
放牧和封育对青藏高原南缘亚高山草甸群落结构及物种多样性的影响
2017-09-13 05:19:26初晓辉单贵莲袁福锦尹海燕
草地学报 2017年5期
初晓辉, 谢 勇#, 单贵莲*, 袁福锦, 陈 功, 尹海燕
(1.云南农业大学草业科学系, 云南 昆明 650201; 2.云南省草地动物科学研究院, 云南 昆明 650212)
青藏高原南缘亚高山草甸不仅是当地藏民发展畜牧业的物质基础,也是滇西北乃至全国生态安全的重要屏障和多民族生存的重要家园。然而,过去几十年,由于气候变化和人类活动的干扰,导致青藏高原南缘亚高山草甸生产力水平下降、生物多样性减少、毒害草蔓延等问题日益突出[1]。
为应对青藏高原南缘亚高山草甸普遍退化所引发的一系列问题,近年来国家及相关部门相继启动了退牧还草、围封转移等草地恢复和治理项目,旨在恢复退化亚高山草甸的生态、生产和生计功能。其中全年封育、生长季封育+非生长季放牧、生长季封育+季末割草是上述生态工程常用的草地恢复和治理措施,被广泛应用。然而,上述封育措施能否有效促进滇西北退化亚高山草甸的恢复,哪种措施更有利于退化亚高山草甸“三生”功能的发挥。查阅相关文献,发现以往学者针对围封对草地恢复的有效性开展过一些研究,多数研究表明,退化草地围封后,由于消除了放牧扰动,以前因过度放牧而被抑制和消弱的种群的生长得以促进,草地生产力和盖度增加,群落物种组成及各物种优势度发生改变[2-6];物种丰富度和多样性增加[7-9]。也有研究指出,与适度放牧相比,长期围封不利于草地植被的恢复[6,10],也不利于群落中优良牧草种类的增加[7]。围封限制了牧草在放牧条件下具有的超补偿性生长机制的发挥,其大量的凋落物和立枯物影响资源的利用效率,因此降低了草地净初级生产力[11-12]。围封尤其是围封年限过长会降低物种多样性[3]和物种丰富度[13]。但上述研究主要是针对围封在恢复退化草地的有效性方面展开的,并且,这些研究主要集中在典型草原、荒漠草原和高寒草甸草原,对青藏高原南缘亚高山草甸,尚未开展过围封有效性及不同围封措施有效性方面的比较研究。
为探讨退化亚高山草甸围封恢复的有效性,筛选出亚高山草甸的适宜围封措施,本研究以青藏高
原南缘香格里拉县小中甸镇和平社区知特小组亚高山草甸的自由放牧样地、4年生长季围封+非生长季放牧样地、4年全年封育样地、以及20年生长季封育+季末割草样地为研究对象,开展放牧和封育对亚高山草甸群落结构及物种多样性影响的比较研究,研究成果可为亚高山草甸的合理利用与有效管理,生物多样性的保护及草地畜牧业的可持续发展提供理论指导。
1 材料与方法
1.1 研究区自然概况
研究区域位于云南省迪庆州香格里拉县小中甸镇,地处N 27°20′~27°43′,E 99°36′~99°59′,海拔3 200~3 400 m,年平均气温5.8°C,年平均降雨量646.9 mm,年蒸发量1 162.1 mm,年日照时数2 167.9 h,≥10℃年积温2 006.9℃,无霜期131~146 d。
研究区域土壤为亚高山草甸土和山地草甸土。植被类型为亚高山草甸,主要植物有草地早熟禾(Poapratensis)、小花剪股颖(Agrostismicrantha)、华扁穗草(Blysmussinocompressus)、大花嵩草(Kobresiamacrantha)、西南委陵菜(Potentillalineata)、车前(Plantagoasiatica)、西南鸢尾(Irisbulleyana)、大狼毒(Euphorbiajolkinii)、银莲花(Anemonecathayensis)等[1]。
