
肝细胞肝癌肿瘤微环境中功能耗竭CD8+T细胞脂代谢特征的观察
2017-09-22 08:31卓乐盈刘芳铭张苗苗刘卫仁武多娇
中国临床医学 2017年4期
沈 琦, 卓乐盈, 刘芳铭, 张苗苗, 王 晗, 刘卫仁, 武多娇*
1.复旦大学附属中山医院实验研究中心,上海 2000322.温州医科大学,温州 325035
·论著·
肝细胞肝癌肿瘤微环境中功能耗竭CD8+T细胞脂代谢特征的观察
沈 琦1, 卓乐盈2, 刘芳铭1, 张苗苗1, 王 晗1, 刘卫仁1, 武多娇1*
1.复旦大学附属中山医院实验研究中心,上海 2000322.温州医科大学,温州 325035
目的: 探讨肝细胞肝癌(hepatocellular carcinoma,HCC)肿瘤微环境中CD8+T细胞耗竭与脂代谢的关系。方法: 利用流式细胞分析、实时细胞能量代谢检测仪、实时PCR等方法检测HCC和癌旁组织中CD8+T细胞脂代谢及功能相关蛋白或基因的表达;在此基础上分析其功能和脂代谢的关系。结果: HCC浸润的CD8+T细胞具有以下特征:功能耗竭(干扰素-γ减少、程序性死亡分子-1表达升高);多个脂肪酸合成酶表达下调,脂肪酸含量下降;线粒体结构、功能受损(P<0.05)。结论: HCC中浸润的功能耗竭CD8+T细胞具有脂肪酸代谢异常特征。
肝细胞肝癌;CD8+T细胞;代谢;功能耗竭;微环境
肿瘤微环境在肿瘤发生、发展过程中有至关重要的作用[1]。细胞毒性CD8+T淋巴细胞是肿瘤特异性适应性免疫反应的关键细胞,攻击表面上具有主要组织相容性Ⅰ类复合物的肿瘤相关抗原肽的肿瘤细胞。CD8+T细胞在与其肿瘤靶标相互作用后,可通过干扰素-γ(IFN-γ)依赖性机制杀伤肿瘤细胞。肿瘤微环境抑制因素的持续存在最终导致浸润CD8+T细胞丧失杀伤肿瘤的能力,成为耗竭性细胞,导致肿瘤的免疫逃逸[1]。逆转CD8+T细胞耗竭(T cell exhaustion),恢复其抗肿瘤活性是肿瘤免疫治疗的重要策略之一。CD8+T细胞耗竭的特征是具有免疫抑制作用的分子,包括程序性死亡分子-1(PD-1)、细胞毒性T淋巴细胞抗原4(CTLA-4)、淋巴细胞活化基因3(LAG-3)等在细胞上表达上调,具有抑制CD8+T细胞活化和效应的功能。因此,研究CD8+T细胞功能障碍的机制至关重要,也是下一步设计联合多靶点肿瘤免疫治疗策略的关键。
目前已有文献[2-3]提示,CD8+T细胞功能耗竭与其代谢异常密切相关,无论是在慢性淋巴细胞性脉络丛脑膜炎病毒(LCMV)感染或黑素瘤小鼠模型中,耗竭的CD8+T细胞过表达共抑制分子的同时都呈现出广泛的代谢和生物能量缺陷的特点。科学家在小鼠肉瘤模型中发现肿瘤细胞掠取T细胞营养,导致哺乳动物雷帕霉素靶蛋白(mTOR)活性和糖酵解能力减弱、效应因子IFN-γ分泌减少,促进了肿瘤进展[4]。结果提示肿瘤浸润的CD8+T细胞代谢异常与微环境中肿瘤细胞营养竞争导致组织中可获取的葡萄糖等营养物质减少有关。
前期研究[5]表明,Toll样受体9(TLR9)激活的浆细胞样树突状细胞发生代谢重组,脂肪酸氧化增强是该类细胞活化并分泌大量Ⅰ型干扰素的能量基础;过氧化物酶体增殖物激活受体α(PPARα)介导了这一分解代谢过程。另外有研究[6-7]发现,黑素瘤中效应性T细胞特异性缺失乙酰辅酶A羧化酶1(ACC1,脂肪酸合成的限速酶),其功能会受损,细胞数量也显著减少;而给予ACC1缺陷型细胞补充外源性脂肪酸,可以恢复其功能。
