
额颞脑区在概念扩展中的作用及其与大五人格的关系*
2017-09-15 08:12尹德圣马军朋杨楠金花朱朝喆林崇德
心理与行为研究 2017年4期
尹德圣马军朋杨 楠金 花朱朝喆林崇德
(1教育部人文社会科学重点研究基地天津师范大学心理与行为研究院,天津 300074)(2北京师范大学认知神经科学与学习国家重点实验室,北京 100875)
额颞脑区在概念扩展中的作用及其与大五人格的关系*
尹德圣1马军朋1杨 楠1金 花1朱朝喆2林崇德2
(1教育部人文社会科学重点研究基地天津师范大学心理与行为研究院,天津 300074)(2北京师范大学认知神经科学与学习国家重点实验室,北京 100875)
对概念扩展神经机制的探讨日益成为创造性认知 “过程式”研究视角下的热点。但以往研究未能排除评估效应对实验的影响,亦未考察过人格因素与任务态下神经活动的关系。实验向被试随机呈现高低扩展两类的报纸用途,要求被试认真思考 “合理的操作方式”,使报纸能够实现呈现的用途。同时对其进行fNIRS的兴趣区扫描,并在事后要求被试完成大五人格量表。结果发现,与低扩展条件相比,高扩展条件会引起颞叶区域 (BA21/ 22)更大的激活,而额极 (BA10)和背外侧前额皮层 (BA9)的激活会显著降低;人格元特质可塑性与额极和眶额皮层的活动正相关。提示概念扩展过程中,可能需要颞叶脑区激活与前额脑区去激活的协同作用,且该过程与人格因素存在相关关系。
创造性加工,概念扩展,前额,去激活,元特质。
1 引言
创造性是人类特有的一种信息加工能力 (Boden,1998;Dietrich,2004),是推动人类文明不断进步和发展的核心要素。生成-探索模型 (The Geneplore model,Finke,Ward,&Smith,1992;Sternberg, 1999)认为,创造性至少可以包括概念扩展 (conceptual expansion)、创造性想象 (creative imagery)、知识固着的突破 (overcoming knowledge constraints)以及顿悟 (insight)四种认知加工成分。对比以往研究对 “创造性认知是多种认知操作过程联合加工结果 (Abraham,2014)”这一问题的忽视,以至于限制了不同认知加工过程中不同脑区所起的作用,该理论为创造性脑机制研究提供了 “过程式”(Kozbelt,Beghetto,&Sternberg,2010)的新研究视角,打破了以往大多数研究将创造性视作单一的认知过程 (Abraham,2014;Abraham et al.,2012; Kröger et al.,2012,2013;Rutter et al.,2012a;Rutter et al.,2012b)的局限。基于该视角,本研究主要考察额颞脑区在概念扩展这一创造性认知加工过程的作用。
概念扩展的核心在于扩展已有概念的界限以获得新的含义 (Abraham et al.,2012;Kröger et al., 2012,2013;Rutter et al.,2012a;Rutter et al., 2012b)。目前,在脑成像研究中,该领域普遍采用的实验任务主要源自用途报告任务以及隐喻任务(Abraham et al.,2012;Kröger et al.,2012,2013; Rutteretal.,2012a,2012b), 包括 “主动”(Abraham et al.,2012)和 “被动” (Kröger et al.,2012;Rutter et al.,2012a,2012b)两种范式。前者,被试需要自发地对刺激的新颖用途进行报告;后者,研究者给被试呈现刺激的用途,或者潜在喻意,需要被试对呈现目标进行新颖性和适用性方面的评估。当他们认为评定目标新颖且合适时,就认为发生了概念扩展。这两种范式均通过被试“尽可能地进行新颖性报告”或对目标刺激的新颖性和适用性评估来诱发概念扩展。这样的任务操作很可能混杂着较大权重的认知评估过程,而导致概念扩展和认知评估相关功能脑区的同时激活,且难以相互分离。
此外,就创造性而言,除了上述任务相关的状态创造性之外,与人格因素密切相关的特质创造性亦是不容忽略的重要方面 (Amabile,1996;Gough, 1979;Li et al.,2014;Piffer,2012;Rhodes,1961;Runco,2014)。然而,相关研究多局限于创造性特质与脑结构关系的探讨,如Li等 (2014)发现,右颞中回后侧的灰质总量与创造性特质呈正相关,且发现了经验开放性中介于上述关系;Jauk,Neubauer,Dunst,Fink和Benedek(2015)发现,经验开放性与默认网络 (default mode network, DMN)之间的关联,如负相关于楔前叶的灰质容积。Leutgeb等 (2016)发现,边界型人格障碍者(borderline personality disorder)额下回和额中回的眶额部分的灰质密度下降和创造性水平低相关。迄今,据我们所知,极少有研究探讨个体执行创造性认知任务过程中伴随的脑功能活动与人格因素之间的关系。Fink等人 (2014)考察了分裂型人格特质与个体主动报告客体用途条件下脑活动模式的关系,但其在实验过程中也未控制认知评估因素的影响,有必要在控制认知评估因素之后,进一步考察个体执行创造性认知任务过程中伴随的脑功能活动与人格因素之间的关系。
