
北京松山国家级自然保护区不同林龄胡桃楸林下植物多样性
2017-10-10 02:38:59郑昌龙王健铭李景文
浙江农林大学学报 2017年5期
郑昌龙,王健铭,李景文
(北京林业大学 林学院,北京100083)
北京松山国家级自然保护区不同林龄胡桃楸林下植物多样性
郑昌龙,王健铭,李景文
(北京林业大学 林学院,北京100083)
以北京松山国家级自然保护区胡桃楸Juglans mandshurica林为研究对象,采用典型抽样法研究林下植物物种组成与多样性。结果表明:胡桃楸林灌木层植物种类随林龄增加而逐渐增加,草本层植物种类则逐渐减少,存在明显的时空动态;幼龄林和中龄林草本层以披针叶薹草Carex lancifolia,大油芒Spodiopogon sibiricus,草乌Aconitum carmichaeli和龙芽草Agrimonia pilosa等为优势种,近熟林和成熟林则以蓝萼香茶菜Rabdosia japonica和白屈菜Chelidonium majus等为优势种;幼龄林和中龄林灌木层以小叶鼠李Rhamnus parvifolia和荆条Vitex negundo var.heterophylla等为优势种,近熟林和成熟林则以红花锦鸡儿Caragana rosea,小花溲疏Deutzia parviflora,毛叶丁香Syringa pubescens和暴马丁香Syringa reticulata var.mandshurica等为优势种。不同林龄胡桃楸林下灌木层与草本层4个物种多样性指数存在显著差异(P<0.05):灌木层物种多样性指数中,随着林龄的增加,Simpson指数从0.598 0增加到0.787 9,Shannon-Wiener指数从1.183 9增加到1.835 6,Pielou均匀度指数从0.682 0增加到0.839 0,Margalef丰富度指数从0.979 0增加到1.614 9,均呈现连续增加的趋势;草本层均匀度指数随着林龄的增加从0.884 6增加到0.987 2,表现为连续递增趋势,其余指数中,Simpson指数从0.843 6减少到0.759 4,Shannon-Wiener指数从2.120 9减少到1.673 9,丰富度指数从1.951 7减少到1.347 1,均随着林龄的增加呈持续递减的趋势。不同林龄胡桃楸林下灌木层和草本层物种丰富度均存在极显著差异(P<0.001),由幼龄林到成熟林,灌木层物种丰富度从3.385 7逐渐增加到6.340 0,在成熟林阶段达到最高值;草本层的变化规律与灌木层呈现完全相反的趋势,随着林龄的增加,林下草本层植物的种类逐渐减少。用半变异函数模型模拟,均匀度指数和Simpson指数为球面模型,Shannon-Wiener指数和丰富度指数则更符合指数模型,研究区的物种多样性为中等的空间相关性,结构性因子所占比例为0.584~0.681,随机性因子所占比例为0.319~0.416。建议该区科学合理地规划景区建设及道路开发,并对当地居民的种子采集等活动进行管理。图4表4参23
森林生态学;胡桃楸;灌木;草本;物种多样性;北京松山
Abstract:To effectively protect the Juglans mandshurica forest and prevent it’s further habitat deterioration in Beijing,the study attempted to clarify the distribution characteristics of the undergrowth plant diversity.Shrub and herb plant species composition and diversity were researched at different forest ages of Juglans mandshurica forest in the Songshan National Nature Reserve by the typical sampling method that twenty-four 20 m×20 m plots were set up in different typical stands,and one 5 m×5 m shrub sample and one 1 m×1 m herb sample were made at each quadrangle and center of each arbor sample.Diversity analysis included richness index(M),Shannon-Wiener diversity index (H),and Simpson diversity index (D).Results showed that as forestsincreased in age,shrub species and quantities increased,and herb species and quantities decreased.The most important herb plant species in the young and middle-aged forests were Carex lancifolia,Spodiopogon sibiricus,Aconitum carmichaelii,and Agrimonia pilosa;whereas,in the nearly mature forests and mature forests Rabdosia japonica and Chelidonium majus were predominant.Shrubs were dominated in the young forests and middle-age forests by Rhamnus parvifolia and Vitex negundo var.heterophylla but in the nearly mature forests and mature forests by Caragana rosea,Deutzia parviflora,Syringa pubescens,and Syringa reticulata var.mandshurica.There were significant differences in species diversity indices between shrub layer and herb layer in Juglans mandshurica forest at different ages (P<0.05).In the shrub layer,with the increase of age,Simpson index increased from 0.598 0 to 0.787 9,the Shannon-Wiener index is 1.183 9 to 1.835 6,the Pielou evenness index is 0.682 0 to 0.839 0,and the Margalef richness index is 0.979 0 to 1.614 9,showing a continuous increase trend;In the herb layer,the Pielou evenness index increased from 0.884 6 to 0.987 2,and the Simpson index decreased from 0.843 6 to 0.759 4,and the Shannon-Wiener index decreased from is 2.120 9 to 1.673 9,while the Margalef richness index is 1.951 7 to 1.347 1,and decreased with the increase of stand age.Shrub layer and herb layer of Juglans mandshurica forest under different forest ages was significantly different (P<0.001).The species richness of shrub layer increased from 3.385 7 to 6.340 0 from young forest to mature forest,and reached the highest value in mature forest stage.The change of herb layer was opposite to that of shrub layer.With the increase of age,the species of the herb layer in the undergrowth layer decreased gradually.Using the semi-variogram model,Pielou evenness index and Simpson index were spherical model,Shannon-Wiener index and Margalef richness index were more consistent with it.The species diversity of the study area was moderate spatial correlation,the proportion of structural factors is 0.584-0.681,and the proportion of random factors is 0.319-0.416.For spatial variation of species diversity,human factors were dominant.Therefore,reasonable planning and development activities at the local level should be undertaken with local residents conducting seed collection and other management activities. [Ch,4 fig.4 tab.23 ref.]