在肝炎、肝硬化等致病因素反复刺激下,肝细胞肝癌(hepatocellular carcinoma,HCC)被认为是典型的“微环境肿瘤”。根治性手术切除目前是HCC治疗首选。然而,由于术后高复发转移率等原因,近十年来HCC生存率并未得到明显改善。本研究从代谢角度探讨HCC微环境中CD8+T细胞耗竭的代谢特征,为后续研究奠定基础。
1 材料与方法
1.1 标本及组织病理 选取复旦大学附属中山医院肝外科2016年6月至2016年10月经手术切除的HCC标本20例,另取癌旁3 cm以上的肝组织标本20例作为对照组。HCC标本组中,10例进行免疫组织化学染色,具体步骤如下:石蜡包埋的组织切成5 μm切片,进行免疫组织化学染色。抗人CD8抗体 (赛默飞公司,货号RB-9009-P0)孵育,苏木精复染。使用ImagePro Plus 软件(美国Media Cybernetics公司,V6.0)在高倍镜视野下,每张切片随机选择5个视野,比较癌和癌旁CD8抗体阳性的细胞数目。本研究获得医院伦理委员会批准,所有患者均知情同意并签署知情同意书。
1.2 单细胞悬液制备 20例HCC标本中有10例癌和癌旁组织分别制备单细胞悬液,步骤如下:在超净台中,将组织放入装有消化液的培养皿中,用眼科剪切成0.5~1.0 mm3。剪碎的组织放入烧瓶中,加入5~10 mL Ⅳ型胶原酶消化液,以及青霉素及链霉素双抗100 U/mL,置于37℃水浴磁力搅拌器中振荡消化,连续观察0.5~1 h,直至组织碎块弥散开为止。消化组织液用200目不锈钢滤网过滤。收集上述液体,1 800 r/min离心5 min,收集细胞沉淀,用Hanks液洗涤2次。
1.3 细胞实时能量代谢特征检测 分选癌旁和肿瘤组织内CD8+T细胞(CD3+CD8+TCRβ+CD45RO+CD62L-),简述步骤如下:(1)如上述获取细胞悬液;(2)进行抗体染色:CD3(FITC,克隆号HIT3a),CD8+(PE,克隆号HIT8a),TCRβ+(PE-cy7,克隆号3C10),CD45RO+(Percp,克隆号UCHL1),CD62L-(APC-cy7,克隆号DREG-56),上述流式抗体都购自Biolegend公司;(3)抗体染色4℃孵育30 min后使用流式缓冲液清洗,缓冲液为含2%胎牛血清(Gibco公司)的磷酸盐缓冲液(PBS);(4)清洗后细胞移入流式上样管,调整管中的细胞密度至(7~9)×106/mL,过滤细胞,避光置冰上运至分选室;(5)上机检测并设定分选条件,选取85 μmol/L喷嘴,调整分选关键参数及设门,分选细胞后上机回测,检测纯度。
细胞实时能量检测仪(安捷伦公司,Seahorse XF96)检测细胞有氧呼吸O2消耗速率(OCR)、细胞外酸化率(ECAR)、细胞储备呼吸能力(SRC)等指标;其中,OCR代表细胞线粒体氧化磷酸化水平;ECAR代表糖酵解水平;SRC代表细胞在应激及压力状态下生存的能力。具体方法如下:将T细胞 (1.5×105细胞/孔)移入特殊的96孔检测培养板中,置入XF96细胞能量代谢实时测定仪检测。在线粒体能量测试中,首先测定基线OCR,然后依次测量加入1 μmol/L寡霉素(oligomycin,ATP合成酶抑制剂)、1.5 μmol/L氟羰基氰化物苯腙(FCCP,离子载体)和100 nmol/L 鱼藤酮(rotenone,电子转运链抑制剂)及1 μmol/L抗霉素A(antimycin A,电子转运链抑制剂)后的OCR。上述试剂来自于线粒体压力测试盒内的配套试剂(安捷伦公司,货号103015-100)。