综上所述,本研究拟从概念扩展出发,进一步考察创造性认知加工过程的神经机制,丰富和完善人类创造性神经机制的研究。研究以报纸用途报告任务为实验任务,并预先通过材料实验将报纸用途分成了高 (偏新颖性,如 “饲料”)、低 (普通用途,如 “练字”)扩展性两类,将它们随机呈现给被试,这样便人为地避免了被试在正式实验中对报纸用途进行认知评估从而混淆实验结果。实验要求他们认真思考呈现用途与报纸之间的合理联结——通过合理的操作方式,使得报纸能够实现这种用途。通过对比执行高低扩展性任务间的神经活动,既可揭示概念扩展特定的神经机制,也有效控制了评估因素对扩展过程的影响。实验中,我们采用高生态效度的功能性近红外光学脑成像技术 (functional nearinfrared spectroscopy,fNIRS)进一步考察概念扩展的神经基础,特别是额颞脑区在概念扩展中的作用,以丰富和完善人类创造性神经机制的研究。
结合先前研究以及颞叶脑区在新颖语义形成方面的特殊作用 (Albright,2012;Hoffman,Evans,& Ralph,2014;Lau,Phillips,& Poeppel,2008; Rogers&McClland,2004),我们预期颞叶区可能在概念扩展过程中显著激活。由于额极广泛参与信息的内在自我评估 (Christoff&Gabrieli,2000),而本研究排除了认知评估过程,只考察单一的扩展加工,因此额极等前额脑区在本实验中可能并不会呈现显著激活状态。在此基础上,研究进一步结合大五人格量表 (朱小佳,2014)获取的数据探讨该神经活动与人格之间的关联。Li等 (2014)的研究表明特质创造性越高,眶额灰质总量越低,越倾向于直觉性思维/冒险性加工。换言之,在结构上已预先具有了冒险性加工倾向优势的高特质创造性个体,在任务态下的眶额活动可能较低的弱化 (眶额皮层的活动弱化与利于创造性发生的冒险性选择加工相关,Jonker,Jonker,Scheltens,&Scherder, 2015)就能满足需要。因此,研究假设眶额活动可能与特质创造性正相关。
2 方法
2.1 被试
22名未参加预实验的大学生有偿参加了本实验。其中,4名被试因利手等原因被剔除。最终,18名有效被试 (男7名,女11名)参与数据分析,年龄范围21-26岁 (7名男生,平均年龄23.50± 1.50岁,年龄范围21-26岁)参与数据分析。所有被试均为右利手,视力或矫正视力正常,无色盲,无脑损伤,无神经疾病或者精神病史,且近期未服用过安定类药物。实验获得学院道德伦理委员会批准。实验前所有被试均填写了知情同意书。
2.2 实验材料
2.2.1 报纸用途任务
以徐芝君,陈学志和邱发忠 (2012)编制的《<报纸的不寻常用途>测验》为初始实验材料,经过适当调整 (将繁体描述用途转换成等意的简体双字词,以及使表述更符合大陆表述习惯)后,编制成7级评定量表,用途项目随机排列。由10名大学生根据各个用途与报纸之间建立合理联系的难易程度进行等级评定,等级越高表明联结越不易产生,概念扩展越高。得到高低扩展组各60个用途刺激 (高扩展组H,如饲料;低扩展组L,如练字)。两组平均联结难度分别为:5.36±0.47;2.76±0.55。内部一致性系数αH=0.98,αL=0.94。
2.2.2 大五人格量表
朱小佳的修订版 (2014)。该量表共包括44个项目,涉及神经质 (N)、外倾性 (E)、开放性(O)、宜人性 (A)以及尽责性 (C)等五个维度,并进一步划分为两个元特质维度:可塑性和稳定性。前者涵盖了外倾性和开放性,后者涵盖了神经质、宜人性和尽责性,相比五个低阶维度,可塑性和稳定性这两个高阶维度可以更好的预测创造性(Silvia,Nusbaum,Berg,Martin,& O′Connor,2009)。量表采用5级等级评定,等级越高表明越倾向于同意量表条目表述。每个维度得分由其涉及的各项目得分相加获得。
2.2.3 事后问卷
自编量表,要求被试对实验中用途刺激材料的新颖性、联结难易分别进行5级等级评定,等级越高表示越新颖、越难联结。
2.3 实验设计
本实验为单因素设计,自变量为任务类型,分为H和L扩展两个水平。因变量有两类,一类为行为学指标,包括反应时、事后新颖性和联结难易的评定等级、人格量表得分;一类为光学成像指标,反映氧合血红蛋白浓度变化的beta值。
近红外光学脑成像实验采用事件相关设计,包括120个trial,每个trial由注视点 (+)、用途刺激构成。实验时先呈现10s空屏,使前期信号稳定下来,随后开始呈现trial。每个trial注视点呈现时间在2-6s之间,之后出现用途刺激 (如 “练字”),呈现时间为5s。要求被试在注视点呈现期间心情尽量保持平静,当屏幕上随机出现用途刺激 (H、L刺激随机呈现)时,认真思考该用途与报纸之间的合理联结——通过合理的操作方式,使得报纸能够实现这种用途。想到后立即按键反应,进入下一trial。实验流程见图1。

图1 概念扩展实验流程图
提醒被试看见刺激即开始思考,只要视野范围内有刺激就不能停止思考,直至想到与报纸之间的合理联系或刺激呈现时间 (5s)结束。除了想到合理联结按键反应后进入下一trial外,刺激呈现时间 (5s)结束后也自动进入下一trial。
2.4 实验程序
被试进入实验室后,先填写知情同意书,并完成利手测验。随后开始fNIRS实验,在被试执行概念扩展任务期间,对其进行fNIRS扫描。
