
高植物蛋白质饲料中添加丁酸钠对大菱鲆幼鱼生长性能、营养物质表观消化率及肝脏抗氧化功能的影响
2017-09-16 05:59魏朝青周慧慧麦康森
动物营养学报 2017年9期
魏朝青 周慧慧 王 旋 麦康森 徐 玮 何 艮
(中国海洋大学水产学院,农业部水产动物营养与饲料重点实验室,青岛266003)
高植物蛋白质饲料中添加丁酸钠对大菱鲆幼鱼生长性能、营养物质表观消化率及肝脏抗氧化功能的影响
魏朝青 周慧慧 王 旋 麦康森 徐 玮 何 艮*
(中国海洋大学水产学院,农业部水产动物营养与饲料重点实验室,青岛266003)
本试验旨在研究高植物蛋白质饲料中添加丁酸钠对大菱鲆幼鱼生长性能、营养物质表观消化率及肝脏抗氧化功能的影响,探索提高水产饲料中植物蛋白质替代鱼粉比例的可行性方法。以鱼粉、豆粕、玉米蛋白粉、谷朊粉、花生粕和啤酒酵母为蛋白质源,鱼油、椰子油和大豆卵磷脂为脂肪源,小麦粉为糖源,配制5种等氮等能的试验饲料。其中,以含60%鱼粉的基础饲料为阳性对照组(FM组),以复合植物蛋白质源替代基础饲料中50%鱼粉的替代饲料为阴性对照组(CON组),以在替代饲料中分别添加0.15%(D1组)、0.30%(D2组)和0.60%(D3组)丁酸钠的饲料为试验组。用上述5种试验饲料分别饲喂初始体重为(13.00±0.01) g的大菱鲆幼鱼58 d,试验在室内养殖系统中进行,每种试验饲料投喂3个重复,每个重复30尾鱼。结果表明:1)随着丁酸钠添加水平的增加,大菱鲆幼鱼的增重率、特定生长率和饲料效率呈现先升高后降低的趋势,其中FM、D1和D2组增重率、饲料效率和特定生长率显著高于CON组(P<0.05),而各组间摄食率无显著差异(P>0.05);除D2组脏体比与FM组无显著差异(P>0.05)外,其他各组的脏体比均显著低于FM组(P<0.05);各组全鱼水分、粗蛋白质和粗脂肪含量无显著差异(P>0.05)。2)D1组干物质表观消化率显著高于CON组(P<0.05),与FM组无显著差异(P>0.05);D1组蛋白质表观消化率显著高于CON、D2和D3组(P<0.05),与FM组无显著差异(P>0.05);D2和D3组之间干物质和蛋白质表观消化率差异不显著(P>0.05)。3)D2组肝脏总抗氧化能力(T-AOC)和过氧化氢酶(CAT)活性显著高于CON组(P<0.05),肝脏丙二醛(MDA)含量显著低于CON组(P<0.05)。由此得出,在本试验条件下,高植物蛋白质饲料中添加0.15%的丁酸钠能够提高大菱鲆幼鱼的生长性能、营养物质表观消化率和肝脏抗氧化功能,过量添加丁酸钠会降低大菱鲆幼鱼的生长性能。
丁酸钠;大菱鲆幼鱼;生长性能;表观消化率;抗氧化能力
由于鱼粉资源的日益短缺,使用动物蛋白质或植物蛋白质等非鱼粉蛋白质替代饲料中的鱼粉已经成为维持水产养殖业可持续发展的必然途径[1]。然而,水产动物饲料中植物蛋白质的使用比例往往存在一定的限制,使用过高水平的植物蛋白质替代鱼粉会导致养殖鱼类生长性能下降、摄食减少、消化率降低[2-4],同时还会引起氧化应激反应[5]。其主要原因归结于与鱼粉相比,植物蛋白质源缺乏多种功能性营养物质,存在氨基酸不平衡、含有难消化的碳水化合物及抗营养因子等[6]。
大量研究表明,在水产饲料中添加植物蛋白质源中缺乏的功能性小分子生物活性物质对于维持鱼体的正常生长至关重要。研究表明,在全植物蛋白质饲料中添加5 g/kg牛磺酸能显著改善虹鳟的生长性能并促进其摄食[7],在高植物蛋白质饲料中添加胆固醇[8]、羟脯氨酸[9]等能促进大菱鲆的生长,饲料中外源添加核苷酸可以提高大西洋鲑的生长性能[10]。因此,加强对功能性营养物质的研究可能是改善水产饲料中非鱼粉蛋白质利用效率的途径之一。
丁酸钠目前主要作为替代抗生素类添加剂在饲料中应用,其在促生长、促消化和提高免疫功能方面的作用已有大量学者报道,是一种十分具有潜力的功能性添加剂之一[11]。研究发现,丁酸钠能提高草鱼的特定生长率[12],其有效成分丁酸具有维持肠黏膜上皮细胞正常状态[13]、促进小肠消化吸收的功能[14]。同时,丁酸钠还可以缓解机体氧化应激,提高鱼体的抗应激能力。关于复合植物蛋白质替代鱼粉后添加丁酸钠对大菱鲆生长性能、消化能力和免疫功能等的影响以及能否提高植物蛋白质的利用率尚未见报道。因此,本试验以大菱鲆幼鱼为研究对象,通过向复合植物蛋白质源替代鱼粉后的饲料中添加不同剂量的丁酸钠,探讨其对大菱鲆幼鱼生长性能、形体指标、营养物质表观消化率和抗氧化功能的影响,就饲料中添加丁酸钠能否提高植物蛋白质替代鱼粉比例问题加以说明,为丁酸钠在水产动物养殖中的综合利用提供科学依据。
1 材料与方法
1.1试验设计与试验饲料