1.4 线粒体特征观察 单细胞悬液用Mitotracker Green(赛默飞公司,货号M7514)、Mitotracker Red(赛默飞公司,货号M22425)等探针染色后,流式细胞仪检测线粒体质量、膜电位等指标。
1.5 流式细胞仪检测 应用Bodipy探针(赛默飞公司,货号D3922)、葡萄糖转运蛋白Glut1抗体(R&D公司,货号MAB1418)标记染色等方法在流式细胞仪(BD公司, FACSAriaTMⅢ)上测量细胞脂质含量及葡萄糖转运蛋白表达水平;同样用抗体标记法检测癌旁和肿瘤组织内CD8+T细胞IFN-γ(Biolegend公司,克隆号GIR-94)、PD-1(Biolegend公司,克隆号EH12.2H7)表达水平。
细胞流式表面抗体染色步骤如下:(1)用PBS洗涤细胞,1 500 r/min离心5~10 min,弃上清,计数;(2)取适量含1%胎牛血清的流式缓冲液,按照比例加入抗体,4℃避光孵育细胞半小时;(3)1 500 r/min离心5~10 min,弃上清;(4)用1%胎牛血清的流式缓冲液洗涤细胞,1 500 r/min离心5~10 min,弃上清。重复洗涤后加入适量流式缓冲液混匀,上机检测。
1.6 Real-time PCR检测 流式细胞仪(BD公司, FACSAriaTMⅢ)分选HCC癌及癌旁的CD8+T细胞,提取核酸后进行Real-time PCR(罗氏公司,LightCycler○R96 Instrument,货号05815916001)检测脂肪酸合成相关的基因表达水平,具体方法如前所述[5]。
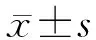
2 结 果
2.1 免疫组化染色结果 结果(图1)表明:癌组织平均高倍视野中CD8抗体表达阳性的细胞数低于癌旁组织[(3.4±0.8)vs(6.5±1.2),P<0.05 ]。
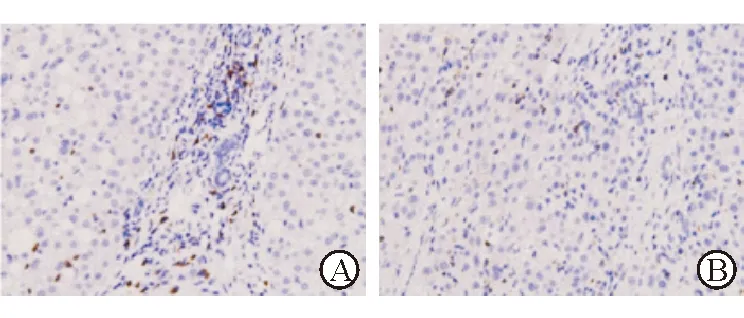
图1 HCC肿瘤组织及癌旁正常组织CD8抗体染色
A:癌旁正常组织;B:癌组织. Original magnification:×200
2.2 流式细胞仪检测 结果(图2)表明:肿瘤组织中CD8+T细胞数量少于癌旁组织(P=0.05),且效应分子IFN-γ表达减少[(13.2±3.6)%vs(5.4±2.1)%,P<0.05],共抑制分子PD-1、CD8+T细胞的表达增加[(9 800±1 660)vs(7 000±1 260),P<0.05],提示肿瘤组织内CD8+T细胞功能耗竭。

图2 CD8+T细胞效应分子及共抑制分子表达
A:CD8+T细胞数量;B:流式细胞仪检测CD8+T细胞IFN-γ表达;C,D:流式细胞仪检测CD8+T细胞PD-1表达.*P<0.05
2.3 代谢特征改变 结果(图3)表明:两组间Glut1表达无显著改变,说明肿瘤内浸润的CD8+T细胞从环境中摄取葡萄糖的活动未受到抑制;但肿瘤组织中的CD8+T细胞较癌旁细胞内脂质含量减少,表现为Bodipy脂质探针平均荧光强度减弱[(5 120±1 060)vs(2 200±1 260),P<0.05]。
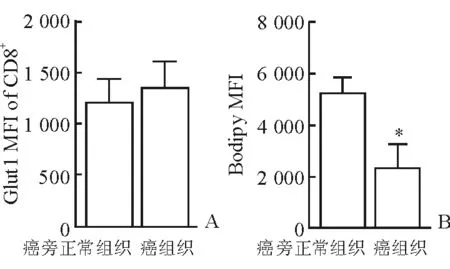
图3 CD8+T细胞代谢特征