Key words:forest ecology;Juglans mandshurica;shrub;herb;species diversity;Songshan in Beijing
植物多样性作为群落的重要特征,是生态系统结构、功能及其稳定性的基础[1-2]。随着生态学研究的不断深入,植物多样性格局及形成机制成为生态学科的热点问题之一[3-4],是理解区域生物多样性维持机制的重要内容,同时对生物多样性的保护和管理有着重要意义[5-8]。研究植物多样性,有助于认识群落的组成、变化和发展趋势,同时也可反映群落及其环境的保护状况[9-10]。胡桃楸Juglans mandshurica是北京唯一的国家保护高大乔木,在生物多样性相对匮乏的北京地区具有特殊的意义。松山国家级自然保护区是主要的胡桃楸林分布区,受到生态旅游及麻核桃Juglans hopeiensis开发等人为活动的影响,生境严重破碎化,林下群落结构遭到破坏,植物多样性受到严重威胁[11]。胡桃楸林不同演替阶段群落结构存在差异[9-10]。本研究对不同林龄阶段的胡桃楸林下植被进行调查,目的在于了解松山国家级自然保护区胡桃楸林下群落的植物多样性及变化规律,找到群落与环境因子和人为干扰等因素的互作机制,为松山国家级自然保护区多样性保护与恢复工作提供理论依据。
1 研究区概况
北京松山国家级自然保护区地处延庆县西北部,森林覆盖率接近90%。地处燕山山脉的军都山,坐标为 40°29′9″~40°33′35″N,115°43′44″~115°50′22″E[12]。属于暖温带大陆性季风气候区, 年平均气温为8.5℃,最高和最低气温分别为39.0℃和-27.3℃,年降水量与蒸发量分别为493.0 mm和1 772.0 mm,年平均日照时数为2 836.3 h。随海拔由低向高依次分布有山地褐土、山地棕壤和山地草甸土[12],土层厚度为20~55 cm。保护区总面积6 264.00 hm2。通过对调查数据的处理,在松山国家级自然保护区一共发现胡桃楸斑块65块,胡桃楸林总计为396.47 hm2。主要分布在大庄科村、兰角沟等地区的沟谷内阴坡一侧,海拔高度为700~1 500 m,以天然纯林为主,萌生情况普遍,少量伴生黑枣Diospyros lotus,榆树Ulmus pumila和大叶白蜡Fraxinus rhynchophylla等。林龄组成以中龄林为主,成熟林和近熟林次之,幼龄林最少。
2 研究方法
2.1 样地设置
采用典型性样方法[13],于2015年7月至8月,在兰角沟、大庄科村附近、公路两侧主要胡桃楸林分布区选取立地条件相近、林龄(以40年生为林龄期限,用生长锥取得树龄数据)不同的典型林分设置20 m×20 m的乔木样方24个(图1),并在每个乔木样方四角及中心位置各做5 m×5 m灌木样方1个和1 m×1 m草本样方1个。样方内记录:①生态因子,如海拔、坡度、坡向、坡位和土壤情况;②乔木树种的名称、胸径、高度以及整个群落的郁闭度;③灌木和草本植物的名称、数量、高度及盖度[14]。

图1 胡桃楸林样方分布图Figure 1 Distribution pattern of Juglans mandshurica forests
