
有翅桃蚜模拟色板参数的单纯形优化
2017-10-25 00:49李为争付国需原国辉郭线茹
河南农业大学学报 2017年4期
常 虹,李为争,胡 璞,付国需,原国辉,郭线茹
(1.商丘职业技术学院,河南 商丘 476000;2.河南农业大学植物保护学院,河南 郑州 450002)
有翅桃蚜模拟色板参数的单纯形优化
常 虹1,李为争2,胡 璞2,付国需2,原国辉2,郭线茹2
(1.商丘职业技术学院,河南 商丘 476000;2.河南农业大学植物保护学院,河南 郑州 450002)
采用改进的单纯形优化法定量地优选了有翅桃蚜计算机模拟色彩诱捕器的2个参数:色调偏角(定义为靶标颜色与纯红色矢量顺时针方向的夹角)和颜色强度(定义为靶标颜色离三色坐标系原点之间的距离,取值范围为0~255之间的自然数)。当颜色强度值为10、25、50、100和200时,桃蚜相对平均位移出现最大值的色调偏角分别为190°、85°、35°、100°和345°。随着光强增加,有翅桃蚜对不同色调的选择性逐渐下降,即表现出一定的“炫目效应”。单纯形优化结果显示,色调偏角与颜色强度的最佳组合为(θ=134.38°,Ib=61.25)。本试验的生物测定方法可以在其他小体日出性昆虫的趋向行为研究中借鉴。
桃蚜;模拟混色;趋光性;单纯形优化法
桃蚜Myzuspersicae(Sulzer)是世界范围内最重要的植物病毒媒介昆虫之一[1]。有翅型个体能定殖到大量寄主植物上,通常造成寄主植物的水分胁迫、叶片皱缩、生长发育延缓,排泄的蜜露还能造成农产品的污染[2]。在桃蚜点片发生阶段,采用粘胶诱捕器捕获迁飞个体,是前景较好的预防植物病毒传播的措施[3]。无翅桃蚜趋向于波长范围为520~580 nm,在这个波段范围内,靶标物体表面反光越强,引诱力越强[4]。在有翅桃蚜方面的测定也得到类似的结果[5]。但是多数大田诱捕试验难以获得足够多光谱性质各异的有机染料来制造粘胶色板,故多数对桃蚜的趋光性特征侧重于定性的而不是定量的描述。计算机绘图软件与高分辨率显示器相结合,可以数字化显示人类可见光范围内的一切颜色。然而,颜色知觉是由许多相互作用的参数决定的,如色调、亮度、饱和度等。所以在蚜虫趋光性研究中,需要较多的生物测定次数。单纯形是一种用于多变量优化的序贯性仿生算法。假设待优化的有N个变量,首先任意寻找N+1个不同的变量组合并进行测试,这些组合条件构成了N+1个顶点的超几何多面体,其中每个顶点对应于一组待优化的变量组合。测试完成后,对单纯形各顶点上的反应值进行排序,找到反应最差的顶点,并计算剩下的N个顶点所在的形心点在超几何多面体中的坐标。然后,计算最差顶点关于形心点的反射点的坐标,测试反应值。以此类推,每次插入一个试验点,单纯形逐渐循环推移下去[6-7]。有时在一轮测试中,最差点及其反射点均不理想,单纯形会进入死循环。为此,一些学者改进了基本的单纯形算法,加入了单纯形的扩张、内收缩和外收缩步骤,单纯形法才真正成为一种比较实用的仿生学算法,是为“变化步长的单纯形方法”[8]。针对本文提出的问题,计算机混色的方法可以精确地确定桃蚜的趋光反应特征,但是涉及的参数很多而且参数之间存在复杂的相互作用,非常适合用变化步长的单纯形法来优化。为此,作者用计算机显示器直接作为光源研究了有翅桃蚜的趋光性特征,利用变化步长的单纯形法优化了桃蚜虚拟色彩诱捕器的有关参数。
1 材料与方法
1.1 供试昆虫
有翅桃蚜于2015年9月采集于河南农业大学科教园区的烟田。
1.2 混色设备与混色原理
HP Dreamcolor LP2480zx型显示器用于提供供试颜色。预先设置如下:亮度40%,对比度40%,色温6 500 K,分辨率1 920 dpi×1 200 dpi,颜色质量32 bit,屏幕刷新率85 Hz。在Cartesian颜色坐标系统中,色调偏角(θ)定义为从纯红色向量轴顺时针方向形成的夹角,光强(Ib)定义为靶标颜色离坐标原点的距离(图1)。一种颜色的RGB数值计算如下:




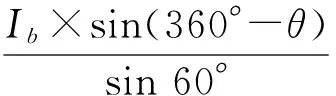

式中:r、g和b分别表示某个象限中一种待测颜色需要的红、绿、蓝3种参数的具体数值,取值范围均为[0,255]。

注:虚线环上的每个点表示光强相同而色调不同的颜色系列。Note: Every point on the dotted circle indicates a color serial with equal intensity (Ib) but different hues.