以鱼粉、豆粕、玉米蛋白粉、谷朊粉、花生粕和啤酒酵母为蛋白质源,鱼油和椰子油和大豆卵磷脂为脂肪源,小麦粉为糖源,配制5种等氮等能的试验饲料,以上饲料原料均购自青岛七好生物科技有限公司。其中,以含60%鱼粉的基础饲料为阳性对照组(FM组),以复合植物蛋白质源替代基础饲料中50%鱼粉的替代饲料为阴性对照组(CON组),以在替代饲料中分别添加0.15%(D1组)、0.30%(D2组)和0.60%(D3组)丁酸钠(购自上海涵乐生物科技有限公司,有效成分含量≥98.5%)的饲料为试验组。分别添加DL-蛋氨酸和赖氨酸使各组饲料中此2种必需氨基酸含量达到FM组的水平,同时在饲料中添加0.1%三氧化二钇(Y2O3)作为测定营养物质表观消化率的指示剂。饲料原料经粉碎后过60目筛,微量成分采取逐级扩大法混合均匀后,加入鱼油等脂肪源充分混合,最后加水和氯化胆碱混匀,经F-26双螺杆挤条制粒机制成硬颗粒饲料。饲料经45 ℃烘箱干燥12 h后置于-20 ℃冰柜中保存备用。试验饲料组成及营养水平见表1。
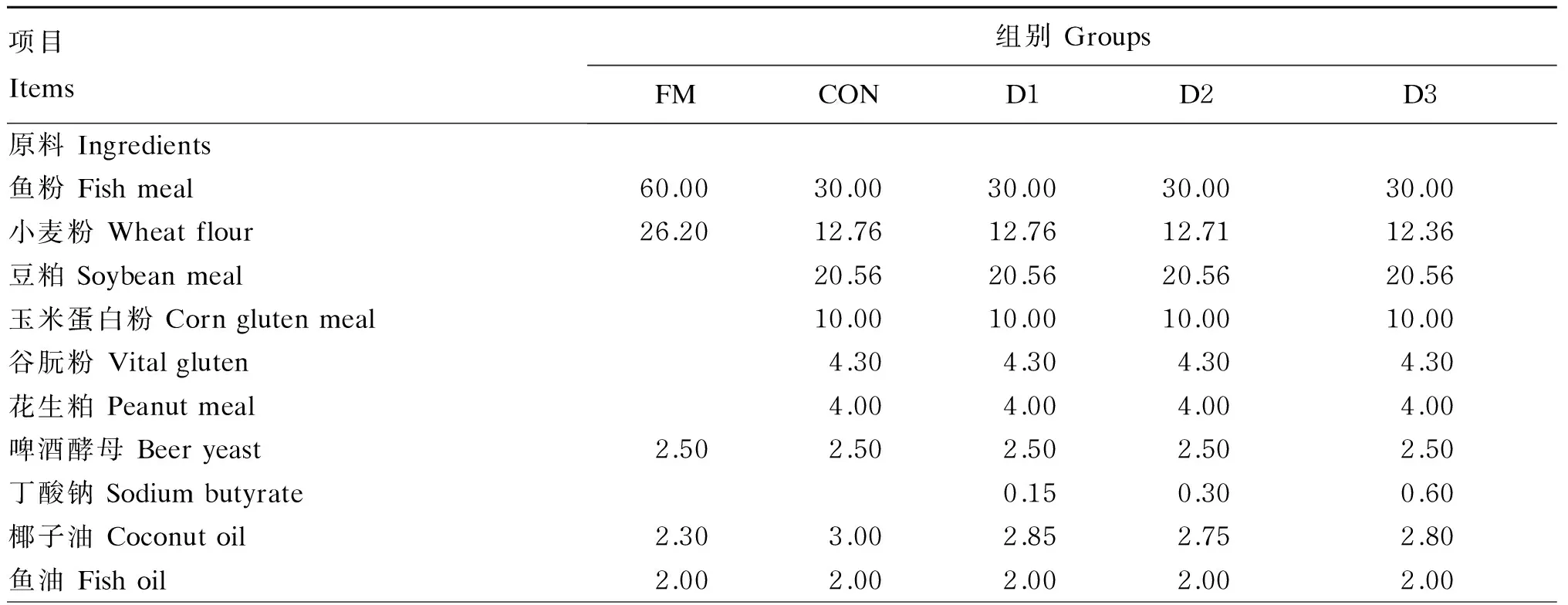
表1 试验饲料组成及营养水平(干物质基础)

续表1项目Items组别GroupsFMCOND1D2D3维生素预混料Vitaminpremix1)2.002.002.002.002.00矿物质预混料Mineralpremix2)1.001.001.001.001.00复合诱食剂Compositeattranct3)1.001.001.001.001.00牛磺酸Taurine1.001.001.001.00褐藻酸钠Sodiumalginate1.001.001.001.00DL-蛋氨酸DL-Met0.250.250.250.25赖氨酸Lys0.950.950.950.95大豆卵磷脂Soybeanlecithin2.502.502.502.502.50氯化胆碱Cholinechloride0.250.250.250.250.25磷酸二氢钙Ca(H2PO4)20.400.400.400.40植酸酶Phytase0.200.200.200.20三氧化二钇Y2O30.100.100.100.100.10丙酸钙Calciumpropionate0.100.100.100.100.10乙氧基喹啉Ethoxyquin0.050.050.050.050.05一水硫酸亚铁FeSO4·H2O0.050.050.050.05一水硫酸锌ZnSO4·H2O0.030.030.030.03合计Total100.00100.00100.00100.00100.00营养水平Nutrientlevels4)粗蛋白质Crudeprotein51.6852.7052.0751.2751.11粗脂肪Crudelipid10.2310.5510.6111.4111.33总能Grossenergy/(MJ/kg)20.1820.6220.5820.6220.65
1)每千克维生素预混料含有Contained the following per kg of vitamin premix:VA (500 000 IU/g) 32 mg,VB1(98%) 25 mg,VB2(80%) 45 mg,VB6(99%) 20 mg,VB12(1%) 10 mg,VD3(500 000 IU/g) 5 mg,VE (50%) 240 mg,VK (51%) 10 mg,泛酸钙 calcium pantothenate (98%) 60 mg,烟酸 nicotinic acid (99%) 200 mg,叶酸 folic acid (98%) 20 mg,生物素 biotin (2%) 60 mg,肌醇 inositol (98%) 800 mg,维生素C磷酸酯 VC phosphate (35%) 2 000 mg,微晶纤维素 microcrystalline cellulose 16 473 mg。
2)每千克矿物质预混料含有 Contained the following per kg of mineral premix:MgSO4·7H2O (15%) 1 200 mg,CuSO4·5H2O (25%) 10 mg,FeSO4·H2O (30%) 80 mg,ZnSO4·H2O (34.5%) 50 mg,MnSO4·H2O (31.8%) 45 mg,CoCl2·6H2O (1%) 50 mg,Na2SeO3(1%) 20 mg,碘酸钙 calcium iodate (1%) 60 mg,沸石粉 zeolite powder 8 485 mg。
3)复合诱食剂组成 Composite attractant composition:甜菜碱 lycine∶二甲基-丙酸噻亭 DMPT∶甘氨酸 glycine∶丙氨酸 alanine∶5-磷酸肌苷 inosine 5-phosphate=4∶2∶2∶1∶1。
4)营养水平均为实测值。Nutrient levels were all measured values.