2.2 数据统计分析
根据样方内植物个体高度、盖度、密度以及频度,计算灌木和草本物种的重要值。公式为:灌木植物重要值=(相对密度+相对高度+相对盖度)/3;草本植物重要值=(相对频度+相对密度+相对高度+相对盖度)/4[5]; 相对频度=该种的频度/所有种的频度之和,相对密度=该种的个体数/所有种的个体数总和,相对高度=该种的平均高度/所有种的平均高度总和,相对盖度=该种的盖度/全部种的盖度总和[5,15-17]。总结前人研究成果,根据多样性指数对反映群落物种多样性状况的实际应用效果, 本研究选取了以下 4种多样性指数[5,15-17]。 Simpson指数Shannon-Wiener指数Pielou 均匀度指数(J): J=H/lnS。 Margalef丰富度指数(M): M=(S-1)/lnN。 其中:S为样方内物种数;N为样方内所有物种个体数总和;Pi为种i的相对重要值,等于种i的重要值占样方中所有种的重要值之和的比例。
利用SPSS 22.0软件进行均值分析,并对不同林龄胡桃楸林植物多样性指数进行单因素方差分析;利用SigmaPlot 12.0对结果进行图形化处理;利用GS+9.0软件对群落物种多样性空间分布进行半变异函数与方差模型分析。
3 结果分析
3.1 不同林龄胡桃楸林下植物物种组成分析
调查记录胡桃楸林下植物43科78种,以菊科Asteraceae,蔷薇科Rosaceae,豆科Leguminosae,毛茛科Ranunculaceae和百合科Liliaceae等为主,其中木本植物18科26种,草本植物25科52种。同时还对胡桃楸林内灌木层和草本层植物的重要值进行了统计(表1与表2)。
灌木层中,葎叶蛇葡萄Ampelopsis humulifolia,小叶鼠李Rhamnus parvifolia,红花锦鸡儿Caragana rosea和山楂叶悬钩子Rubus crataegifolius在不同林龄林下都有分布。红花锦鸡儿与小叶鼠李在4种林龄林下都有较大优势,而山楂叶悬钩子和葎叶蛇葡萄则存在差异。荆条Vitex negundo var.heterophylla和山杏Armeniaca sibirica分布在幼龄林及中龄林中,毛叶丁香Syringa pubescens,小花溲疏Deutzia parviflora,暴马丁香Syringa reticulata var.mandshurica和土庄绣线菊Spiraea pubescens等在近熟林与成熟林中分布,细叶小檗Berberis poiretii和大花溲疏Deutzia grandiflora仅在成熟林中可见。整体而言,幼龄林及中龄林的灌木种类较少,随着林龄的增加,灌木层植物种类逐渐增加,存在明显的变化趋势。保护区内胡桃楸林下灌木群落以小花溲疏、毛叶丁香、小叶鼠李、红花锦鸡儿等为主要优势种。