实验开始之前,要求被试坐在近红外实验室内。主试根据国际10-20系统。主试给被试佩戴好光极帽,并在相应位置插入光极。然后,要求被试实验时两眼注视电脑屏幕中央,眼睛距离屏幕65cm左右。正式实验前被试先进行练习以熟悉实验程序和要求。实验完成后,除去被试光极帽,并指导其填写完成事后评定问卷以及大五人格量表。
2.5 数据采集及处理
2.5.1 数据采集
采用日本-岛津公司生产的多通道功能性近红外光谱系统LABNIRS采集数据,应用三种波长(780mm、805mm、830mm)的近红外线半导体激光检测皮层血氧变化。因fNIRS测量指标中氧合血红蛋白信噪比最佳 (如Ozawa,Matsuda,&Hiraki, 2014),故本研究选取其浓度变化为脑功能性评定指标。计算基于矫正后的Beer-Lambert定律。血红蛋白浓度变化的信号采样率设定为10Hz。
2.5.2 通道布局
采用3×9光极帽进行多通道布局,包括14个反射光极和13个探测光极,两类光极探头间隔排布,相邻间隔距离为3cm。发射光极与相邻的探测光极之间构成一个通道,这样共涉及42个通道。光极定位以国际10-20系统为参照。鉴于已有研究尽管具体的实验范式和激活脑功能区有所差别,但均发现了额颞位置在概念扩展过程中的参与作用(Abraham et al.,2012;Kröger et al.,2012,2013; Rutter et al.,2012a,2012b),所以本研究设置的光极位置主要覆盖前额和颞叶部分的脑区。沿中线“鼻根-FPz-Fz-Cz-Pz-Oz-枕骨隆凸”左右对称分布,以鼻根上10%的 “鼻根-枕骨隆凸”距离为光极帽底排中间光极点的位置,并以此为基布局剩余光极点。
另外,以3D定位仪为辅助工具将fNIRS通道位置与MNI空间进行概率配准,模拟计算各通道对应的大脑结构的三维信息。
2.5.3 数据处理
使用SPSS16.0对被试事后等级评定数据进行分析,进一步检验实验材料的有效性。分析分别以被试对H、L的反应时、新颖性以及联结难易的评定等级为指标,进行配对样本t检验,并对新颖性和联结难易评定等级进行了相关分析。
使用基于2008b的NIRS-SPM软件包处理近红外数据。在个体水平上,主要包括以下几个步骤: (1)数据预处理,采用 CSBI(correlation based signalimprovement,Cui,Bray,& Reiss, 2010)与 Wavelet相结合的方法 (Jang et al.,2009)去噪,去漂移; (2)利用血液动力学反应函数 (hrf)低通滤波; (3)基于一般线性模型(GLM)评估数据。另外,需要指出的是,在5s内未能想到与报纸之间合理联结的事件仍纳入H事件,因为本实验关注的是个体概念扩展过程相关的神经活动。虽然个体未能在规定时间内想到合理联结,但在整个刺激呈现期间,其都在进行概念扩展的操作性思考; (4)经过评估,得到并提取出任务相关的beta值,主要为H、L事件分别与基线(注视点)比较后的两类beta值。
在群体水平分析中,将提取的 beta值导入SPSS16.0,进行下列统计检验: (1)L组beta值的单样本t检验; (2)H组beta值的单样本t检验; (3)H组vs.L组beta值的配对样本t检验。所有检验结果以p<0.05以及 Overlap(可能覆盖率%)>10为筛选阈限。并且,根据被试量表施测得分,对量表各维度与各个通道的激活程度进行相关分析。
3 结果
3.1 行为结果
对两组材料的反应时进行配对样本t检验,t(59)=11.31,p<0.0001,d=2.02,H组 (3.00± 0.74)显著长于L组 (1.79±0.40),见图2a。对两组材料间新颖性的评定等级进行配对样本t检验,t(59)=17.49,p<0.0001,d=3.19,H组 (2.71± 0.42)显著大于L组 (1.52±0.30),见图2b。
对两组材料间联结难易 (扩展高低)的评定等级进行配对样本 t检验,t(59)=17.04,p<0.0001,d=3.11,H组 (2.61±0.48)显著大于L组(1.46±0.29),见图2c。
对材料的新颖性和联结难易 (扩展高低)进行相关分析,结果显示r=0.975,p<0.0001,二者显著正相关,见图2d。
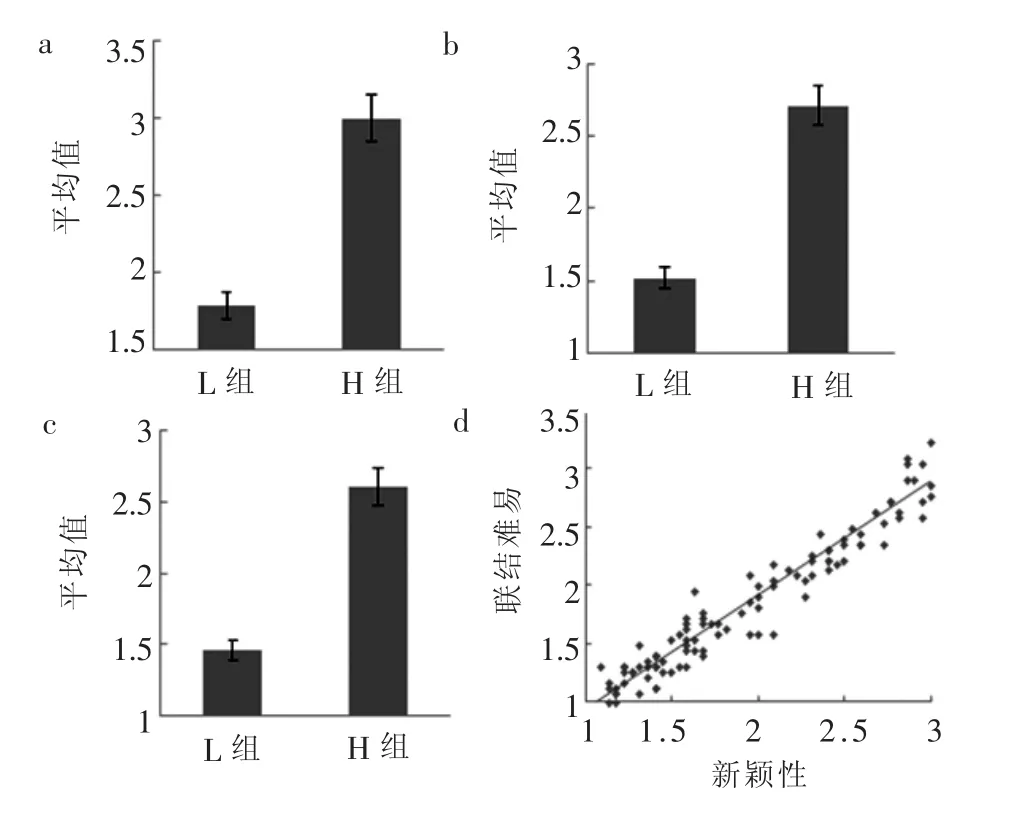
图2 行为数据结果示图
3.2 脑成像结果
3.2.1 与基线比较,高、低扩展 (L)条件下通道活动情形
与基线相比,个体执行L任务时,通道22(覆盖脑区为额极 (BA10))、26(覆盖脑区为颞上回(BA22)、颞中回 (BA21)、前辅助运动区 (BA6))、30(覆盖脑区为额极 (BA10))的激活水平达到了显著性水平。即,L条件下到达显著性水平的主要脑区为双侧额极和左颞上回,详见表1。

表1 L条件下激活通道及对应脑区*
与基线相比,H条件下,共有6个通道达到了显著性激活水平,主要为双侧激活,包括额眼区、背外侧前额叶皮层、额极以及眶区,见表2。
3.2.2 高扩展vs.低扩展大脑通道活动情形
与低扩展条件比较,高扩展条件下通道26覆盖的脑区 (右颞上回 (BA22)、右颞中回(BA21))显著正激活;通道 3(右前额皮层(BA8/9)、通道11(右背外侧前额皮层 (BA9/46)、右额极 (BA10))、通道14(左背外侧前额皮层(BA9)、左额极 (BA10))显著去激活,见表3。