1.2试验用鱼和养殖条件
试验用大菱鲆幼鱼购自烟台市莱州养殖场,养殖试验在青岛亿海丰水产品有限公司进行,正式试验前将大菱鲆幼鱼在养殖系统中暂养2周并驯化,期间投喂商业饲料以适应环境。驯化结束后选取大小均一、活力旺盛的大菱鲆幼鱼[初重(13.00±0.01) g]随机分为5组,每组3个重复,每个重复30尾鱼。试验鱼以重复为单位饲养于养殖桶(200 L)中。每种试验饲料随机投喂1组试验鱼。养殖试验在室内流水系统进行,养殖用水经水泵抽送到过滤池中,经沙滤后流到养殖桶内,流速保持一致。养殖期间,分别在每天07:00和19:00进行表观饱食投喂,摄食后进行残饵回收与统计,并换水以保证水质。
1.3样品采集
养殖试验结束后,饥饿24 h,用丁香酚(1∶10 000)麻醉所有试验鱼进行称重、计数。每桶随机取5尾鱼保存于-20 ℃冰箱以分析体组成;再从每桶中随机取4尾鱼,进行称重、量体长,解剖取内脏、肝脏并称重,以计算肝体比、脏体比,取肝脏置于离心管中放于液氮中,取样结束后置于-80 ℃冰箱中保存。以上样品每2尾鱼置于同一离心管内,每桶2管,每组共6个样品。每桶随机取4尾鱼,在摄食后4 h用挤压法[15]收集粪便样品。
1.4样品分析方法
1.4.1 生长性能指标
饲料原料、试验饲料、鱼体常规营养成分含量的测定均参照AOAC(1995)方法。其中,样品的水分和干物质含量用105 ℃烘箱烘干至恒重求得;粗蛋白质含量采用全自动凯氏定氮仪(TM-8400,瑞典FOSS公司)测定;粗脂肪含量采用索氏抽提仪(SOXTEC-8000,瑞典FOSS公司)测定;总能用氧弹热量仪(Parr1281,美国Parr公司)测定。
1.4.2 营养物质表观消化率
以添加在饲料中的0.1%Y2O3为指示剂,采用高频电感耦合等离子体发射光谱仪(VIATA-MPX-ICP,美国VARIAN公司)测定饲料和粪便中的钇含量,计算出饲料干物质和蛋白质的表观消化率。
1.4.3 肝脏抗氧化指标测定
准确称取一定重量的解冻的肝脏组织,按重量(g)∶体积(mL)=1∶9的比例加入0.9%的生理盐水,冰水浴条件下匀浆,制备成10%的组织匀浆液,4 ℃、3 000 r/min离心10 min,取上清液并根据不同指标的要求用生理盐水稀释成不同浓度进行测定。肝脏蛋白质含量采用考马斯亮蓝法测定,测定的肝脏抗氧化指标包括总抗氧化能力(total antioxidant capacity,T-AOC),过氧化氢酶(catalase,CAT)、超氧化物歧化酶(superoxide dismutase,SOD)活性以及丙二醛(malonyldialdehyde,MDA)含量,以上指标均采用南京建成生物工程研究所生产的试剂盒测定。
1.5计算公式
特定生长率(SGR,%/d)=100×(ln终末体重-ln初始体重)/养殖天数;增重率(WGR,%)=100×(终末体重-初始体重)/初始体重;摄食率(FI,%/d)=100×摄食饲料量/[(初始体重+终末体重)/2]/养殖天数;饲料效率(FE)=鱼体增重/摄食饲料量;肥满度(CF,%)=100×体重(g)/体长(cm)3;肝体比(HSI,%)=100×肝脏重/体重;脏体比(VSI,%)=内脏团重/体重×100;干物质表观消化率(%)=100×[1-(饲料中Y2O3含量/粪便中Y2O3含量)];蛋白质表观消化率(%)=100×[1-(粪便中粗蛋白质含量/饲料中粗蛋白质含量)×(饲料中Y2O3含量/粪便中Y2O3含量)]。
1.6数据处理与统计分析
数据采用SPSS 17.0软件进行统计学处理,并进行单因素方差分析(one-way ANOVA),当组间差异显著时,采用Tukey’s法进行多重比较,P<0.05为差异显著。试验数据用平均值±标准误(mean±SE)表示。
2 结 果
2.1高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼生长性能的影响
如表2所示,在终末体重、增重率和特定生长率方面,D1组与CON组相比显著升高(P<0.05),而与FM组无显著差异(P>0.05);在饲料效率方面,D1、D2组显著高于CON组(P<0.05),而与FM组无显著差异(P>0.05),D1组和D2组之间也无显著差异(P>0.05);在摄食率方面,各组间均无显著差异(P>0.05)。
2.2高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼形体指标的影响
如表3所示,在高植物蛋白质饲料中添加不同水平的丁酸钠对大菱鲆幼鱼的肥满度、肝体比均无显著影响(P>0.05);除D2组脏体比与FM组无显著差异(P>0.05)外,其他各组的脏体比均显著低于FM组(P<0.05)。
2.3高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼体组成的影响
如表4所示,高植物蛋白质饲料中添加不同水平的丁酸钠对大菱鲆幼鱼全鱼水分、粗蛋白质和粗脂肪含量均无显著影响(P>0.05)。
2.4高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼营养物质表观消化率的影响
如表5所示,干物质和蛋白质表观消化率随着丁酸钠添加水平的增加呈现先升高后降低的趋势,D1组的干物质和蛋白质表观消化率分别比CON组高6.93%和6.64%,差异显著(P<0.05),各试验组(D1、D2、D3组)的干物质表观消化率与FM组差异不显著(P>0.05),各试验组之间干物质表观消化率亦无显著差异(P>0.05);D1组的蛋白质表观消化率显著高于其他试验组和CON组(P<0.05),而与FM组差异不显著(P>0.05)。

表2 高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼生长性能的影响
同列数据肩标无字母或相同字母表示差异不显著(P>0.05),不同字母表示差异显著(P<0.05)。下表同。
In the same column, values with no letter or the same letter superscripts mean no significant difference (P>0.05), while with different small letter superscripts mean significant difference (P<0.05). The same as below.
