄异株植物对干旱胁迫响应的性别差异[J].植物资源与环境学报,2015,24(1):99-106.
He M,Meng M,Shi D W,et al.On gender difference of dioecious plant in response to drought stress[J].Journal of Plant Resources and Environment,2015,24(1):99-106.
5.陈娟,李春阳.环境胁迫下雌雄异株植物的性别响应差异及竞争关系[J].应用与环境生物学报,2014,20(4):743-750.
Chen J,Li C Y.Sex-specific responses to environmental stresses and sexual competition of dioecious plants[J].Chinese Journal of Applied and Environmental Biology,2014,20(4):743-750.
6.吉晟男.圈养普氏野马(Equusferusprzewalskii)行为与生理生态及适应性的非损伤性研究[D].北京:北京林业大学,2013.
Ji S N.Non-invasive study of the behavioral and physiological ecology adaptation in captive Przewalski’s Horse(Equusferusprzewalskii)[D].Beijing:Beijing Forestry University,2013.
7.陈中海,陈晓静.雌雄异株果树的性别决定及性别鉴定的研究进展[J].福建农业大学学报,2000,29(4):429-434.
Chen Z H,Chen X J.Progress of study on sex determination and sex identification in dioecious fruit trees[J].Journal of Fujian Agricultural University,2000,29(4):429-434.
8.Espírito-Santo M M,Madeira B G,Neves F S,et al.Sexual differences in reproductive phenology and their consequences for the demography ofBaccharisdracunculifolia(Asteraceae),a dioecious tropical shrub[J].Annals of Botany,2003,91(1):13-19.
9.杨赵平,刘琴,李志军.胡杨雌雄株叶片的比较解剖学研究[J].西北植物学报,2011,31(1):79-83.
Yang Z P,Liu Q,Li Z J.Leaf blade comparative anatomy between the female and the male ofPopuluseuphraticaOliv[J].Acta Botanica Boreali-Occidentalia Sinica,2011,31(1):79-83.
10.胥晓,杨帆,尹春英,等.雌雄异株植物对环境胁迫响应的性别差异研究进展[J].应用生态学报,2007,18(11):2626-2631.
Xu X,Yang F,Yin C Y,et al.Research advances in sex-specific responses of dioecious plants to environmental stresses[J].Chinese Journal of Applied Ecology,2007,18(11):2626-2631.
11.刘金平,游明鸿,段婧,等.水分胁迫下雌雄异株植物葎草繁殖策略的可塑性调节[J].草业学报,2015,24(3):226-232.
Liu J P,You M H,Duan J,et al.Plasticity of reproductive strategy of dioeciousHumulusscandensin response to variation in water deficit stress[J].Acta Prataculturae Sinica,2015,24(3):226-232.
12.何士敏,汪建华,秦家顺.几种沙棘叶片组织结构特点和抗旱性比较[J].林业科技开发,2009,23(1):16-19.
He S M,Wang J H,Qin J S.Comparison of tissue structure characteristics and drought-resistant properties in someHippophaespeciesleaves[J].China Forestry Science and Technology,2009,23(1):16-19.
13.王官,韩有志,安雄韬,等.不同品种大果沙棘叶片与果实特征的研究[J].山西林业科技,2013,42(1):32-34.
Wang G,Han Y Z,An X T,et al.Study on characteristics of leaf and berry of different large-berry cultivars of seabuckthorn[J].Shanxi Forestry Science and Technology,2013,42(1):32-34.
14.王国富,李连国,李晓燕,等.沙棘叶片表面形态特征与抗旱性的关系[J].园艺学报,2006,33(6):1310-1312.
Wang G F,Li L G,Li X F,et al.Relationship between the morphological characteristics of leaf surface and drought resistance of Sea Buckthorn[J].Acta Horticulturae Sinica,2006,33(6):1310-1312.
15.潘志辉.沙棘叶片形态特征的数学分析[J].安徽农业科学,2013,41(12):5408-5409.
Pan Z H.The mathematical analysis of leaf morphological characteristics ofHippophaerhamnoideL.[J].Journal of Anhui Agricultural Sciences,2013,41(12):5408-5409.
16.潘志辉.沙棘叶片表面形态特征与抗旱性分析[J].安徽农业科学,2014,42(21):7054-7056.
Pan Z H.The Morphological characteristics and drought resistance analysis on leaf surface ofHippophaerhamnoidesL.[J].Journal of Anhui Agricultural Sciences,2014,42(21):7054-7056.
17.吴林,霍焰,聂小兰,等.沙棘叶片组织结构观察及其与抗旱性关系的研究[J].吉林农业大学学报,2003,25(4):390-393.
Wu L,Huo Y,Nie X L,et al.Studies on leaf tissue parameters and their relations to drought resistance of sea buckthorn[J].Journal of Jilin Agricultural University,2003,25(4):390-393.
18.李小燕,王林和,李连国,等.沙棘叶片组织解剖构造与其生态适应性研究[J].干旱区资源与环境,2006,20(5):209-212.
Li X Y,Wang L H,Li L G,et al.A study on the relation between tissue structure and ecological stability in seabuckthorn leaf[J].Journal of Arid Land Resources and Environment,2006,20(5):209-212.
19.赵延霞,骆有庆,宗世祥,等.不同沙棘品种雌雄株叶片解剖结构及抗旱性比较[J].北京林业大学学报,2012,34(6):34-41.
Zhao Y X,Luo Y Q,Zong S X,et al.Leaf anatomical structure and drought resistance of different sex and varieties of sea buckthorn[J].Journal of Beijing Forestry University,2012,34(6):34-41.