草本层中鸭跖草Commelina communis在不同林龄下均有分布,但重要值存在差异。不同林龄的胡桃楸林下草本群落组成差异显著,随着林龄增加,大油芒Spodiopogon sibiricus,披针叶薹草Carex lancifolia,蝙蝠葛Menispermum dauricum等逐渐被北重楼Paris verticillata,鼠掌老鹳草Geranium sibiricum,蓝萼香茶菜Rabdosia japonica等取代,形成较稳定的顶级群落。优势种上,幼龄林和中龄林主要是披针叶薹草,大油芒,草乌Aconitum carmichaeli和龙芽草Agrimonia pilosa等,近熟林和成熟林则以蓝萼香茶菜和白屈菜Chelidonium majus等为主。整体而言,随着林龄的增加,草本层植物种类减少的趋势明显。
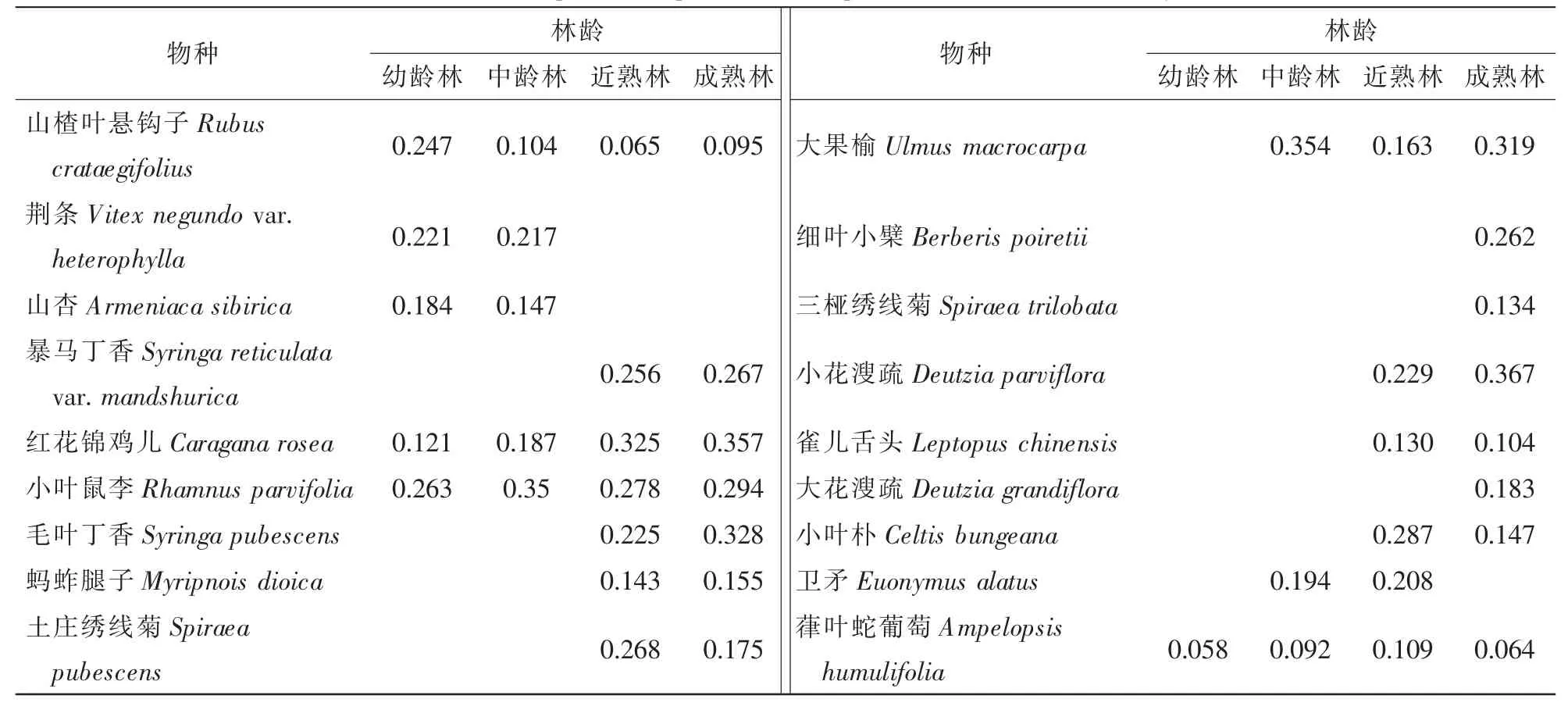
表1 灌木层主要组成物种及其重要值Table 1 Main species composition and important values of the shrub layer
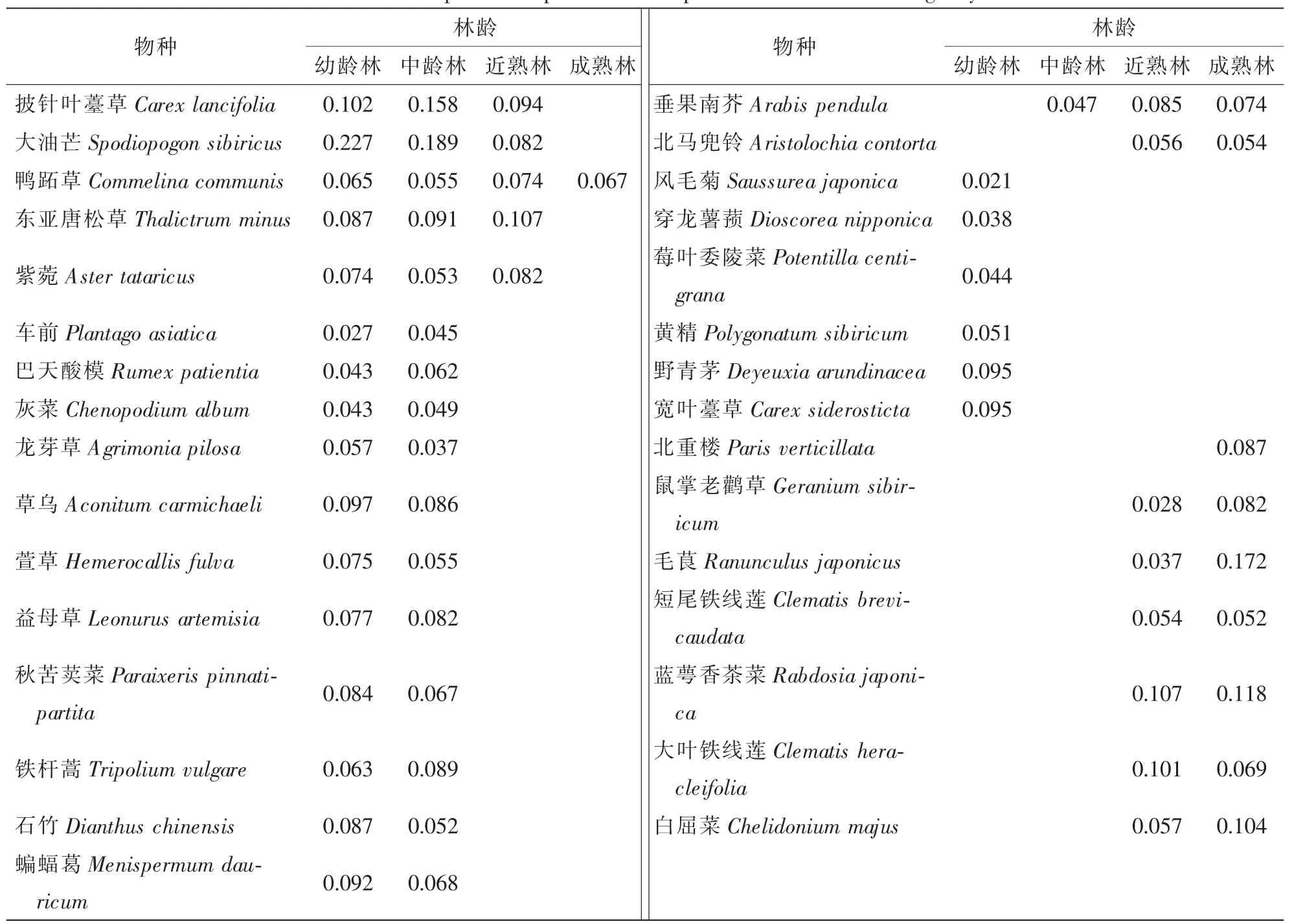
表2 草本层主要组成物种及其重要值Table 2 Main species composition and important values of the herbage layer
3.2 不同林龄胡桃楸林下植物多样性差异分析
3.2.1 不同林龄胡桃楸林下灌木层物种多样性分析 通过单因素方差分析,对不同林龄胡桃楸林下灌木层物种多样性进行差异显著性分析。结果显示:不同演替阶段的灌木层Simpson指数存在显著差异(P<0.05),随着林龄增加,灌木层物种变化明显,指数值不断增加(图2和表3);而Shannon-Wiener多样性指数和均匀度指数分别呈极显著(P<0.001)和显著(P<0.05)的差异,随着林龄的增加而逐渐增加。这表明,随着演替阶段的深入,灌木的物种多样性表现出不断递增的规律。
3.2.2 不同林龄胡桃楸林下草本层物种多样性分析 单因素方差分析显示,不同林龄下草本层的Simpson多样性指数存在显著差异(P<0.05),并随林龄的增加,呈现逐渐降低的规律(图3与表3);Shannon-Wiener多样性指数差异更为显著(P<0.001),随着林龄增加,多样性指数下降的趋势更加明显;不同演替阶段的草本层均匀度指数也存在显著差异(P<0.001),但变化趋势是上升的。这表明,随着演替阶段的深入,草本植物有逐渐退出生态系统的趋势,最后形成一个稳定的顶级群落,这种变化规律与Bohanek 等人提出的观点是一致的[17]。
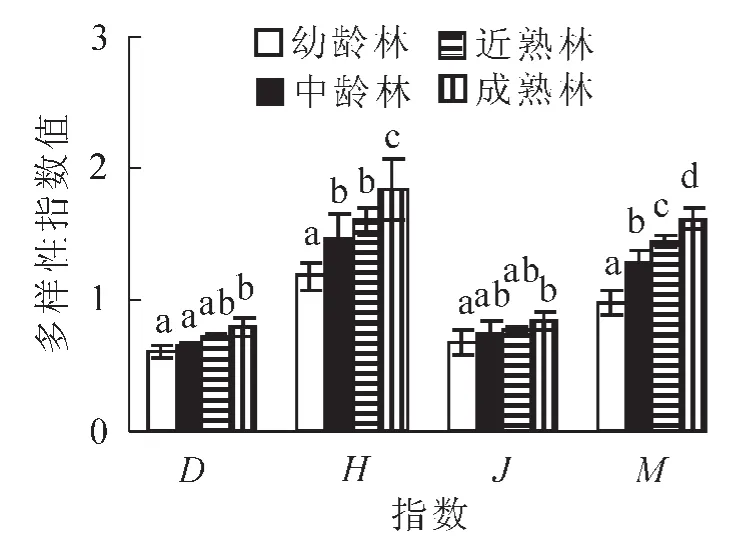
图2 不同林龄胡桃楸林下灌木层物种多样性指数Figure 2 Species diversity index of shrub layers under forest at different ages of manchurian walnut plantations
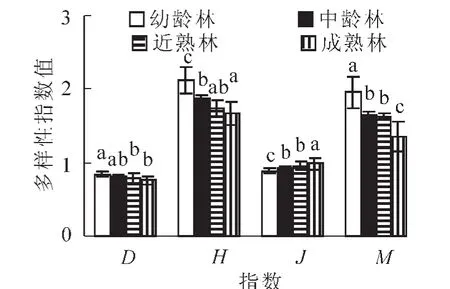