表2 H条件下激活通道及对应脑区
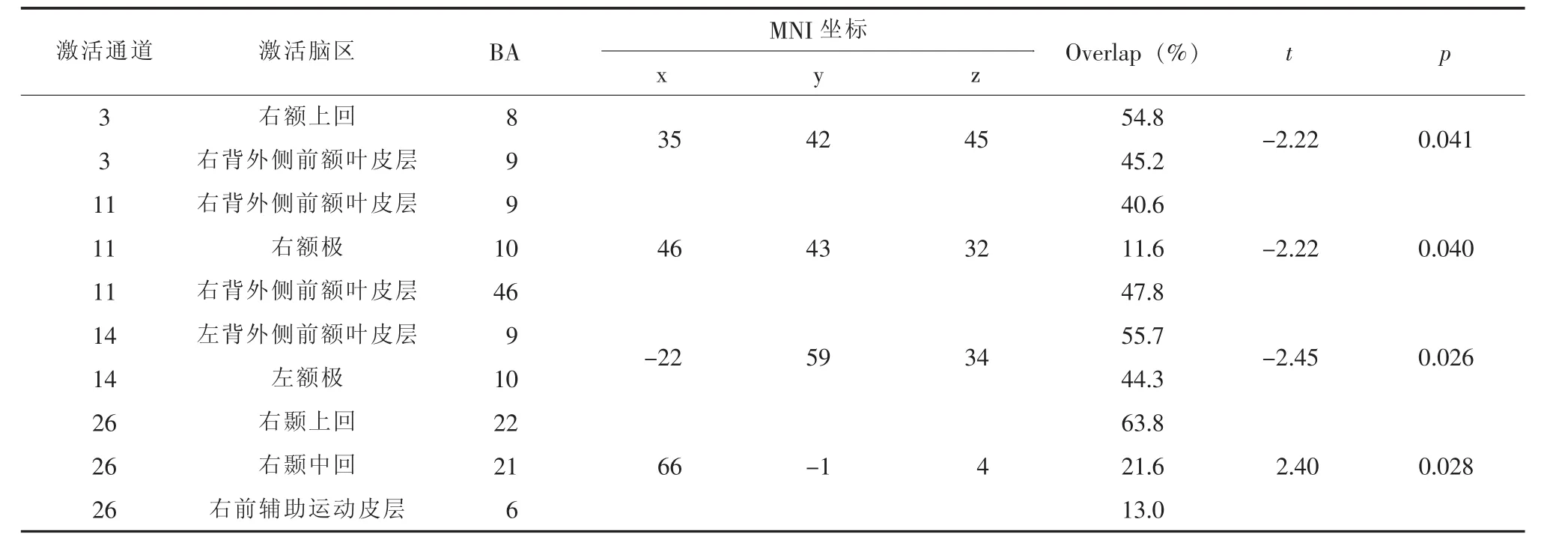
表3 高扩展vs.低扩展激活通道及对应脑区
以各个通道检验后的t值为激活程度的变化区间,基于 MATLAB程序绘制各个通道的 cluster map图谱,见图3。

图3 高扩展vs.低扩展大脑各个通道激活情形
3.3 人格元特质与概念扩展相关神经活动的关系
经统计分析发现,被试在人格可塑性和稳定性维度的平均得分分别为59.95±9.15,83.56±7.00。
将可塑性和稳定性维度得分分别与H条件下各个通道激活beta值进行斯皮尔曼相关分析。结果显示,可塑性维度与通道38覆盖脑区 (右额极(BA10)、右眶额区 (BA11))的活动显著正相关,r= 0.58,p=0.012,见图4;没有通道与稳定性维度相关。

图4 可塑性与脑区活动相关图
4 讨论
本研究基于概念扩展的核心特征,通过预先选定的两类 (高低扩展)用途刺激诱发的脑活动的对比,控制评估因素影响之后,进一步考察概念扩展的神经机制及其与人格特质的关系。可见,预先设置的二组材料间的概念扩展差异是本研究成立的前提条件。首先,预实验为二组实验材料的有效性提供了先期保障。更进一步,本研究的行为学数据显示,高扩展组的反应时显著长于低扩展组,在新颖性和联结难度评定等级方面,高扩展组也皆显著高于低扩展组,说明被试在进行该组概念扩展过程中需要跨越更大的 “意义距离”将目标刺激与报纸建立联结 (Mednick,1962)。这符合高扩展vs.低扩展联结难易评定等级差异性检验结果,即高扩展组的评定等级显著高于低扩展组。这也在统计学上验证了实验材料的有效性。同时,通过对材料新颖性和联结难易评定等级的相关分析,发现二者显著正相关。这与Acar和Runco(2014)的研究结果相一致。他们利用三种语义评定量表对新颖和普通客体用途与客体之间的反应间距进行了评定,发现随着客体用途新颖程度的增加,其与客体之间的语义间距呈扩大趋势。鉴于概念扩展的本质特征—扩展既有概念的界限以获取概念新含义,该结果进一步为实验材料的有效性提供了依据。
综合起来,本实验选定的高低扩展两组概念扩展刺激材料有着足够的扩展差距。通过对比两组材料诱发的脑功能活动,有可能分离出概念扩展的神经基础。
4.1 概念扩展相关脑功能活动分析
对任务诱发的近红外光学信号变化分析结果与预期假设基本一致。与低扩展条件相比,高扩展条件下右颞上回和颞中回 (BA21/22)表现出更大的激活,而双侧额极皮层 (BA10)和右背外侧前额皮层 (BA9)表现出更小的激活。可见,前额叶和颞叶等皮层均参与了概念扩展过程,但颞叶皮层表现为功能活动加强,而更多的前额叶皮层表现为功能活动减弱。
4.1.1 颞叶脑区在概念扩展过程中的作用
本研究结果与前人研究 (Abraham et al., 2012;Kröger et al.,2012,2013;Rutter et al., 2012a,2012b)一致,都表明右颞上回和颞中回参与了概念扩展。该脑区位于ATL(anterior temporal lobe)区。以往研究发现,ATL与某一背景下语义表征的联结和整合 (Lau et al.,2008),以及概念新知识的获取 (Hoffman et al.,2014)有关,是一个能够 “整合信息以形成新的连贯性概念”(Albright,2012;Hoffman et al.,2014;Rogers& McClland,2004)的脑区。尤其是右颞上回,它在整合较为弥散、粗糙的语义联结中扮演着激活远距离语义信息的角色 (Jung-Beeman et al.,2004; Kuperberg, Lakshmanan, Caplan, & Holcomb, 2006)。针对本研究而言,个体在以用途为目标导向下思考与报纸之间合理联结过程中,可能需要从报纸这一特定事物出发,努力突破其固有概念特征的界限,以新颖合理的方式整合其语义特征,实现其与呈现高扩展目标之间的联结,赋予报纸概念以新含义,从而完成对报纸概念的创造性操作。这一操作过程可能需要颞叶脑区具有的语义加工功能的支持,在神经层面表现为显著激活状态。
4.1.2 额极在概念扩展过程中的作用
与我们的预期相一致,实验没有观察到额极的显著正激活。先前研究显示额极皮层在概念扩展过程中显著激活 (Abraham et al.,2012;Kröger et al.,2012,2013;Rutter et al.,2012a,2012b),这可能是先前实验操纵的概念扩展过程含有评估过程所致。在先前探讨概念扩展神经机制的研究中,研究者依据被试 “尽可能的新颖性用途报告”或对客体-用途/隐喻新颖性和适用性的评估界定概念扩展,将概念扩展置于认知评估的前提之下。本实验则通过预试验从具有良好信效度的 《报纸用途测验》中筛选出高低二组材料,在统计学上对实验材料的新颖性和适用性预先进行了保障;其后,在实验操作过程中,要求被试思考材料刺激与报纸之间的合理联结,无需进行新颖性和适用性的评估。有研究曾指出 “当生成信息需要个体进行内在的自我评估时,额极脑区广泛参与其中” (Christoff& Gabrieli,2000)。因此,先前部分实验发现额极的广泛激活,更可能是基于认知评估前提下的扩展导致的,而非单一的扩展加工引起的。