A:葡萄糖转运蛋白Glut1表达;B:Bodipy染色显示脂质含量
2.4 线粒体特征改变 结果(图4)表明:肿瘤内浸润的CD8+T细胞线粒体电位下降[(8 100±960)vs(6 200±1 060),P<0.05)],伴随ATP产量下降(P<0.05)、氧化磷酸化下降(P<0.001)。
2.5 基因表达改变 Real-time PCR检测显示,肿瘤内浸润的CD8+T细胞ATP-柠檬酸裂解酶(ACLY)、脂肪酸合酶(FASN)基因表达下调(P<0.01,图5)。

图4 CD8+T细胞线粒体及实时能量代谢特征


图5 HCC肿瘤内浸润的CD8+T细胞脂肪酸合成相关酶表达
3 讨 论
脂肪酸代谢与细胞的功能密切相关。首先,脂肪酸和甾醇的合成是细胞增殖必需的原料。脂肪酸合成途径即细胞内脂质合成过程,可利用其他几种代谢途径的中间产物作为原料,例如糖酵解、三羧酸循环和磷酸戊糖途径。脂肪酸可以与糖酵解衍生的丙三醇缩合产生多种三酰基甘油和磷脂的组合,这是许多细胞结构的关键组分[8]。其次,合成的脂肪酸除了是细胞膜结构的关键组成部分,还可在线粒体或过氧化物酶体中进行氧化,产生许多具有重要生理功能的代谢中间产物,包括乙酰辅酶A、尼克酰胺腺嘌呤二核苷酸(NADH)和还原型黄素二核苷酸(FADH2)。另外,脂肪酸氧化产生大量单一棕榈酸酯分子(ATP,哺乳动物细胞中的主要脂肪酸),ATP的完全β-氧化具有产生超过100个ATP分子的潜力[9],是细胞供能的重要方式。
虽然已有部分文献[10-12]报道了脂肪酸代谢与免疫细胞的功能活化密切相关,但研究非常有限,且主要聚焦在脂肪酸氧化与病毒感染的动物模型上。本研究在人HCC标本中发现肿瘤浸润的CD8+T细胞比癌旁组织中CD8+T细胞数目减少且功能受损。这类T细胞在肿瘤杀伤功能异常的同时,表现出了代谢异常的特征,包括脂肪酸含量以及脂肪酸氧化和氧化磷酸化水平下降(氧耗率和储备呼吸能力SRC下降)、产能减少(ATP产生减少、线粒体膜电位下降)。胞内脂肪酸含量直接影响细胞脂肪酸氧化及氧化磷酸化水平,而脂肪酸氧化是免疫细胞生成ATP的关键方式;并且脂质是组成细胞膜结构的重要组成部分,脂肪酸从头合成减少会影响含膜结构细胞器的功能,如线粒体功能。线粒体功能可直接影响T细胞分化及免疫应答[13]。本研究发现,HCC中浸润的CD8+T细胞SRC减少,代表了细胞线粒体呼吸能力损伤[14]。另外,脂质是细胞增殖所必需的材料,脂质减少可显著妨碍细胞增殖和信号转导,这可部分解释肿瘤内浸润的CD8+T细胞数目比癌旁组织少。因此本研究中观察到HCC中CD8+T细胞脂肪酸含量下降可能是细胞产能及效应功能受抑制的重要原因之一。
其次,本研究发现HCC中耗竭的CD8+T细胞脂肪酸从头合成的酶ACLY、FASN表达下降。由于HCC组织样本中CD8+T细胞分选所得数量有限,目前还不能进行免疫印迹实验检测这些基因的蛋白水平。脂肪酸从头合成酶的表达主要由固醇调节元件结合蛋白(SREBPs)控制。SREBPs可能是HCC浸润的CD8+T细胞代谢重组的关键调控因子。SREBPs是3个基本螺旋-环-螺旋-亮氨酸拉链转录因子家族。SREBPs与其靶基因的启动子中的固醇调节元件(SRE)和一些E盒序列结合。目前已知蛋白激酶B/哺乳动物靶蛋白雷帕霉素复合物1(AKT/mTORC1)与SREBPs构成信号转导轴,诱导SREBPs核聚集,连接了致癌信号传导和脂质代谢。AKT/mTORC1-SREBP轴及其下游基因是否在HCC中耗竭的CD8+T细胞代谢中起关键调控作用目前尚不清楚,有待进一步探讨。