图3 不同林龄胡桃楸林下草本层物种多样性指数Figure 3 Species diversity index of herb layers under forest at different ages of manchurian walnut plantations
3.2.3 不同林龄胡桃楸林下物种丰富度差异分析 通过单因素方差分析,灌木层的物种丰富度差异极显著(P<0.001),与幼龄林相比,中龄林灌木层物种丰富度增加了20%以上,近熟林比中龄林又增加了近43%(图4),最后在成熟林阶段达到了最高值,但与近熟林相比,成熟林的物种丰富度增加比例不到9%。这表明:从胡桃楸群落演替早期到达到稳定状态的期间,物种丰富度先快速增加,到中期增速达到最快,而到了后期阶段,随着群落逐渐稳定,植物物种的替换速率也随之下降。草本层的变化规律与灌木层呈现完全相反的趋势。随着胡桃楸林龄的增加,林下草本层植物的种类逐渐减少(图4)。不同林龄胡桃楸林下草本层物种丰富度存在极显著差异(P<0.001),在演替初期,大量的草本植物出现,形成早期的先锋群落,但随着林龄增加,群落中草本植物不断退出群落,尽管有部分新的植物进入,但在整体上,最后的稳定状态的顶级群落与早期相比,草本层物种丰富度下降非常明显。
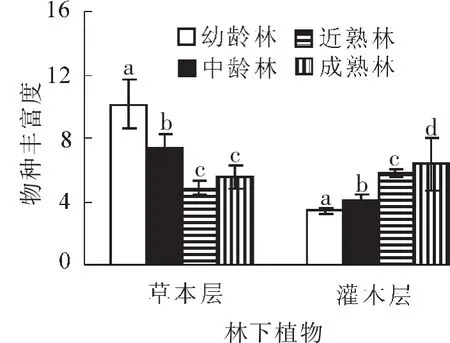
图4 胡桃楸林下植物物种丰富度指数Figure 4 Species richness index of undergrowth under forest of manchurian walnut plantations
3.3 胡桃楸林下植物多样性空间异质性分析
从表4物种多样性半变异函数模型及方差分析结果可知,松山胡桃楸林4种多样性指数中,均匀度指数和Simpson多样性指数为球面模型,Shannon-Wiener多样性指数和丰富度指数则更符合指数模型,半变异函数模型能够很好地反映物种多样性指数的空间结构特性。在分析模型中块金值表征由于取样误差和小于取样尺度上随机引起的空间异质性,基台值越大,总的空间异质性越高;块金与基台的比值表示由系统变量引起的空间相关性,比值小于0.25表示空间相关性强烈,0.25~0.75表示中等的空间相关性,大于0.75表示空间相关性较弱[18];4种多样性指数比值都为0.25~0.75,因此研究区的物种多样性为中等的空间相关性。
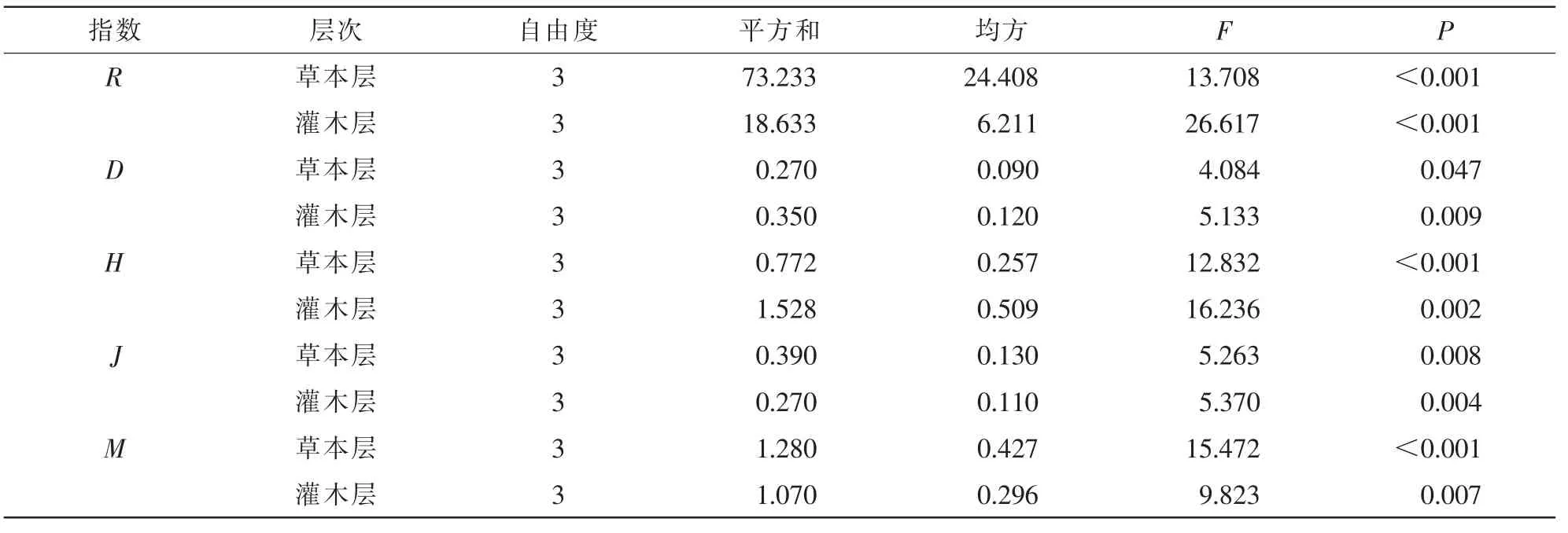
表3 林龄对不同层物种多样性的影响Table 3 One-way ANOVA for shrub and herbage diversity index affect by forest age
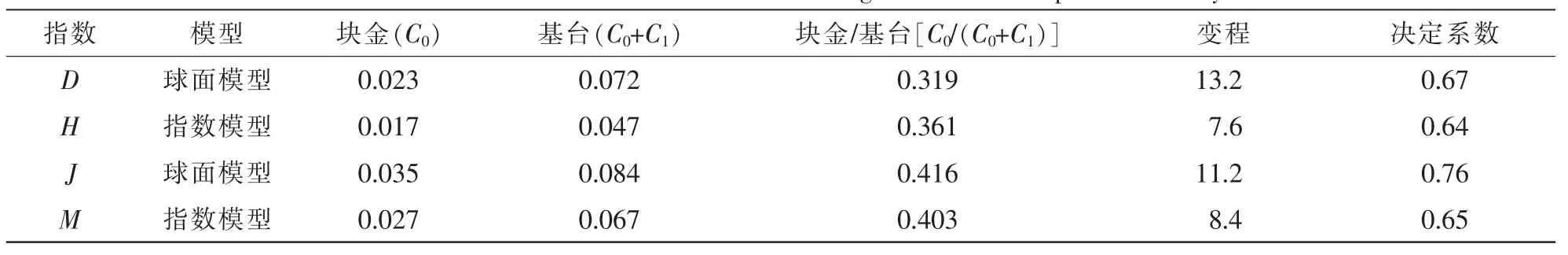
表4 物种多样性半变异函数模型及其相关参数Table 4 Parameters and test values of theoretical variogram models for species diversity indexes
同时也说明:在大于研究尺度,小于变程的范围内结构性因子所占比例为0.584~0.681,随机性因子所占比例为0.319~0.416,说明尽管结构性因子仍然占主导地位,但诸如人为砍伐、旅游开发等人为活动引起的空间异质性起着非常重要的作用,人为活动的干扰对胡桃楸植物多样性有着显著影响。