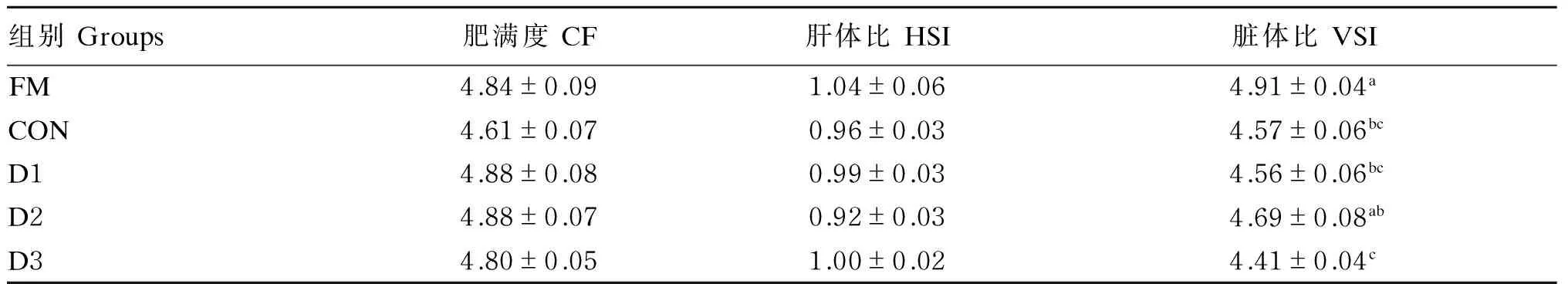
表3 高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼形体指标的影响

表4 高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼体组成的影响
2.5高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼肝脏抗氧化指标的影响
如表6所示,肝脏SOD活性随丁酸钠添加水平的增加呈先升高后降低的趋势,以D2组最高,与FM组差异不显著(P>0.05),显著高于D3组(P<0.05),相对于CON组有所升高但差异不显著(P>0.05);肝脏T-AOC随着丁酸钠添加水平的增加呈现逐渐升高的趋势,但各试验组之间无显著差异(P>0.05),且各试验组显著高于CON组(P<0.05),与FM组差异不显著(P>0.05);CON组肝脏MDA含量最高,显著高于FM、D2、D3组(P<0.05),与D1组差异不显著(P>0.05);肝脏CAT活性随着丁酸钠添加水平增加呈现先升高后降低的趋势,D1和D2组显著高于CON和D3组(P<0.05),与FM组差异不显著(P>0.05),D1和D2组之间无显著差异(P>0.05)。

表5 高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼营养物质表观消化率的影响
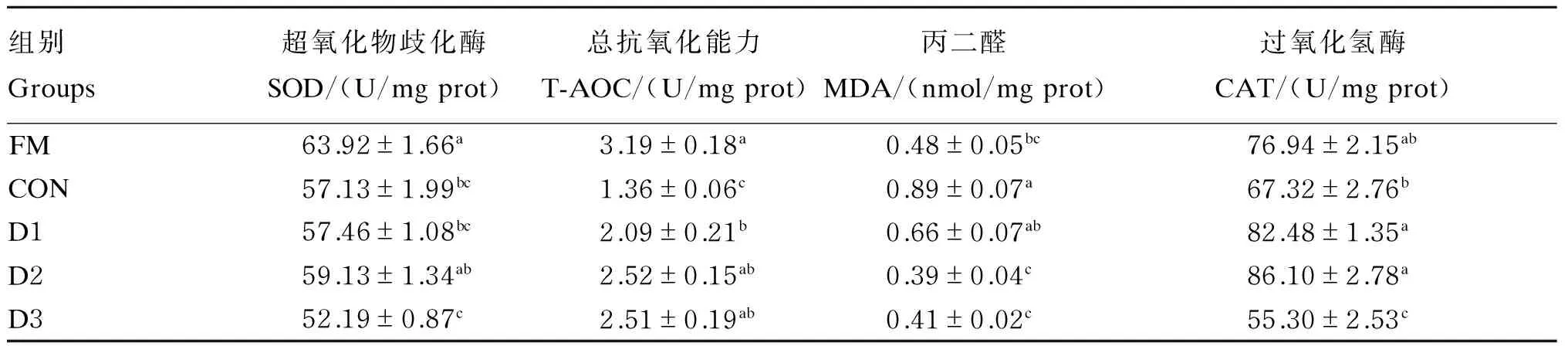
表6 高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼肝脏抗氧化指标的影响
3 讨 论
3.1高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼生长性能的影响
丁酸钠对不同动物生长性能的促进作用不尽相同,且最佳添加量也有所差异。研究发现,含0.05%丁酸钠的饲粮能显著影响生长兔的生长性能,并提高肠道健康状态[16]。饲粮中添加0.08%的丁酸钠对断奶仔猪的前期有显著的促生长作用,增重率显著高于对照组[17],这与本结果研究一致,即饲料中添加0.15%的丁酸钠能显著提高大菱鲆幼鱼的增重率。Liu等[12]研究发现,添加1 000或2 000 mg/kg丁酸钠的饲料均能显著提高草鱼的特定生长率。也有研究发现饲料中含0.2%的丁酸钠对鲈鱼的增重率和特定生长率并没有显著影响,但会显著影响免疫相关基因的表达,增强鲈鱼的免疫机能[18]。本研究结果表明,以复合植物蛋白质源替代饲料中50%的鱼粉后会影响大菱鲆幼鱼的生长性能,导致其增重率、饲料效率和特定生长率等降低,添加丁酸钠后可提高大菱鲆幼鱼的增重率、饲料效率及特定生长率,其中以0.15%添加组生长性能最好,终末体重、增重率等达到了FM组的水平,原因是:一方面,添加丁酸钠提高了大菱鲆幼鱼对营养物质的消化率;另一方面,添加丁酸钠提高了大菱鲆幼鱼的抗氧化能力。
在本研究中,添加过量的丁酸钠抑制了大菱鲆幼鱼的生长,这可能是由于过量的丁酸钠抑制了大菱鲆肠道的发育,降低了肠道对营养物质的消化利用率。在本研究中丁酸钠并没有对大菱鲆幼鱼的摄食率产生显著影响,这与张淞琳等[19]的研究结果类似,说明丁酸钠不会影响饲料的适口性[20],其对养殖鱼的促生长作用并不是通过提高摄食量来实现的,而是通过提高饲料的利用率来实现的。
3.2高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼体组成的影响
有研究表明,饲料中添加丁酸钠不会影响草鱼全鱼水分、粗蛋白质、粗脂肪和粗灰分含量[12]。翟秋玲等[21]研究表明,添加丁酸钠类似物三丁酸甘油酯不会影响菊黄东方鲀的鱼体粗蛋白质和粗脂肪含量。本试验结果显示,饲料中添加不同水平的丁酸钠没有对大菱鲆幼鱼全鱼水分、粗蛋白质和粗脂肪含量产生显著影响,与以上研究结果相似。也有研究发现丁酸钠会显著增加罗非鱼全鱼粗蛋白质含量,可能原因是丁酸钠促进摄入的食物更有效的转化为结构蛋白,从而使鱼体产生更多肌肉[22],但具体的机制还有待进一步研究。
3.3高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼营养物质表观消化率的影响