综上所述,本研究发现了HCC内浸润的功能耗竭的CD8+T细胞具有线粒体功能受损、脂肪酸代谢异常的特征。HCC中CD8+T细胞脂肪酸从头合成减少,可通过影响线粒体膜结构、限制了脂肪酸氧化等机制,导致线粒体结构和功能受损,最终削弱CD8+T细胞分泌IFN-γ等肿瘤杀伤功能。目前关于脂代谢与CD8+T细胞的研究还远不够深入,下一步可通过细胞及动物实验进一步验证脂代谢异常与T细胞功能异常的关系,继而探讨靶向干预CD8+T细胞脂代谢异常能否重激活T细胞的肿瘤杀伤作用。
[ 1 ] HANAHAN D, WEINBERG R A. Hallmarks of cancer: the next generation[J]. Cell, 2011, 144(5):646-674.
[ 2 ] WHERRY E J, HA S J, KAECH S M, et al. Molecular signature of CD8+T cell exhaustion during chronic viral infection[J]. Immunity, 2007, 27(4):670-684.
[ 3 ] SCHARPING N E, MENK A V, MORECI R S, et al. The tumor microenvironment represses T cell mitochondrial biogenesis to drive intratumoral T cell metabolic insufficiency and dysfunction[J]. Immunity, 2016, 45(2):374-388.
[ 4 ] CHANG C H, QIU J, O'SULLIVAN D, et al. Metabolic competition in the tumor microenvironment is a driver of cancer progression[J]. Cell, 2015, 162(6):1229-1241.
[ 5 ] WU D, SANIN D E, EVERTS B, et al. Type 1 interferons induce changes in core metabolism that are critical for immune function[J]. Immunity, 2016, 44(6):1325-1336.
[ 6 ] LEE J, WALSH M C, HOEHN K L, et al. Regulator of fatty acid metabolism, acetyl coenzyme a carboxylase 1, controls T cell immunity[J]. J Immunol, 2014, 192(7):3190-3199 .
[ 7 ] YANG W, BAI Y, XIONG Y, et al. Potentiating the antitumour response of CD8(+) T cells by modulating cholesterol metabolism[J]. Nature, 2016, 531(7596):651-655.