1.3 趋光反应装置
趋光反应装置由1根不透明钙塑外套管(51 cm L×2.5 cm ID×3.0 cm OD)和1根精确标定长度的有机玻璃内衬管(52 cm L×3.0 cm ID×4.0 cm OD)组成,其中内衬管的两端有透明的内嵌式盖子。内衬管内部为生测室,光源相反的一端留出5 cm的长度作为释放试虫的空间,然后以1 mm的精度标记45 cm长的蚜虫爬行空间(图2)。
1.4 生物测定方法
在Adobe photoshop CS2绘图软件中创建一个32 bit、RGB模式的画布(“自定义”,72像素/英寸,白色背景,50%亮度),用笔刷工具(“正常模式”,硬度=100%,不透明度 = 100%,流量=100%)将一种待测颜色、参照色和对照色分别涂布在画布适当的位置。参照色指的是用数码相机获得的桃蚜经常取食的烟草顶叶部位的超级微距图片。对照色指的是全黑背景(R=G=B=0),用于扣除缺乏光刺激的情况下桃蚜的自发性运动。每次测试同时使用三套相同的生测装置,在每根内衬管的试虫释放区释放15头有翅桃蚜,盖上盖子后迅速推入外套管,并分别朝向垂直放置的显示器上绘图画布的对照色、参照色和待测颜色区域。10 min后将内衬管抽出,立刻根据内衬管壁上的标尺记录所有被试的位移。为了避免桃蚜告警素、蜜露等产物的影响,相邻两次测试间隔期,用蒸馏水清洁内衬管内壁,并用2 100 W Philips HP 4983头发烘干器对准内衬管的一端吹3 min,冷却至室温后进行下一次测定。

图2 桃蚜趋光性的生物测定装置Fig.2 Experimental setting for testing phototactic response of M.persicae
1.5 优化方法
为了构建初始单纯形,将待测颜色的光强水平设置为10、25、50、100、200共5组,每种光强下再按5°的间隔划分为72种待测颜色,共设计了360种待测颜色。由于显示器中人类视觉三原色的数值可以设置的范围为[0,255],故上述5个光强水平的设计涵盖了计算机显示器能够达到的大部分光强范围。根据上述颜色坐标系中不同象限的计算方程,将四舍五入后的R、G、B值输入Adobe photoshop CS2软件的调色板中,再用笔刷工具将待测颜色涂布在画布上进行测定。每个待测颜色重复测定3次,每次测试更换新的被试。用相对平均位移(Relative Mean Displacement,RMD)[9]作为衡量蚜虫趋光反应强弱的标准:
RMD=(MDb-MDCK)/(MDref-MDCK)。
式中:MDb指的是桃蚜趋向靶标颜色的平均位移,MDref指的是桃蚜趋向参照颜色的平均位移,MDCK指的是桃蚜趋向对照颜色的平均位移。