丁酸钠对营养物质的消化率有一定影响,Guilloteau等[23]研究表明丁酸钠对小牛营养物质消化率有积极影响。丁酸钠能够刺激胆囊收缩素释放来促进胰腺分泌胰液等物质,口服丁酸钠会刺激机体胰腺组织分泌胰肽酶E,使胰肽酶E的分泌量增加50%[24],还可以提高仔猪肠道不同部位的双糖酶活性,从而提高肠道消化酶活性[25];此外,丁酸钠还能促进断奶仔猪空肠后段的总蛋白酶、脂肪酶以及回肠淀粉酶的活性[26],从而促进机体能量沉积和蛋白质消化[27]。但Ribeiro等[28]对早期断奶的家兔研究表明,丁酸钠对其营养物质表观消化率并没有产生显著影响。本研究发现,饲料中植物蛋白质含量升高会造成大菱鲆幼鱼对干物质和蛋白质的表观消化率显著降低,添加一定量的丁酸钠后显著提高了大菱鲆幼鱼对饲料中干物质和蛋白质的表观消化率,其中以0.15%添加组效果最好,随着丁酸钠添加水平的增加,饲料中干物质和蛋白质的表观消化率有所降低。丁酸钠如何是提高大菱鲆幼鱼对饲料中营养物质的消化能力的还需进一步研究。
研究表明,不同收集粪便的方法会对养殖鱼类的营养物质表观消化率产生显著的影响,挤压法收集粪便测得的干物质和蛋白质表观消化率显著低于虹吸法[29],这是因为收集到的粪便易混入未消化的饵料、体液等,导致营养物质表观消化率偏低,而体外收粪法易造成营养物质在水中溶失,这也是虹吸法测得的营养物质表观消化率偏高的原因之一,但这2种收集粪便的方法均存在其合理性[30-31],由于本研究收集粪便采用的挤压法,因此测得的干物质和蛋白至表观消化率偏低。此外,营养物质表观消化率的高低也和饲料的加工工艺有关,本试验采用的冷挤压法制作饲料,这也导致了营养物质表观消化率偏低。
3.4高植物蛋白质饲料中添加不同水平丁酸钠对大菱鲆幼鱼肝脏抗氧化功能的影响
丁酸钠通过调节机体免疫和抗氧化功能来增强机体清除自由基的能力,减少组织和细胞损伤[32],缓解不良外界因素带来的负面影响。SOD能清除超氧阴离子自由基,保护细胞免受损伤;T-AOC的强弱与健康程度存在密切联系,该防御体系由酶促和非酶促2个体系组成,协同防护机体氧化;MDA含量的高低间接反映了机体细胞受自由基攻击的严重程度;CAT能分解过氧化氢,减轻机体过氧化程度。有研究表明,丁酸钠能显著提高奶牛血清SOD活性和T-AOC[33];在脂多糖(LPS)应激条件下,丁酸钠能显著提高肉鸡血清和肝脏SOD、CAT活性,并降低血清和肝脏MDA含量,从而改善营养代谢,维持机体抗氧化功能,增强抗炎功能[34];饲料中添加0.1%的丁酸钠能使鳗鱼肝脏T-AOC和CAT活性分别提高25%和15%,肝脏MDA含量降低15%[19]。本研究结果表明,复合植物蛋白质源替代饲料中50%鱼粉后对大菱鲆幼鱼肝脏的抗氧化指标产生了显著影响,肝脏SOD和CAT活性及T-AOC均随丁酸钠添加水平的增加呈先升高后降低的趋势,其中0.15%添加组的T-AOC和CAT活性均显著高于CON组,在添加水平为0.30%时达到最高值,使其达到了FM组水平,且丁酸钠的添加降低了肝脏MDA含量。这表明丁酸钠能在一定程度上能缓解植物蛋白质替代鱼粉产生的氧化应激,增强养殖鱼在应激条件下的抗氧化能力,这对增强机体的免疫能力具有一定意义。
综合分析得出,丁酸钠的添加水平在0.30%时大菱鲆肝脏的抗氧化能力最强,但此添加水平时养殖鱼的生长性能已开始下降,说明机体抗氧化能力最高时其生长性能不一定最好,因为此时机体耗能增多,导致生长缓慢,这与机体免疫能力最高时,在此核苷酸添加水平下黄颡鱼[35]和大菱鲆[36]未表现出最佳的生长性能的研究相似;此外,0.30%添加组的终末体重和增重率与0.15%添加组相比显著降低,饲料效率和特定生长率也呈下降趋势,这可能其与蛋白质表观消化率显著降低有关。因此,在本试验条件下,综合生长性能和肝脏抗氧化指标,建议丁酸钠的添加水平为0.15%。研究表明,高植物蛋白质饲料易引起大菱鲆幼鱼肠道损伤[37],具体表现为肠壁肌层变薄、肠绒毛损伤变短和杯状细胞增多等,导致肠道重量下降[38],这可能是脏体比降低的主要原因,0.30%添加组的脏体比与FM组差异不显著,说明丁酸钠可能对修复肠道损伤有一定的效果[39-40]。
3.5丁酸钠作为新型添加剂在鱼粉替代中的意义
以往对大菱鲆饲料中鱼粉替代的研究表明,植物蛋白质替代大菱鲆饲料中鱼粉的比例有限。许丹丹等[41]研究发现,以单一蛋白质源玉米蛋白粉替代45%的鱼粉蛋白显著抑制大菱鲆幼鱼的生长;豆粕可替代大菱鲆饲料中30%的鱼粉,替代后对增重率、蛋白质效率和特定生长率的影响不显著[42]。复合植物蛋白质也只能替代鲆鲽类饲料中较低水平的鱼粉,Wang等[43]发现复合植物蛋白质替代大菱鲆饲料中40%的鱼粉会影响其生长性能;Imsland等[44]的研究能将复合植物蛋白质替代大菱鲆饲料鱼粉的比例提高到43%而不影响其生长;Liu等[9]报道,复当合植物蛋白质替代大菱鲆饲料50%的鱼粉时,大菱鲆幼鱼的增重率和特定生长率显著下降,本研究在此研究的基础上向以复合植物蛋白质替代50%鱼粉后的饲料中添加营养性添加剂——丁酸钠,使得大菱鲆幼鱼的生长性能未出现显著降低,提高了植物蛋白质在大菱鲆饲料中的使用比例,主要原因是适量的丁酸钠促进了大菱鲆幼鱼对营养物质的消化率,又不影响其适口性。本研究不仅为丁酸钠在水产动物养殖中的综合利用提供了一定的科学依据,而且为饲料工业中提高植物蛋白质源的利用率提供理论依据。