另一方面,本研究中额极 (BA10)更小的激活结果对揭示概念扩展神经基础具有特殊提示,可能反映了对 “报纸”概念既有内在信息结构的打破而进行创造性扩展的过程。Green,Fugelsang,Kraemer,Shamosh和Dunbar(2006)指出额极在整合概念内在信息表征加工中显著激活。据此,就概念扩展而言,打破概念的既存界限以产生新颖含义联结的过程,可能需要解离概念的内在信息以发展新认识。这种与整合相对的解离概念既有内在信息的过程可能就表现为额极的去激活。
4.1.3 背外侧前额皮层在概念扩展过程中的作用
背外侧前额叶隶属于执行控制系统,在创造性思维中起着重要作用。它在创造性思维中的激活可能与创新观念生成后的评估有关。如 Ellamil,Dobson,Beeman和Christoff(2012)分离了创造性生成和评价两个过程考察创造性思维的脑激活模式。结果发现创造性生成更多地与内侧颞叶 (双侧海马、双侧海马旁回)的激活有关,而创造性评价与执行 (双侧背外侧前额叶、扣带回背前部)和默认网络的激活有关。Hao等 (2016)以脑电alpha波的同步为指标也发现额叶与观念生成时的评估关系密切。Fink等 (2017)也发现认知重评时前额叶alpha功率增加更为显著。如前所述,本研究观察到的背外侧前额激活弱化很可能与本实验设计的任务避免了"评估"过程有关。Liu等 (2015)的实验结果为这一解释提供了直接的证据。他们发现,背外侧前额在创造性思维中的激活呈现阶段性,观念生成时背外侧前额叶明显去激活但观念修正时重新变强;创造性水平高的作家前额叶的这种去激活更为显著,作者认为这可能反映了作家更有效地抑制了认知控制。
4.1.4 其他脑区在概念扩展过程中的作用
值得指出的是,研究还发现了眶额、额眼等脑区更小的激活现象,这可能有助于我们进一步完善对概念扩展乃至整个创造性神经机制的认识。
首先,对眶额区来说,一般认为其主要参与执行控制加工,负责抑制与执行任务无关的或不合适的信息加工 (Bryden&Roesch,2015;Jonker et al.,2015;Pu et al.,2015;Stalnaker,Cooch,& Schoenbaum,2015),如有研究指出眶额区的损伤会导致个体冲动选择、冒险性赌博等行为的发生(Bryden&Roesch,2015),而且其激活性的降低与冒险性赌博选择相关 (Jonker et al.,2015)。据此推测,眶额区可能参与个体认知加工过程中执行策略的选择和运用。在执行概念扩展任务过程中,个体面对不熟悉的新颖的联结目标时,可能更倾向于运用冒险性加工策略,以尽可能地联结目标用途与客体,促进创造性扩展的产生。这种加工策略的选择可能需要眶额区去激活化参与。
额眼区一般被认为其与注意有关 (Duan, 2014;Esterman et al.,2015)。Noudoost,Clark,和Moore(2014)曾指出,在注意加工过程中,“如果没有来自额眼功能区的信息输入,V4神经元将会搜集更多视觉方向的信息,从而相对降低对视觉目标特征的加工…额眼区的去激活能够显著增强非目标区对V4神经元活动的抑制效应”。说明额眼区对视觉客体目标信息的获取具有特殊作用,其去激活化有利于除视觉客体目标本身之外信息的加工。针对本研究的概念扩展任务,报纸实为潜在的扩展对象,视觉目标只是扩展联结的结果。因此,在进行联结加工时所利用的信息可能主要源于报纸这一潜在的背景性概念。这种较大权重地提取 “非目标”信息的加工,可能需要额眼区去激活的支持。该推测还需要后续实验更具有针对性的探讨。
4.2 脑激活模式与人格因素的关系
随着比大五人格更高层次的元特质概念的提出,研究者开始在该层次上思考人格与创造性认知的关系 (贾绪计,林崇德,2014)。元特质包括可塑性和稳定性两个维度,前者涵盖外倾性和经验开放性两个子维度,反映 “个体在认知和行为方面探索和参与新异事物的灵活性”;后者涵盖宜人性、尽责性和神经质三个子维度,反映 “个体在情绪、社会和动机方面保持稳定的倾向” (贾绪计,林崇德, 2014;Silvia et al.,2009)。结合前面被试执行概念扩展任务时,前额皮层脑区多呈现去激活以支持扩展的进行,如额极和眶额皮层去激活以支持个体更加灵活的选择合适的加工方式。若此推断具有合理性,那么本身就倾向于灵活加工信息的个体在大脑该部位的去激活程度应该相对较弱,即额极或眶额皮层脑区的活动与可塑性维度得分间正相关——随着个体可塑性的增强,概念扩展过程中额极或眶额皮层的去激活程度具有减弱的趋势。
本研究的相关分析证实了这一推测,发现可塑性维度与额极皮层、右眶额皮层显著正相关。但该结果和Li等人 (2014)的结构像研究结果在统计学上并不一致,他们发现 “外倾性与眶额皮层的灰质总量负相关”。但从功能性角度来看,是具有潜在一致性的。Li等人指出,外倾性 (特质创造性)越高,眶额灰质总量越低,越倾向于直觉性思维/冒险性加工。那么,当特质创造性高的个体在执行需要冒险性加工的任务时,眶额在结构上已经确立了这种加工方式的优势。在功能上可能就无需通过过度抑制 (眶额较低的激活与冒险性选择加工相关,Jonker et al.,2015)来满足上述加工方式的需要,即在功能上表现为特质创造性因素与相应脑区间的正相关。
综上,研究在神经层面为状态创造性和特质创造性皆为创造性不可或缺的方面提供了初步的证据,表明创造性的发生可能受到人格特征和执行任务的交互影响。至于二者之间具有怎样的交互效应,还需要后续更具针对性的实验探讨。