根据上述测试结果,选择(85°,25)、(35°,50)和(190°,10)作为初始单纯形的3个顶点。当单纯形收敛域(Convergence Domain)达到ε=0.04时终止。反射系数、扩张系数、外收缩系数和内收缩系数分别设置为1、2、0.5和0.5。Ib值小于0的顶点超出了色域的边界,将它们的色调偏角值加上或者减去180°,使得其Ib恒为正值。当色调偏角超出了[0°,360°)范围时,加上或减去360°进行转换。从Cartesian色系角度看,这些转换得到的是等价的结果。这种优化方法不涉及文献常用的统计分析方法,因为是根据每个测试点响应值的大小进行单纯形推移优化,每个组合条件仅测试一次。理论上讲,如果某一次测试出现了反常的结果,总可以通过单纯形的推移迅速矫正。随着优化过程的进行,离最优的条件组合越来越近,试验指标变异越来越小。这时对接近最优的组合条件进行若干次重复验证,观察得到的结果是否稳健(用单纯形收敛域ε的值来衡量稳健性)。
2 结果与分析
2.1 相对平均位移反应谱
图3总结了桃蚜暴露于不同待测颜色下的相对平均位移。在不同的光强度下,桃蚜最偏好的色调是不同的。当光强值分别为10、25、50、100和200时,最优的色调偏角分别为190°、85°、35°、100°和345°,相应的相对平均位移分别为1.96、2.74、2.27、1.79和1.53。
随着光强的增加,有翅桃蚜对不同色调的趋向反应选择性越低。如图3所示,当光强值为10和25时,桃蚜的色调反应是相似的,但是前者出现了许多尖削的峰,而后者在这些峰附近扩展为3个敏感的色谱反应范围:75°~120°,145°~170°,和 185°~220°。当光强值为50时,引诱力最强的色调范围偏向了黄色和紫色。当光强值高达100和200时,则看不到明显的反应峰,表现出一定的“炫目效应”。
2.2 单纯形优化结果
根据上述结果,选择下述3个组合变量组作为初始单纯形的顶点:(85°,25),(35°,50)和(190°,10)。单纯形的推移、顶点的变量值、顶点对应的RGB值和靶标颜色引起桃蚜的平均位移值如表1所示。初始单纯形中顶点1的响应值最好(18.26),但是响应值最差的顶点2的反射点和外收缩点都没有得到满意的结果,因此寻找次差的顶点3的反射点进行测试。在第3轮循环中,原始单纯形的顶点1重新测定,因为在3轮测试中这个顶点总是保留下来。重测值为19.08,与该顶点在原始单纯形中得到的测试只有4%的差异。在第4和第6轮循环中,我们对单纯形进行整体收缩来促进其收敛。共经过8轮循环20次测定,在组合条件(134.38°,61.25)处得到了最优反应值,处于单纯形设置的收敛域范围之内。另外,经过4次测定,发现这个最优结果是稳健的。

注:曲线上的每个点离坐标中心的距离表示相对平均位移,外围的角度值为色调偏角,图中部的环线表示相对平均位移为1。该环内部的数据点表示相应的待测颜色引诱活性不比参照色强,反之亦然。
Note: The distances from the origin of coordinate indicateRMDs, and the angles in each subfigure indicate θ values. The mid-circle in each subfigure indicates meanRMD=1, any points inside the circle indicate the test color elicited lower attractive response than that of the reference, and verse visa.