[ 8 ] MOON J S, LEE S, PARK M A, et al. UCP2-induced fatty acid synthase promotes NLRP3 inflammasome activation during sepsis[J]. J Clin Invest, 2015, 125(2):665-680.
[ 9 ] NOMURA M, LIU J, ROVIRA I I, et al. Fatty acid oxidation in macrophage polarization[J]. Nat Immunol, 2016, 17(3):216-217.
[10] WANG C, YOSEF N, GAUBLOMME J, et al. CD5L/AIM regulates lipid biosynthesis and restrains Th17 cell pathogenicity[J]. Cell, 2015,163(6):1413-1427.
[11] ANGELIN A, GIL-DE-GMEZ L, DAHIYA S, et al. Foxp3 reprograms T cell metabolism to function in low-glucose, high-lactate environments[J]. Cell Metab, 2017,25(6):1282-1293, e7.
[12] PAN Y, TIAN T, PARK C O, et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism[J]. Nature, 2017, 543(7644):252-256.
[13] BUCK M D, O′SULLIVAN D, KLEIN GELTINK R I, et al. Mitochondrial dynamics controls T Cell fate through metabolic programming[J]. Cell, 2016, 166(1):63-76.
[14] RÖHRIG F, SCHULZE A. The multifaceted roles of fatty acid synthesis in cancer[J]. Nat Rev Cancer, 2016, 16(11):732-749.
[本文编辑] 叶 婷, 贾泽军
Lipid metabolic characteristics of functionally exhausted CD8+T cells in tumor microenvironment of hepatocellular carcinoma
SHEN Qi1, ZHUO Le-ying2, LIU Fang-ming1, ZHANG Miao-miao1, WANG Han1, LIU Wei-ren1, WU Duo-jiao1*
1. Department of Experimental Research Center, Zhongshan Hospital, Fudan University, Shanghai 200032, China2. Wenzhou Medical University, Wenzhou 325035, Zhejiang, China
Objective: To explore the lipid metabolic characteristics of functionally exhausted CD8+T cells in tumor microenvironment of hepatocellular carcinoma (HCC).Methods: By using flow cytometry analysis and real-time PCR to detect the functions and lipid metabolism-related proteins and genes expression of CD8+T cells in HCC and peri-tumor tissues, then the relationship between function and lipid metabolism was analyzed.Results: The infiltrated CD8+T cells in HCC have the following characteristics: functional exhaustion (decreased interferon-γ secretion and up regulated programmed death molecule-1 expression); down regulated expression of multiple fatty acid synthases, and the decreased content of intracellular fatty acids; impaired mitochondrial structure and function (P<0.05).Conclusions: Fatty acid metabolism abnormalities are the characteristics of exhausted CD8+T cell infiltrated in HCC.
hepatocellular carcinoma; CD8+T cells; metabolism; functional exhaustion; microenvironment
R 735.7
A
2017-07-22 [接受日期] 2017-08-19
上海市浦江人才计划(17PJD006),上海市卫计委科研项目(201740101). Supported by Pujiang Talent Plan of Shanghai (17PJD006) and Project of Shanghai Municipal Health Planning Commission (201740101).
沈 琦,技师. E-mail: shen.qi@zs-hospital.sh.cn
*通信作者(Corresponding author). Tel: 021-64040731, E-mail: wu.duojiao@zs-hospital.sh.cn
10.12025/j.issn.1008-6358.2017.20170621
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
化工管理(2022年13期)2022-12-02
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国卒中杂志(2021年7期)2021-11-29
中老年保健(2021年4期)2021-08-22
中国生殖健康(2020年7期)2021-01-18
中国生殖健康(2020年5期)2021-01-18
健康博览(2020年11期)2020-11-23
中华养生保健(2020年4期)2020-11-16
意林·全彩Color(2019年6期)2019-07-24