4.3 局限性
本实验的局限性在于没有控制高、低概念扩展组之间可能存在的任务难度差异。而功能脑区的激活水平随着任务难度的增加而增强的现象已被多个研究所证实 (Chee,Westpha,Goh,Graham,& Song,2003;Dräger et al.,2004;Gould,Brown, Owen, Ffytche, & Howard, 2003; Heuninckx, Wenderoth,Debaere,Peeters,& Swinnen,2005; Tregellas,Davalos,&Rojas,2006;Livesey,Wall, &Smith,2007)。因此,本实验难以排除任务难度在观察到的脑激活中的影响。但是,首先,这种增强不一定表现为其他任务不相关脑区的补偿性参与。例如,Gould等 (2003)的研究结果发现:随着任务难度增加,与任务相关的特定脑区的激活强度出现了增强,而不是激活了更多其他脑区。刘昌,翁旭初,李恩中,李德明和马林 (2005)的研究也提示任务难度的增加并不激活额外的脑区活动。其次,王大华,付艳,唐丹,段云云和于春水(2012)以汉语名词双字词对为材料,采用线索回忆任务,发现任务难度和大脑激活面积、强度之间均不存在对应的递增变化关系。最后,更值得注意的是,新近的两项同样使用近红外光学脑成像技术的研究结果提示本实验观察到的脑区激活与任务难度无关。潘津津,焦学军,姜劲,徐凤刚和杨涵钧(2014)以大学生为实验对象,考察了任务难度和大脑皮层血氧变化之间的关系,结果发现,无论是简单任务还是复杂任务,确实存在某些脑区含氧血红蛋白 (HbO)含量随任务难度提升而增加的现象,但对任务难度敏感的fNIRS信号出现在大脑前额叶皮层 (PFC)而非本实验观察到正激活的右颞中回等。潘津津等 (2015)的研究中分离了复杂任务的难度特征进一步考察大脑皮层血氧情况会随着任务难度增加而变化的现象,同样发现了前额叶激活水平与任务难度的相关性。他们发现,任务复杂度增加导致的任务难度变化主要改变前额叶含氧血红蛋白的响应程度,时间压力变化导致的任务难度变化与PFC、VC区域含氧血红蛋白的响应程度相关。MC区域含氧血红蛋白的响应程度对有无运动控制输出特征有较高的激活程度响应。综合起来,可以认为,在本实验中,与低概念扩展条件比较,高概念扩展条件下右颞中回、右辅助运动区表现出的正激活很可能反映了条件间概念扩展上的差异性而与任务难度无关。
5 结论
概念扩展过程需要颞叶和前额脑区的协同参与,其中颞叶脑区显著激活以支持新颖语义联结的建立;前额脑区去激活来支持概念既有界限的打破、合理高效加工方式的择取等认知加工;人格可塑性维度与额极皮层、右眶额皮层显著正相关。
贾绪计,林崇德.(2014).创造力研究:心理学领域的四种取向.北京师范大学学报 (社会科学版),(1),61-67.
刘昌,翁旭初,李恩中,李德明,马林.(2005).青老年组不同难度下心算活动的脑功能磁共振成像研究.心理科学,28(4), 845-848.
潘津津,焦学军,姜劲,徐凤刚,杨涵钧.(2014).利用功能性近红外光谱成像方法评估脑力负荷.光学学报,(11),336-341.
潘津津,焦学军,焦典,王春慧,徐凤刚,姜劲,张朕,曹勇(2015).利用功能性近红外光谱法研究大脑皮层血氧情况随任务特征变化规律.光学学报,(8),226-232.
王大华,付艳,唐丹,段云云,于春水.(2012).任务难度与老化对大脑激活的影响——以线索记忆编码为例.心理与行为研究, 10(1),25-31.
徐芝君,陈学志,邱发忠.(2012). 《报纸的不寻常用途》测验之编制.创造学刊,3(2),33-56.
朱小佳.(2014).大五人格量表 (BFI)的初步修订 (硕士学位论文).扬州大学.
Abraham,A.(2014).Creative thinking as orchestrated by semantic processing vs.cognitive control brain networks.Frontiers inHuman Neuroscience,8(1),95.
Abraham,A.,Pieritz,K.,Thybusch,K.,Rutter,B.,Kröger,S., Schweckendiek,J.,Stark,R.,Windmann,S.,&Hermann,C. (2012).Cre ativity and the brain:Uncovering the neural signature of conceptual expansion.Neuropsychologia,50(8), 1906-1917.
Acar,S.,&Runco,M.A.(2014).Assessing associative distance among ideas elicited by tests of divergent thinking.Creativity Research Journal,26(2),229-238.
Albright,T.D.(2012).On the perception of probable things: Neural substrates of associative memory, imagery, and perception.Neuron,74(2),227-245.