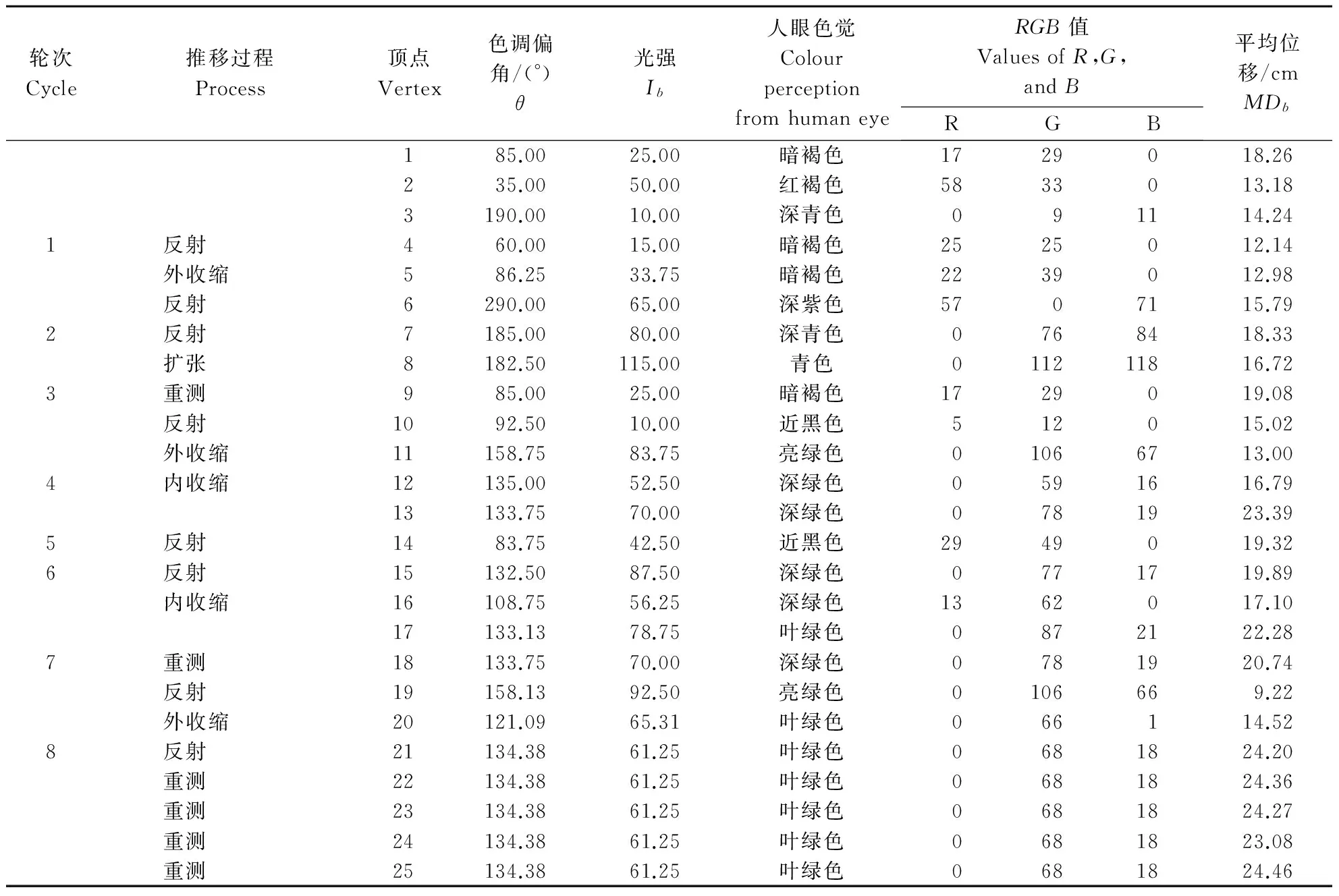
图3 有翅桃蚜暴露于5种光强值不同色调下的相对平均位移Fig.3 Relative mean displacement profiles of alate M.persicae exposed to blended colors under different intensities
3 讨论
颜色是植食性昆虫寄主选择过程中经常利用的外界刺激信息之一[10]。不同昆虫种类甚至同一种类的不同性别,也会表现出不同的颜色偏好性[11-12]。根据本研究的单纯形优化结果,色调偏角为134.38°、光强为61.25的供试颜色对桃蚜的引诱力最强。另外,我们发现高强度的光线会造成有翅桃蚜彩色视觉系统的“炫目效应”,而以前报道高强度的光线下昆虫对色调的分辨力增强[5]。
昆虫具备2-5种不同类型的光感受器,通常是UV-G-B模式的三色系统,用于感应不同波长的光子。果蝇对绿色果实的视觉检测,是基于对绿色敏感的光感受器和对蓝色敏感的光感受器之间的拮抗性相互作用实现的,这可以解释许多植食性昆虫对黄色刺激的偏好性比绿色更强的现象[10]。一般绿色叶片反射最强的波长范围为500~600 nm (峰值为550 nm),黄色可能是绿叶反射谱中反射光最强的颜色[13-14]。本研究测得的引诱力最强的颜色处于绿色和青色之间。尽管桃蚜趋向黄色和绿色均有报道,但显然大田应用中最常使用的仍然是黄板。出现测试结果差异的潜在原因有两个:一是以往的测试是计算不同颜色的色板放置在自然界较长一段时间之后蚜虫的分布情况,这些色板是与蓝色天空、绿色植被、褐色土壤等自然背景混杂在一起的,而本研究的试验指标是特定时间内蚜虫群体趋向视野中单一颜色的速度;二是以往的测试使用的不是光源而是物体表面的反射色,而本研究使用的是显示器呈现的光源。尽管如此,本试验得到的结果对于桃蚜防治实践仍然具有重要的意义。显然,光源和色板相比,投射距离远,辐射面积大,夜间也能被有翅桃蚜知觉,受叶片等障碍物影响小,耐候性强。如能设计出本文最优化颜色参数的冷光型LED光源,并研制与其配套的无色粘虫胶,有希望改善基于蚜虫趋光性的防治措施的效果。本文的研究目的,是期望在室内条件下,排除自然干扰因子,科学地比较桃蚜对精密控制参数的颜色的趋向反应。未来需要在模拟自然背景色彩的环境中,验证本文最优化的颜色对蚜虫的引诱作用,此时适合采用付国需等[15]测试“绿叶-待测颜色竞争系数”的生物测定装置,并与该文献采用正交设计最优化的色彩参数进行比对。
尽管许多节肢动物的彩色视觉系统为UV-G-B模式,显示器并不能够释放出足够强度的紫外光,但针对日出性的小体昆虫而言是非常适宜的,因为在它们的视野中主要是绿色植物等发出的反射光,也不带有很强的紫外线。一些学者已经构建了RGB颜色值与波长之间的对应函数关系[16-17],为农业害虫最优色板的定量化设计提供了很好的前景。