20.江川,罗大庆,王立辉.西藏半干旱区5种灌木叶片结构的抗旱特征研究[J].西北林学院学报,2011,26(4):13-17.
Jiang C,Luo D Q,Wang L H.Drought-resistant characteristics of leaf structures of five shrubs in semi-arid region of Tibet[J].Journal of Northwest Forestry University,2011,26(4):13-17.
21.Li C Y,Xu G,Zang R G,et al.Sex-related differences in leaf morphological and physiolo-gical responses inHippophaerhamnoidesalong an altitudinal gradient[J].Tree Physiology,2007,27(3):399-406.
22.李华西,曹迎春.小五台自然保护区野生经济植物[J].河北林业科技,1998(2):34-36.
Li H X,Cao Y C.Wild economic plants in Xiaowutai Nature Reserves[J].The Journal of Hebei Forestry Science and Technology,1998(2):34-36.
23.李代琼,梁一民,黄瑾,等.半干旱黄土区沙棘的形态解剖学特性研究[J].沙棘,2004,17(1):8-13.
Li D Q,Liang Y M,Huang J,et al.Study on the characteristic of anatomy of seabuckthorn on the semiarid loess region[J].Hippophae,2004,17(1):8-13.
24.高鹤.吉林省不同生态环境四种蔷薇科植物比较结构研究[D].长春:东北师范大学,2005.
Gao H.Comparative structure study on four species of Rosaceae palnts living in different ecosystem in Jilin[D].Changchun:Northeast Normal University,2005.
25.Dawson T E,Ehleringer J R.Gender-specific physiology,carbon isotope discrimina- tion,and habitat distribution in boxelder,Acer negundo[J].Ecology,1993,74(3):798-815.
26.Obeso J R,Retuerto R.Sexual dimorphism in holly Ilex aquifolium:cost of reproduce- tion,sexual selection or physiological differentiation[J].Revista Chilena de Historia Natural,2002,75(1):67-77.
27.李德颖.野牛草雌雄单性植株对水分胁迫反应的差异[J].园艺学报,1996,23(1):62-66.
Li D S.Different response to water deficit in male and female plants of Buffalo grass[J].Acta Horticulturae Sinica,1996,23(1):62-66.
28.郭学民,高忠明,刘振林,等.美国红梣雄株和雌株茎导管分子的形态解剖比较[J].林业科学,2010,46(8):51-55.
Guo X M,Gao Z M,Liu Z L,et al.Comparative anatomy of vessel elements in staminate and pistillate Plants ofFraxinuspennsylvanica[J].Scientia Silvae Sinicae,2010,46(8):51-55.
29.Hesse E,Pannell J R.Sexual dimorphism in a dioecious population of the wind- pollinated herbMercurialisannua:the interactive effects of resource availability and competition[J].Annals of Botany,2011,107(6):1039-1045.
30.杨好伟,夏国军,刘万代.植物的第二性征[J].河南科学,1996,14(S):137-138,165.
Yang H W,Xia G J,Liu W D.Secondary sex characters in plants[J].Henan Science,1996,14(S):137-138,165.
Natural Science Foundation of Hebei Province(C2014407077)
introduction:LI Na(1991—),female,master,mainly engaged in the study of plant developmental cell biology.
date:2016-09-14
ComparisononLeafAnatomicalStructuresbetweentheFemaleandtheMalePlantsofEippophaerhamnoidesL.
LI Na GUO Xue-Min*MENG Yang
(College of Life Science & Technology,Hebei Normal University of Science & Technology,Changli 066600)
We observed the differences in leaf anatomical structure between the male and the female using paraffin method to investigate leaf secondary sex characteristics ofHippophaerhamnoidesL.. Results showed that: (1)Both the male and the female leaves consisted of epidermis, mesophyll and veins. The epidermis was composed of a layer of cells with developed trichomes, including similar bulliform cells in upper epidermis; the differentiation of palisade tissue and spongy tissue was obvious in mesophyll. (2)There were more similar bulliform cells in upper epidermis, more ergastic substances in some parenchymal cells of phloem and below the phloem, more trichomes in lower epidermis in the female plant than in the male one; the male plant were significantly higher than the female in leaf thickness, thickness of leaf upper epidermis, thickness of palisade tissue, thickness ratio palisade tissue/sponge tissue, and had more developed vascular bundle than the female in main vein. These results suggested that there were obvious differences in leaf anatomical structure between the female and the male plants of the species, and the differences were the performances of secondary sex characteristics, also robust adaptive strategy formed in the long-term evolution, which might be good for reproduction of the species.
HippophaerhamnoidesL.;dioecism;leaf blade;comparative anatomy
河北省自然科学基金资助项目(C2014407077)
李娜(1991—),女,硕士研究生,主要从事植物发育细胞生物学。
* 通信作者:E-mail:xueminguo@126.com
2016-09-14
* Corresponding author:E-mail:xueminguo@126.com
S793.6
A
10.7525/j.issn.1673-5102.2017.01.001
猜你喜欢
安徽农业科学(2024年5期)2024-04-20 11:38:27
西北园艺·果树(2024年1期)2024-04-01 09:10:14
南方农业学报(2021年2期)2021-06-30 01:56:03
湖南文理学院学报(自然科学版)(2020年2期)2020-05-08 08:00:14
园林科技(2020年1期)2020-01-18 05:13:00
新课程·小学(2019年7期)2019-09-17 12:58:34
江苏农业科学(2019年21期)2019-01-03 02:05:58
现代农业科技(2018年17期)2018-10-13 12:31:24
现代农业科技(2009年7期)2009-07-29 07:11:32
现代农业科技(2009年20期)2009-03-20 05:27:10