4 结 论
① 在高植物蛋白质饲料中添加适量丁酸钠不仅能够促进大菱鲆幼鱼对饲料的消化,进而提高生长性能,还可以提高机体的抗氧化功能,同时对体组成没有产生显著影响,但过量添加丁酸钠会降低大菱鲆幼鱼的生长性能。
② 在本试验条件下,综合生长性能和肝脏抗氧化指标,建议丁酸钠的添加水平为0.15%。
[1] 许丹丹.大菱鲆幼鱼对不同蛋白源营养感知与应答机制的初步研究[D].博士学位论文.青岛:中国海洋大学,2014:3-6.
[2] FRANCIS G,MAKKAR H P S,BECKER K,et al.Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish[J].Aquaculture,2001,199(3/4):197-227.
[3] PENN M H,BENDIKSEN E A,CAMPBELL P,et al.High level of dietary pea protein concentrate induces enteropathy in Atlantic salmon (SalmosalarL.)[J].Aquaculture,2011,310(3/4):267-273.
[4] SILVA F C P,NICOLI J R,ZAZBONINO-INFANTE J L,et al.Influence of partial substitution of dietary fish meal on the activity of digestive enzymes in the intestinal brush border membrane of gilthead sea bream,Sparusaurataand goldfish,Carassiusauratus[J].Aquaculture,2010,306(1/2/3/4):233-237.
[6] BONALDO A,ROEM A J,FAGIOLI P,et al.Influence of dietary levels of soybean meal on the performance and gut histology of gilthead sea bream (SparusaurataL.) and European sea bass (DicentrarchuslabraxL.)[J].Aquaculture Research,2008,39(9):970-978.
[7] GAYLORD T G,BARROWS F T,TEAGUE A M,et al.Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchusmykiss)[J].Aquaculture,2007,269(1/2/3/4):514-524.
[8] YUN B,MAI K S,ZHANG W B,et al.Effects of dietary cholesterol on growth performance,feed intake and cholesterol metabolism in juvenile turbot (ScophthalmusmaximusL.) fed high plant protein diets[J].Aquaculture,2011,319(1/2):105-110.
[9] LIU Y Z,HE G,WANG Q C,et al.Hydroxyproline supplementation on the performances of high plant protein source based diets in turbot (ScophthalmusmaximusL.)[J].Aquaculture,2014,433:476-480.
[10] BURRELLS C,WILLIAMS P D,SOUTHGATE P J,et al.Dietary nucleotides:a novel supplement in fish feeds:2.Effects on vaccination,salt water transfer,growth rates and physiology of Atlantic salmon (SalmosalarL.)[J].Aquaculture,2001,199(1/2):171-184.
[11] 李丹丹,冯国强,钮海华,等.丁酸钠对断奶仔猪生长性能及免疫功能的影响[J].动物营养学报,2012,24(2):307-313.