[1] NAMBA R, SYLVESTER E S. Transmission of cauliflower mosaic virus by the green peach, turnip, cabbage, and pea aphids [J]. Journal of Economic Entomology, 1981, 74(5): 546-551.
[2] MCLEOD P J, STEINKRAUS D C, CORRELL J C, et al. Prevalence ofErynianeoaphidis(Entomophthorales: Entomophthoraceae) infections of green peach aphid (Homoptera: Aphididae) on spinach in the Arkansas River Valley [J]. Environmental Entomology, 1998, 27(3): 796-800.
[3] NIAZ T, AYUB M. Population pattern ofMyzuspersicaeon potato crop at aisalabad [J]. Journal of Agricultural Research, 2007, 45(4): 305-311.
[4] HODGSON A J, ELBAKHIET I B. Effect of colour and shape of 'target' hosts on the orientation of emigrating adult apterousMyzuspersicaein the laboratory [J]. Entomologia Experimentalis et Applicata, 1985, 38(3): 267-272.
[5] COSTELLO M J. Spectral reflectance from a broccoli crop with vegetation or soil as background: influence on immigration by Brevicoryne brassicae andMyzuspersicae[J]. Entomologia Experimentalis et Applicata, 1995, 75(2): 109-118
[6] BOX G E P. Evolutionary operation: a method of increasing industrial productivity [J]. Applied Statistics, 1957, 6(2): 81-101.
[7] SPENDLEY W, HEXT G R, HIMSWORTH F R. Sequential application of simplex designs in optimisation and evolutionary operation [J]. Technometrics, 1962, 4(4): 441-461.