4 讨论与结论
4.1 不同林龄胡桃楸林下植物物种组成特征与变化趋势
灌木层中红花锦鸡儿、小叶鼠李等在各林分都属于优势度较大的树种,草本层中大油芒、披针叶薹草等属于优势植物。随着林龄的增加,草本植物物种数逐渐下降,可能是由于林下生存空间及养分的竞争加剧,竞争力差的植物逐渐淘汰出局[19-21]。葎叶蛇葡萄、东亚唐松草、鸭跖草等几乎在胡桃楸各个林龄阶段都有出现,说明这些物种非常适应胡桃楸林下的环境[22]。由于不同物种对特定的林下生境的适应能力各不相同,群落中的优势种类随演替阶段不断发生变化。整体而言,演替早期胡桃楸草本群落主要以披针叶薹草、大油芒、草乌、龙芽草等为优势种,中后期则以蓝萼香茶菜、白屈菜等为优势种;灌木群落以小花溲疏、毛叶丁香、小叶鼠李、红花锦鸡儿等为优势种。
4.2 不同林龄胡桃楸林下植物多样性
杨立学等[10]认为:胡桃楸林龄可以对植物群落的完整性、抗逆性及演替的进程等状况进行真实反映。随着林龄的增加,胡桃楸林下灌木层物种多样性指数呈现连续增加的趋势,而草本层的Simpson指数、Shannon-Wiener指数、物种丰富度指数呈现持续递减的规律,均匀度指数则与之相反,表现为连续递增趋势。这表明,随着胡桃楸的干扰逐渐增强,林下灌木层和草本层的响应并不一致,适应能力较强的灌木在群落中稳定下来,适应能力较弱的草本植物则有退出群落的趋势;而幼龄林林下植被的均匀度指数明显不如成熟林高,可能是因为到了成熟林阶段,林分结构逐渐稳定下来,外部环境相对比较固定,对林下植物生长与分布的限制作用趋于减弱。
胡桃楸不同演替阶段植物多样性之所以存在显著差异,主要源于在以胡桃楸为优势树种的林分中,随着林龄的增加,胡桃楸在群落中的优势地位增强,造成林下植物对有限的空间和资源的竞争更为激烈,只有适应能力更强的个体才得以生存,部分植物逐渐趋于淘汰、消亡,最终完成胡桃楸林下群落演替过程。
4.3 建议
自然状态下,胡桃楸的化感作用使其群落保持优势,但林龄的变化会直接导致林地中胡桃醌的含量变化,最终影响胡桃楸林群落结构的维持[11,23]。松山胡桃楸林多分布于沟谷、道路两旁的区域,这些地区是保护区发展生态旅游的重点区域,景区设施与道路的建设,加上村民对胡桃楸种子的采集,造成很多地方胡桃楸天然实生更新困难。人为活动对胡桃楸林植物多样性产生了显著影响,胡桃楸的自然更新与维持受到了威胁,因此,科学合理的规划景区建设及道路的开发,有效地对当地居民的种子采集等活动进行管理,就显得十分重要。
[1] LEVINEL J M,LAMBERS J H R.The importance of niches for maintenance of species diversity [J].Nature,2009,461(7261):254 - 257.
[2] THIBAUT L M, CONNOLLY S R.Understanding diversity-stability relationships:towards a unified model of portfolio effects[J].Ecol Lett,2013,16(2):140 - 150.