[12] LIU M M,GUO W,WU F,et al.Dietary supplementation of sodium butyrate may benefit growth performance and intestinal function in juvenile grass carp (Ctenopharyngodonidellus)[J].Aquaculture Research,2016,doi:10.1111/are.13230.
[13] CLAUS R,GUNTHNER D, LETZGUΒ H.Effects of feeding fat-coated butyrate on mucosal morphology and function in the small intestine of the pig[J].Journal of Animal Physiology and Animal Nutrition,2007,91(7/8):312-318.
[14] PIVA A,PIZZAMIGLIO V,MORLACCHINI M,et al.Lipid microencapsulation allows slow release of organic acids and natural identical flavors along the swine intestine[J].Journal of Animal Science,2007,85(2):486-493.
[15] 杨传哲,何艮,周慧慧,等.大菱鲆幼鱼对几种蛋白质源中营养物质的表观消化率及膨化处理对其产生的影响[J].动物营养学报,2016,28(7):2045-2054.
[16] HASSANIN A,TONY M A,SAWIRESS F A R,et al.Influence of dietary supplementation of coated sodium butyrate and/or synbiotic on growth performances,caecal fermentation,intestinal morphometry and metabolic profile of growing rabbits[J].Journal of Agricultural Science,2015,7(2):9752-9760.
[17] PIVA A,MORLACCHINI M,CASADEI G,et al.Sodium butyrate improves growth performance of weaned piglets during the first period after weaning[J].Italian Journal of Animal Science,2002,1(1):35-41.
[18] TEROVA G,DIAZ N,RIMOLDI S,et al.Effects of sodium butyrate treatment on histone modifications and the expression of genes related to epigenetic regulatory mechanisms and immune response in European sea bass (Dicentrarchuslabrax) fed a plant-based diet[J].PLoS One,2016,11(7):e0160332.
[19] 张淞琳,常建波,叶继丹,等.丁酸钠对美洲鳗鲡摄食、生长性能和抗氧化能力的影响[J].福建农业学报,2011,26(4):549-551.
[20] ROBLES R,LOZANO A B,SEVILLA L,et al.Effect of partially protected butyrate used as feed additive on growth and intestinal metabolism in sea bream (Sparusaurata)[J].Fish Physiology and Biochemistry,2013,39(6):1567-1580.
[21] 翟秋玲,张春晓,孙云章,等.三丁酸甘油酯和甘露寡糖对菊黄东方鲀生长性能、体组成及肠道健康指标的影响[J].动物营养学报,2014,26(8):2197-2208.
[22] AHMED H A,SADEK K M.Impact of dietary supplementation of sodium butyrate and/or protexin on the growth performance,some blood parameters,and immune response ofOreochromisniloticus[J].International Journal of Agriculture Innovations and Research,2015,3(4):985-991.
[23] GUILLOTEAU P,SAVARY G,JAGUELIN-PEYRAULT Y,et al.Dietary sodium butyrate supplementation increases digestibility and pancreatic secretion in young milk-fed calves[J].Journal of Dairy Science,2010,93(12):5842-5850.
[24] ARAUJO G,TERRÉ M,MEREU A,et al.Effects of supplementing a milk replacer with sodium butyrate or tributyrin on performance and metabolism of Holstein calves[J].Animal Production Science,2016,56(11):1834-1841.
[26] 陈国顺,徐振飞,赵芳芳,等.包膜丁酸钠对断奶仔猪生长性能和肠道消化酶活性的影响[J].国外畜牧学-猪与禽,2011,31(2):65-68.
[27] MALLO J J,PUYALTO M,RAMA RAO S V.Evaluation of the effect of sodium butyrate addition to broiler diet on energy and protein digestibility,productive parameters and size of intestinal villi of animals[J].Feed Livestock,2012,8:26-30.
[28] RIBEIRO J,GASPAR S,PINHO M,et al.Sodium butyrate in growing and fattening diets for early-weaned rabbits[J].World Rabbit Science,2012,20(4):199-207.
[29] 史东杰,孙向军,梁拥军,等.粪便不同收集方法对锦鲤表观消化率测定结果的影响[J].水产科技情报,2013,40(5):266-271.
[30] SHIPTON J A,BRITZ P J.An assessment of the use of chromic oxide as a marker in protein digestibility studies withHaliotismidaeL.[J].Aquanculture,2001,203(1/2):69-83.
[31] RAMSAY J M,CASTELL J D,ANDERSON D M,et al.Effects of fecal collection methods on estimation of digestibility of protein feedstuffs by winter flounder[J].Northern American Journal of Aquaculture,2000,62(3):168-173.
[32] 岳敏,许丽,方翠林,等.丁酸钠对母猪生产性能和抗氧化功能及后代仔猪生长的影响[J].中国畜牧杂志,2014,50(23):44-47.
[33] 阿明古丽·牙生,余雄,李胜利,等.丁酸钠对围生期奶牛免疫和抗氧化功能的影响[J].饲料研究,2015,5(5):30-34.
[34] 鞠婷婷,郭孝烨,随佳佳,等.不同剂型丁酸钠对脂多糖应激肉鸡血清生化指标、抗氧化和抗炎功能的影响[J].动物营养学报,2015,27(10):3146-3154.
[35] ZHAO H X,CAO J M,HUANG Y H,et al.Effects of dietary nucleotides on growth,physiological parameters and antioxidant responses of juvenile yellow catfishPelteobagrusfulvidraco[J].Aquaculture Research,2017,48(1):214-222.
[36] MENG Y,MA R,MA J,et al.Dietary nucleotides improve the growth performance,antioxidative capacity and intestinal morphology of turbot (Scophthalmusmaximus)[J].Aquaculture Nutrition,2016,doi:10.1111/anu.12425.
[37] ROMARHEIM O H,ZHANG C,PENN M,et al.Growth and intestinal morphology in cobia (Rachycentroncanadum) fed extruded diets with two types of soybean meal partly replacing fish meal[J].Aquaculture Nutrition,2008,14(2):174-180.