Amabile,T.(1996).Creativity in context.Westview press.
Boden,M.A.(1998).Creativity and artificial intelligence.Artificial Intelligence,103,347-356.
Bryden,D.W.,&Roesch,M.R.(2015).Executive control signals in orbitofrontal cortex during response inhibition.The Journal of Neuroscience,35(9),3903-3914.
Chee,M.W.L.,Westphal,C.,Goh,J.,Graham,S.,&Song,A.W. (2003).Word frequency and subsequent memory effects studied using event-related fMRI.Neuroimage,20(2),1042-1051.
Christoff,K.,&Gabrieli,J.D.(2000).The frontopolar cortex and human cognition:Evidence for a rostrocaudal hierarchical organization within the human prefrontal cortex.Psychobiology, 28(2),168-186.
Cui,X.,Bray,S.,&Reiss,A.L.(2010).Functional near infrared spectroscopy(NIRS) signalimprovementbased on negative correlation between oxygenated and deoxygenated hemoglobin dynamics.Neuroimage,49(4),3039-3046.
Dietrich,A.(2004).The cognitive neuroscience ofcreativity. Psychonomic Bulletin&Review,11(6),1011-1026.
Dräger,B.,Jansen,A.,Bruchmann,S.,Förster,A.F.,Pleger,B.,& Zwitserlood, P., et al. (2004) . How does the brain accommodate to increased task difficulty in word finding? Afunctional mri study.Neuroimage,23(3),1152-1160.
Duan,C.A.(2014).Beyond eye movements:A role for frontal eye fields in evaluating decisions.The Journal of Neuroscience,34 (24),8061-8062.
Ellamil,M.,Dobson,C.,Beeman,M.,& Christoff,K.(2012). Evaluative and generative modes of thought during the creative process.NeuroImage,59(2),1783-1794.
Esterman,M.,Liu,G.,Okabe,H.,Reagan,A.,Thai,M.,& DeGutis,J.(2015).Frontal eye field involvement in sustaining visual attention: Evidence from transcranial magnetic stimulation.Neuroimage,111,542-548.
Fink,A.,Weber,B.,Koschutnig,K.,Benedek,M.,Reishofer,G., Ebner,F.,Papousek,I.,&Weiss,E.M.(2014).Creativity and schizotypy from the neuroscience perspective.Cognitive, Affective,&Behavioral Neuroscience,14(1),378-387.
Fink,A.,Weiss,E.M.,Schwarzl,U.,Weber,H.,de Assun O,V. L.,Rominger,C.,Schulter,G.,Lackner,H.K.,&Papousek,I. (2017).Creative ways to well-being:Reappraisal inventiveness in the context of anger-evoking situations.Cognitive,Affective, &Behavioral Neuroscience,17(1),94-105.
Finke,R.A.,Ward,T.B.,& Smith,S.M.(1992).Creative cognition:Theory,research,and applications.Cambridge:MIT Press.
Gough,H.G.(1979).A creative personality scale for the Adjective Check List.Journal of Personality and Social Psychology,37 (8),1398.
Gould,R.L.,Brown,R.G.,Owen,A.M.,Ffytche,D.H.,& Howard,R.J.(2003).Fmri bold response to increasing task difficulty during successful paired associates learning. Neuroimage,20(20),1006-1019.
Green,A.E.,Fugelsang,J.A.,Kraemer,D.J.,Shamosh,N.A.,& Dunbar,K.N.(2006).Frontopolar cortex mediates abstract integration in analogy.Brain Research,1096(1),125-137.
Hao,N.,Ku,Y.,Liu,M.,Hu,Y.,Bodner,M.,Grabner,R.H.,& Fink,A.(2016).Reflection enhances creativity:Beneficial effects ofidea evaluation on idea generation.Brain and Cognition,103,30-37.
Heuninckx,S.,Wenderoth,N.,Debaere,F.,Peeters,R.,&Swinnen, S.P.(2005).Neuralbasisofaging:Thepenetration of cognition into action control.The Journal of Neuroscience,25 (29),6787-6796.
Hoffman,P.,Evans,G.A.,& Ralph,M.A.L.(2014).The anterior temporal lobes are critically involved in acquiring new conceptual knowledge:Evidence for impaired feature integration in semantic dementia.Cortex,50,19-31.
Jang,K.E.,Jeong,Y.,Ye,J.C.,Tak,S.,Jung,J.,&Jang,J. (2009).Wavelet minimum description length detrending for near-infrared spectroscopy.Journal of Biomedical Optics,14 (3),034004.
Jauk,E.,Neubauer,A.C.,Dunst,B.,Fink,A.,&Benedek,M. (2015).Gray matter correlates of creative potential:A latent variable voxel-based morphometry study.NeuroImage,111, 312-320.
Jonker,F.A.,Jonker,C.,Scheltens,P.,&Scherder,E.J.(2015).The role of the orbitofrontal cortex in cognition and behavior. Reviews in the Neurosciences,26(1),1-11.
Jung-Beeman,M.,Bowden,E.M.,Haberman,J.,Frymiare,J.L., Arambel-Liu,S.,Greenblatt,R.,Reber,P.J.,&Kounios,J. (2004).Neural activity when people solve verbal problems with insight.PLoS Biology,2(4),500-510.
Kozbelt,A.,Beghetto,R.A.,&Sternberg,R.J.(2010).Theories of creativity.In handbook of creativity.Cambridge:Cambridge University Press.
Kröger,S.,Rutter,B.,Stark,R.,Windmann,S.,Hermann,C.,& Abraham,A.(2012).Using a shoe as a plant pot:Neural correlatesofpassive conceptualexpansion.Brain Research, 1430,52-61.
Kröger,S.,Rutter,B.,Hill,H.,Windmann,S.,Hermann,C.,& Abraham,A.(2013).An ERP study ofpassive creative conceptualexpansion using a modified alternate usestask. Brain Research,1527,189-198.
Kuperberg,G.R.,Lakshmanan,B.M.,Caplan,D.N.,&Holcomb,P. J.(2006).Making sense of discourse:An fMRI study of causal inferencing across sentences.Neuroimage,33(1),343-361.