[3] BURKE A.Classification and ordination of plant communities of the Naukluft mountains,Namibia[J].J Veg Sci,2001,12(1):53 - 60.
[4] 张雪妮,吕光辉,王庭权,等.荒漠区垂直河岸带植物多样性格局及其成因[J].生态学报,2015,35(18):5966-5974.ZHANG Xueni, LÜ Guanghui, WANG Tingquan,et al.Patterns and mechanisms influencing plant diversity in an arid desert region riparian zone of the Aqikesu River, Xinjiang, China [J].Acta Ecol Sin, 2015, 35(18):5966 -5974.
[5] 张锦春,王继和,赵明,等.库姆塔格沙漠南缘荒漠植物群落多样性分析[J].植物生态学报,2006,30(3):375-382.ZHANG Jinchun, WANG Jihe, ZHAO Ming, et al.Plant community and species diversity in the south fringe of Kumtag Desert[J].J Plant Ecol, 2006,30(3): 375 - 382.
[6] 袁蕾,周华荣,宗召磊,等.乌鲁木齐地区典型灌木群落结构特征及其多样性研究[J].西北植物学报,2014,34(3): 595 - 603.YUAN Lei, ZHOU Huarong,ZONG Zhaolei,et al.Structural characteristics and diversity of typical shrub plant community in the Urumqi Region [J].Acta Bot Boreal-Occident Sin, 2014, 34(3): 595 - 603.
[7] 孟莹莹,周莉,周旺明,等.长白山风倒区植被恢复26年后物种多样性变化特征[J].生态学报,2015,35(1):142 - 149.MENG Yingying, ZHOU Li, ZHOU Wangming, et al.Characteristics of plant species diversity in a windthrow area on Changbai Mountain after 26 years of natural recovery [J].Acta Ecol Sin, 2015, 35(1):142 - 149.
[8] GASTON K J.Global patterns in biodiversity [J].Nature,2000,405(6783):220 - 227.