[8] NELDER J A, MEAD R. A simplex method for function minimization[J].The Computer Journal, 1965, 7(4): 308-313.
[9] 付国需,李为争,吴少英,等. 桃蚜对不同单色光趋性反应的测定 [J]. 昆虫学报,2009, 52(10): 1171-1176.
[10] KELBER A. Receptor based models for spontaneous colour choices in flies and butterflies [J]. Entomologia Experimentalis et Applicata, 2001, 99(2): 231-244.
[11] HOBACK W W, SVATOS T M, SPOMER S M, et al. Trap color and placement affects estimates of insect family-level abundance and diversity in a Nebraska salt marsh [J]. Entomologia Experimentalis et Applicata, 1999, 91(3): 393-402.
[12] ENTWISTLE P F. Some evidence for a colour sensitive phase in the flight period of Scolytidae and Platypodidae [J]. Entomologia Experimentalis et Applicata, 1963, 6(2): 143-148.
[13] HILL A R, Hooper G H S. Attractiveness of various colours to Australian tephritid fruit flies in the field [J]. Entomologia Experimentalis et Applicata, 1984, 35(2): 119-128.
[14] MENSAH R K, MADDEN J L. Field studies on colour preferences ofCtenarytainathysanurain Tasmanian boronia farms [J]. Entomologia Experimentalis et Applicata, 1992, 64(2): 111-115.
[15] 付国需,李为争,安靖靖,等. 桃蚜诱捕色板的青-洋红-黄-黑四色模式优化[J]. 昆虫知识,2010,47(2): 368-373.
[16] 李剑波,覃泽涛. 光学图样的仿真处理 [J]. 光电工程,2002,29(6): 70-72.
[17] DÖRING T F, CHITTKA L. Visual ecology of aphids-A critical review on the role of colours in host finding [J]. Arthropod Plant Interactions, 2007, 1(1): 3-16.
SimplexoptimizationofsimulatedcolorparametersofalateMyzuspersicae(Sulzer)
CHANG Hong1, LI Weizheng2, HU Pu2, FU Guoxu2, YUAN Guohui2, GUO Xianru2
(1.Shangqiu Polytecnic, Shangqiu 476000,China; 2.College of Plant Protection, Henan Agricultural University, Zhengzhou 450002, China)
Two variables of simulated color board for trappingMyzuspersicaealates, the angle starting from Red vector clockwisely (θ) and intensity of blended color (Ib), were optimized using variable-size Simplex method. The relative mean displacements of the aphids exposed to the colors with intensities of 10, 25, 50, 100, and 200, defined by Adobe photoshop CS4, respectively peaked at 190°, 85°, 35°, 100°, and 345°. With the increasing of the light intensity, alateM.persicaeshowed lower preference of phototactic response to different hues, suggesting a “dazzling effect”. Results of simplex optimization show that the optimal combination (θ,Ib) was 134.38° of the angle (θ) and 61.25 of the light intensity (Ib). This method of color sticky traps has potential application values in testing the preference behaviors of other diurnal insect pests.
Myzuspersicae;color blending simulation;phototaxis;simplex optimization
2017-02-13
郑州市农业科技创新项目(郑农财[2015]23号)
常 虹(1964-),女,河南郑州人,副教授,硕士,主要从事植物保护方面的研究。
李为争(1978-),男,河南洛阳人,副教授,博士。
1000-2340(2017)04-0541-06
S436.621
A
(责任编辑:蒋国良)
猜你喜欢
果农之友(2022年9期)2022-09-27
河北农业科学(2022年6期)2022-02-13
上海建材(2021年1期)2021-11-22
果农之友(2021年10期)2021-11-03
民用飞机设计与研究(2020年4期)2021-01-21
新课程·中学(2019年7期)2019-09-17
物理教学探讨(2018年1期)2018-02-13
爱你(2015年4期)2015-11-15
爱你·健康读本(2015年2期)2015-04-09
西北园艺(果树)(2015年6期)2015-02-23