Lau,E.F.,Phillips,C.,&Poeppel,D.(2008).A cortical network for semantics:(de)constructing the N400.Nature Reviews. Neuroscience,9(12),920-933.doi:10.1038/nrn2532
Leutgeb,V.,Ille,R.,Wabnegger,A.,Schienle,A.,Schoggl,H., Weber,B.,et al.(2016).Creativity and borderline personality disorder:Evidence from a voxel-based morphometry study. Cognitive Neuropsychiatry,1-14.
Li,W.,Li,X.,Huang,L.,Kong,X.,Yang,W.,Wei,D.,et al. (2015).Brain structure links trait creativity to openness to experience.Social Cognitive&Affective Neuroscience,10(2), 191-198.
Liu,S.,Erkkinen,M.G.,Healey,M.L.,Xu,Y.,Swett,K.E.,Chow, H.M.,et al.(2015).Brain activity and connectivity during poetry composition:Toward a multidimensional model of the creative process.Human Brain Mapping,36(9),3351-3372.
Livesey,A.C.,Wall,M.B.,& Smith,A.T.(2007).Time perception:Manipulation oftask difficulty dissociatesclock functions from other cognitive demands.Neuropsychologia,45 (2),321-331.
Mednick,S.(1962).The associative basis of the creative process. Psychological Review,69(3),220.
Noudoost,B.,Clark,K.L.,& Moore,T.(2014).A distinct contribution of the frontal eye field to the visual representation of saccadic targets.The Journal of Neuroscience,34(10), 3687-3698.
Ozawa,S.,Matsuda,G.,& Hiraki,K.(2014).Negative emotion modulates prefrontal cortex activity during a working memory task: A NIRS study.Frontiers in Human Neuroscience,8(1),46.
Piffer,D.(2012).Can creativity be measured?An attempt to clarify the notion of creativity and general directions for future research.Thinking Skills and Creativity,7(3),258-264.
Pu,S.,Nakagome,K.,Yamada,T.,Yokoyama,K.,Matsumura,H., Yamada,S.,et al.(2015).Suicidal ideation is associated with reduced prefrontal activation during a verbal fluency task in patients with major depressive disorder.Journal of Affective Disorders,181,9-17.
Rhodes,M.(1961).An analysis of creativity.Phi Delta Kappan, 305-310.
Rogers,T.T.,&McClelland,J.L.(2004).Semantic cognition:A parallel distributed processing approach.MIT press.
Runco,M.A.(2014).Creativity:Theories and themes:Research, development,and practice.Elsevier.
Rutter,B.,Kröger,S.,Stark,R.,Schweckendiek,J.,Windmann,S., Hermann,C.,& Abraham,A.(2012).Can clouds dance? Neuralcorrelates ofpassive conceptualexpansion using a metaphor processing task:Implications for creative cognition. Brain and Cognition,78(2),114-122.
Rutter,B.,Kröger,S.,Hill,H.,Windmann,S.,Hermann,C.,& Abraham,A.(2012).Can clouds dance?Part 2:An ERP investigation of passive conceptual expansion. Brain and Cognition,80(3),301-310.
Sifonis,C.M.(1995).Scene schemas and creativity:Examining the influence of schema based knowledge on the creative process. Doctoral dissertation,Texas A&M University.
Silvia,P.J.,Nusbaum,E.C.,Berg,C.,Martin,C.,&O′Connor,A. (2009).Opennessto experience,plasticity,and creativity: Exploring lower-order,high-order,and interactive effects. Journal of Research in Personality,43(6),1087-1090.
Stalnaker,T.A.,Cooch,N.K.,&Schoenbaum,G.(2015).What the orbitofrontal cortex does not do.Nature Neuroscience,18 (5),620-627.
Sternberg,R.J.(1999).Handbook ofcreativity.Cambridge University Press.
Tregellas,J.R.,Davalos,D.B.,&Rojas,D.C.(2006).Effect of task difficulty on the functional anatomy of temporal processing. Neuroimage,32(1),307-315.
Activities of Prefrontal-temporal Regions and its Correlation with the Big Five Personality during Conceptual Expansion
Yin Desheng1,Ma Junpeng1,Yang Nan1,Jin Hua1,Zhu Chaozhe2,Lin Chongde2
(1 Key Research Base of Humanities and Social Sciences of the Ministry of Education,Academy of Psychology and Behavior,Tianjin Normal University,Tianjin 300074;2 National Key Laboratory of Cognitive Neuroscience and Learning,Beijing Normal University,Beijing 100875)
The study of the neural mechanism of Conceptual expansion has become a hotspot from a research perspective ofcrearivecognitive“process” approach.However,previousstudieshave failed to exclude the influence of evaluate effort on experiments.And no studies investigated whether brain activity during performing creative task is also relevant to personality.Here,participants were asked to view a use and to think out the reasonable connection,the reasonable operational way that can make paper implement the use,between the use and paper.According to material pre-experiment,the uses were separated into high-unusual and low-usual ones in advance.The prefrontal and temporal activities were recorded by functional near-infrared spectroscopy(fNIRS) during the experiment.To explore the correlations between neural activity related to conceptual expansion and personality,participants were asked to complete the Big Five Inventory.Results found decreased activity in dorsolateral prefrontal cortex(BA9,involved in default mode network)and frontopolar area(BA10)and increased activity in temporal regions(BA21,BA22)in the high as compared to the low expansion condition.In addition, personal meta-trait plasticity(including openness and extraversion)of Big Five Inventory was positively associated with activity in frontopolar areas and right orbitofrontal areas.Taken together,the findings indicate that temporal regions and prefrontal brain is involved in conceptual expansion and personal factors are related to frontal areas during performing creative expansion task.
creative processing,conceptual expansion,prefrontal,deactivation,meta-trait.
B842.1
2016-11-21
教育部哲学社会科学研究重大课题攻关项目 (11JZD040)、天津师范大学引进人才基金项目 (5RW108)和天津市教育科学 “十三五”规划课题项目 (HE1025)。
金 花,E-mail:jennyjin2@163.com。
猜你喜欢
浙江大学学报(理学版)(2021年1期)2021-01-26
养生月刊(2020年10期)2020-10-14
学生天地(2019年33期)2019-08-25
中学科技(2018年9期)2018-12-19
健康管理(2017年3期)2017-04-20
小雪花·初中高分作文(2016年5期)2016-05-14
中国医学科学院学报(2012年3期)2012-03-25
视野(2009年2期)2009-03-11
意林(2009年8期)2009-02-11
cookie world(2008年11期)2008-12-12