[9] 崔宁洁,陈小红,刘洋,等.不同林龄马尾松人工林林下灌木和草本多样性[J].生态学报,2014,34(15):4313-4323.CUI Ningjie, CHEN Xiaohong, LIU Yang, et al.Shrub and herb diversity at different ages of Pinus massoniana plantation [J].Acta Ecol Sin, 2014, 34(15):4313 - 4323.
[10] 杨立学,孙跃志.不同林龄核桃楸林下植物多样性的差异[J].生态学杂志,2013,32(4):807-812.YANG Lixue,SUN Yuezhi.Biodiversity of understory vegetation in different-aged Manchurian walnut plantations[J].Chin J Ecol, 2013, 32(4):807 - 812.
[11] 蒋万杰,杜连海,刘桂林,等.松山自然保护区的生物多样性及其威胁因素研究[J].林业建设,2008(4):6-9.JIANG Wanjie, DU Lianhai, LIU Guilin, et al.Biodiversity and threatening factors in Songshan National Nature Reserve [S].For Construct, 2008(4):6 - 9.
[12] 苏日古嘎,张金屯,王永霞.北京松山自然保护区森林群落物种多样性及其神经网络预测[J].生态学报,2013, 33(11):3394 - 3403.Suriguga, ZHANG Jintun, WANG Yongxia.Species diversity of forest communities and its forecasting by neural network in the Songshan National Nature Reserve, Beijing [J].Acta Ecol Sin, 2013, 33(11):3394 - 3403.
[13] 宋永昌.植被生态学[M].上海:华东师范大学出版社,2001:550-579.
[14] 邢韶华,袁秀,林大影,等.北京雾灵山自然保护区胡桃楸群落结构[J].浙江林学院学报,2006,23(3):290-296.XING Shaohua, YUAN Xiu, LIN Daying, et al.Juglans mandshurica community structure in Nature Reserve of Mount Wuling [J].J Zhejiang For Coll, 2006, 23(3): 290 - 296.
[15] 吴征镒.中国植被[M].北京:科学出版社,1980.
[16] JENNINGS M D,FABER-LANGENDOEN D,LOUCKS O L,et al.Standards for associations and alliances of the US National Vegetation Classification [J].Ecol Monogr,2009,79(2):173 - 199.
[17] 张金屯.数量生态学[M].北京:科学出版社,2003.
[18] BOHANEK J R,GRONINGER J W.Productivity of European black alder (Alnus glutinosa) interplanted with black walnut(Juglans nigra) in Illinois,USA [J].Agrofor Syst,2005,64(2):99 - 106.
[19] 马斌,周志宇,张莉丽,等.阿拉善左旗植物物种多样性空间分布特征[J].生态学报,2008,28(12):6099-6106.MA Bin, ZHOU Zhiyu, ZHANG Lili, et al.The spatial distribution characteristics of plant diversity in Alex Left Banner[J].Acta Ecol Sin, 2008, 28(12): 6099 - 6106.
[20] 赵成章,石福习,董小刚,等.祁连山北坡退化林地植被群落的自然恢复过程及土壤特征变化[J].生态学报,2011, 31(1): 115 - 122.ZHAO Chengzhang, SHI Fuxi,DONG Xiaogang,et al.Dynamics of vegetation structure and soil properties in the natural restoration process of degraded woodland on the northern slope of Qilian Mountains, northwestern China [J].Acta Ecol Sin, 2011, 31(1): 115 - 122.
[21] 冶民生,关文彬,吴斌,等.岷江干旱河谷植物群落的复杂性[J].生态学报,2006,26(10):3159-3165.YE Minsheng,GUAN Wenbin,WU Bin,et al.Plant community complexity in the arid valley of Minjiang River [J].Acta Ecol Sin,2006,26(10):3159 - 3165.
[22] 吴彦,刘庆,何海,等.亚高山针叶林人工恢复过程中物种多样性变化[J].应用生态学报,2004,15(8):1301-1306.WU Yan,LIU Qing,HE Hai,et al.Dynamics of species diversity in artificial restoration process of subalpine coniferous forest[J].Chin J Appl Ecol,2004,15(8):1301 - 1306.
[23] PONDER F J,TADROS S H.Juglone concentration in soil beneath black walnut interplanted with nitrogen-fixing species [J].J Chem Ecol,1985,11(7):937 - 942.
Undergrowth diversity at different ages of Juglans mandshurica forests in the Songshan National Nature Reserve,Beijing
ZHENG Changlong,WANG Jianming,LI Jingwen
(College of Forestry,Beijing Forestry University,Beijing 100083,China)
S718.5
A
2095-0756(2017)05-0825-08
2016-09-26;
2016-10-22
国家林业局第2次全国重点保护野生植物调查项目(林护发〔2012〕87号)
郑昌龙,从事生态学研究。E-mail:574441437@qq.com。通信作者:李景文,教授,博士,从事生态学研究。E-mail:lijingwen@bjfu.edu.cn
猜你喜欢
江西农业大学学报(2022年1期)2022-02-04 11:51:42
江西农业大学学报(2021年2期)2021-05-16 02:18:24
防护林科技(2020年10期)2020-12-22 11:46:42
疯狂英语·初中天地(2019年10期)2019-10-23 05:25:20
学与玩(2017年4期)2017-02-16 07:05:44
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:50
防护林科技(2016年5期)2016-09-05 01:19:43
小学教学研究(2016年36期)2016-05-17 05:51:58
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