[38] CAI C F,WANG W J,YE Y T,et al.Effect of soybean meal,raffinose and stachyose on the growth,body composition,intestinal morphology and intestinal microflora of juvenile allogynogenetic silver crucian carp (Carassiusauratusgibelio♂×Cyprinuscarpio♀)[J].Aquaculture Research,2012,43(1):128-138.
[39] LIU W S,YANG Y O,ZHANG L L,et al.Effects of dietary microencapsulated sodium butyrate on growth,intestinal mucosal morphology,immune response and adhesive bacteria in juvenile common carp (Cyprinuscarpio) pre-fed with or without oxidised oil[J].British Journal of Nutrition,2014,112(1):15-29.
[40] IRAPORDA,ERREA A,ROMANIN D E,et al.Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells[J].Immunobiology,2015,220(10):1161-1169.
[41] 许丹丹,何艮,周慧慧,等.玉米蛋白粉部分替代鱼粉对大菱鲆幼鱼的生长性能和游离氨基酸代谢的影响[J].中国海洋大学学报:自然科学版,2016,46(2):26-34.
[42] WANG L,ZHOU H H,HE R J,et al.Effects of soybean meal fermentation byLactobacillusplantarumP8 on growth,immune responses,and intestinal morphology in juvenile turbot (ScophthalmusmaximusL.)[J].Aquaculture,2016,464:87-94.
[43] WANG Q,HE G,MAI K,et al.Fishmeal replacement by mixed plant proteins and maggot meal on growth performance,target of rapamycin signalling and metabolism in juvenile turbot (ScophthalmusmaximusL.)[J].Aquaculture Nutrition,2016,22(4):752-758.
[44] IMSLAND A K D,HELMVIG T,KRISTJANSSON G Ö,et al.Effect of fish protein replacement in diets for juvenile turbot (ScophthalmusmaximusL.)[J].Turkish Journal of Fisheries and Aquatic Sciences,2016,16(2):267-273.
*Corresponding author, professor, E-mail: hegen@ouc.edu.cn
(责任编辑 菅景颖)
Effects of Adding Different Levels of Sodium Butyrate in High Plant Protein Diets on Growth Performance, Nutrient Apparent Digestibility Coefficients and Liver Antioxidant Function of Juvenile Turbot (Scophthalmus maximus L.)
WEI Chaoqing ZHOU Huihui WANG Xuan MAI Kangsen XU Wei HE Gen*
(Key Laboratory of Aquaculture Nutrition and Feeds, Fisheries College, Ocean University of China, Qingdao 266003, China)
In this study, a feeding trial was conducted to evaluate the effects of adding different levels of sodium butyrate in high plant protein diets on growth performance, nutrient apparent digestibility coefficients and liver antioxidant function of juvenile turbot (ScophthalmusmaximusL.), and to explore the feasible method for improving substitution of fish meal by plant protein. Using fish meal, soybean meal, corn gluten meal, vital gluten, peanut meal and beer yeast as protein sources, fish oil, coconut oil and soybean lecithin as lipid sources, and wheat meal as carbohydrate source, five isonitrogen and isocaloric experimental diets were formulated. Among them, the basal diet formulated with 60% fish meal was set as the positive control group (FM group), the substituted diet which used mixed plant protein sources to replace 50% fish meal was set as the negative control group (CON group), and the other three diets which were supplemented with 0.15% (D1 group), 0.30% (D2 group) and 0.60% (D3 group) sodium butyrate into the substituted diet, respectively, were set as the experimental groups. Each diet was assigned to three replicates of 30 juvenile turbot with an initial body weight of (13.00±0.01) g for 58 days in an indoor aquaculture system. The results showed as follows: 1) with the increase of sodium butyrate supplemental level, the weight gain rate (WGR), specific growth rate (SGR) and feed efficiency (FE) were increased firstly and decreased subsequently. The WGR, SGR and FE in FM, D1 and D2 groups were significantly higher than those in CON group (P<0.05), while no significant difference in feed intake (FI) was observed among groups (P>0.05). For the viscerosomatic index (VSI), compared with FM group, the D2 group had no significant difference (P>0.05), but the other group were significantly decreased (P<0.05). No significant differences in whole fish moisture, crude protein and crude lipid contents were observed among groups (P>0.05). 2) The apparent digestibility coefficient of dry matter in D1 group was significantly higher than that in CON group (P<0.05), while it showed no significant difference between the D1 and FM groups (P>0.05). For the apparent digestibility coefficient of protein, D1 group was significantly higher than that in CON, D2 and D3 groups (P<0.05), but no significant differences were showed between the D1 and FM groups (P>0.05). No significant difference in the apparent digestibility coefficients of dry matter and protein were observed between the D2 and D3 groups (P>0.05). 3) Compared with the CON group, liver total antioxidant capacity (T-AOC) and catalase (CAT) activity in D2 group were significantly increased (P<0.05), but the liver malondialdehyde (MDA) content was significantly decreased (P<0.05). In conclusion, under this experimental condition, adding 0.15% sodium butyrate in the high plant protein diets can improve the growth performance, nutrient apparent digestibility coefficients and liver antioxidant function of juvenile turbot, but excessive adding sodium butyrate will decrease the growth performance of juvenile turbot.[ChineseJournalofAnimalNutrition,2017,29(9):3392-3402]
sodium butyrate; juvenile turbot; growth performance; apparent digestibility; antioxidant ability
10.3969/j.issn.1006-267x.2017.09.045
2017-02-15
替代渔用饲料中鱼粉的新蛋白源开发利用技术(201303053);国家自然科学基金面上项目(31572627)
魏朝青(1990—),男,山东德州人,硕士研究生,从事水产动物营养与饲料学研究。E-mail: 780071148@qq.com
*通信作者:何 艮,教授,博士生导师,E-mail: hegen@ouc.edu.cn
S963
:A
:1006-267X(2017)09-3392-11
猜你喜欢
黑龙江水产(2022年6期)2022-12-03
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2019年6期)2019-07-25
当代水产(2019年1期)2019-05-16
中国水产(2019年3期)2019-03-25
浙江农业学报(2016年7期)2016-06-15
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
应用海洋学学报(2014年1期)2014-11-22