1.2 研究对象
在研究区域选取和平社区支特小组相邻的自由放牧、4年生长季节封育+非生长季放牧、4年全年封育及20年生长季封育+季末割草样地为研究对象,研究对象地理位置、优势物种构成及利用与管理方式如表1所示。
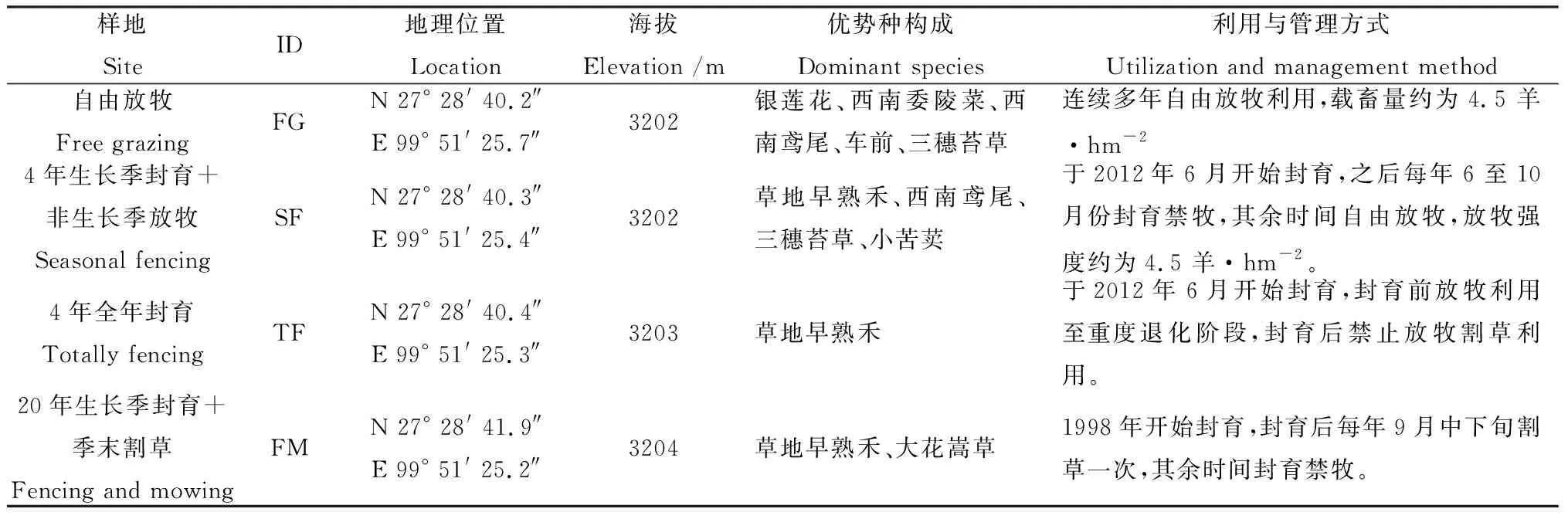
表1 研究样地地理位置与管理方式Table 1 Location and management method of experiment sites
1.3 研究方法
2015 年 7 月中旬植物生长高峰期,在试验样地内随机选取10个1 m ×1 m 的样方,测定记录植物种类、总盖度、各物种密度后,齐地面刈割,65℃烘干,分种测定地上现存量干重。
1.4 指标计算及数据分析
1.4.1物种优势度/重要值 体现种群在群落中地位和作用的综合数量指标,计算公式为:
IV=(Y′+D′)/2
式中IV为物种优势度(重要值),Y′为相对产量,D′为相对密度。
1.4.2物种多样性 体现群落在物种结构上的复杂程度及种群在群落中地位和作用的综合数量指标。包括物种丰富度、多样性、均匀度和生态优势度,分别采用Margalef丰富度指数(Ma)、Shannon-Weaver多样性指数(H)、Pielou均匀度指数(Epi)和Simpson优势度指数(D)表示,计算公式为:
Ma=(S-1)/lnN
H=-∑PiLnPi
Epi=H/LnS
D=∑(Ni/N)2
式中:S为物种数目;N为群落中所有物种个体总数;Pi=ni/N,ni为物种i的重要值;H为Shannon-Weaver指数;Ni为物种i的个体数。
1.5 数据分析
采用 SPSS 19.0 对群落植被特征进行单因素方差分析(One-way ANOVA),Duncan法进行多重比较。
2 结果与分析
2.1 放牧和封育对亚高山草甸群落物种组成的影响
表2为放牧和封育管理下亚高山草甸群落物种组成和各物种优势度的测定结果。由表2可知,在自由放牧样地,群落以西南鸢尾、西南委陵菜、银莲花为优势物种;放牧草地经4年的生长季围封+非生长季放牧管理,西南鸢尾、西南委陵菜、银莲花的优势地位显著下降,其优势度由18.60,18.07,13.73下降为11.62,8.56,1.73,草地早熟禾和三穗苔草的优势地位增加,由自由放牧样地的10.07,10.34增加为27.52、11.06,即SF样地群落以草地早熟禾、西南鸢尾、三穗苔草和西南委陵菜为优势物种;经4年的全年封育管理,群落中草地早熟禾的优势地位继续增加,由自由放牧样地的10.07增加为70.07,即TF样地以草地早熟禾为单优势物种;经20年的生长季封育+季末割草管理,群落以草地早熟禾和大花嵩草的优势度最高,分别为51.77和23.20,即FM样地以草地早熟禾和大花嵩草为优势物种。
2.2 放牧和封育对亚高山草甸主要物种优势地位的影响
由图1可看出,自由放牧草地采取封育管理措施后,西南委陵菜、西南鸢尾、银莲花及三穗苔草的优势地位逐渐衰减,这些物种为退化亚高山草甸恢复过程中的典型衰退型种群;草地早熟禾种群采取封育管理措施后优势地位迅速增加,该物种是亚高山草甸封育恢复过程中的优势增长型种群;大花嵩草在封育割草管理下会演替为群落中的优势物种,原因是大花嵩草属莎草科嵩草属低矮多年生草本,常年割草有利于低矮草本的生长。
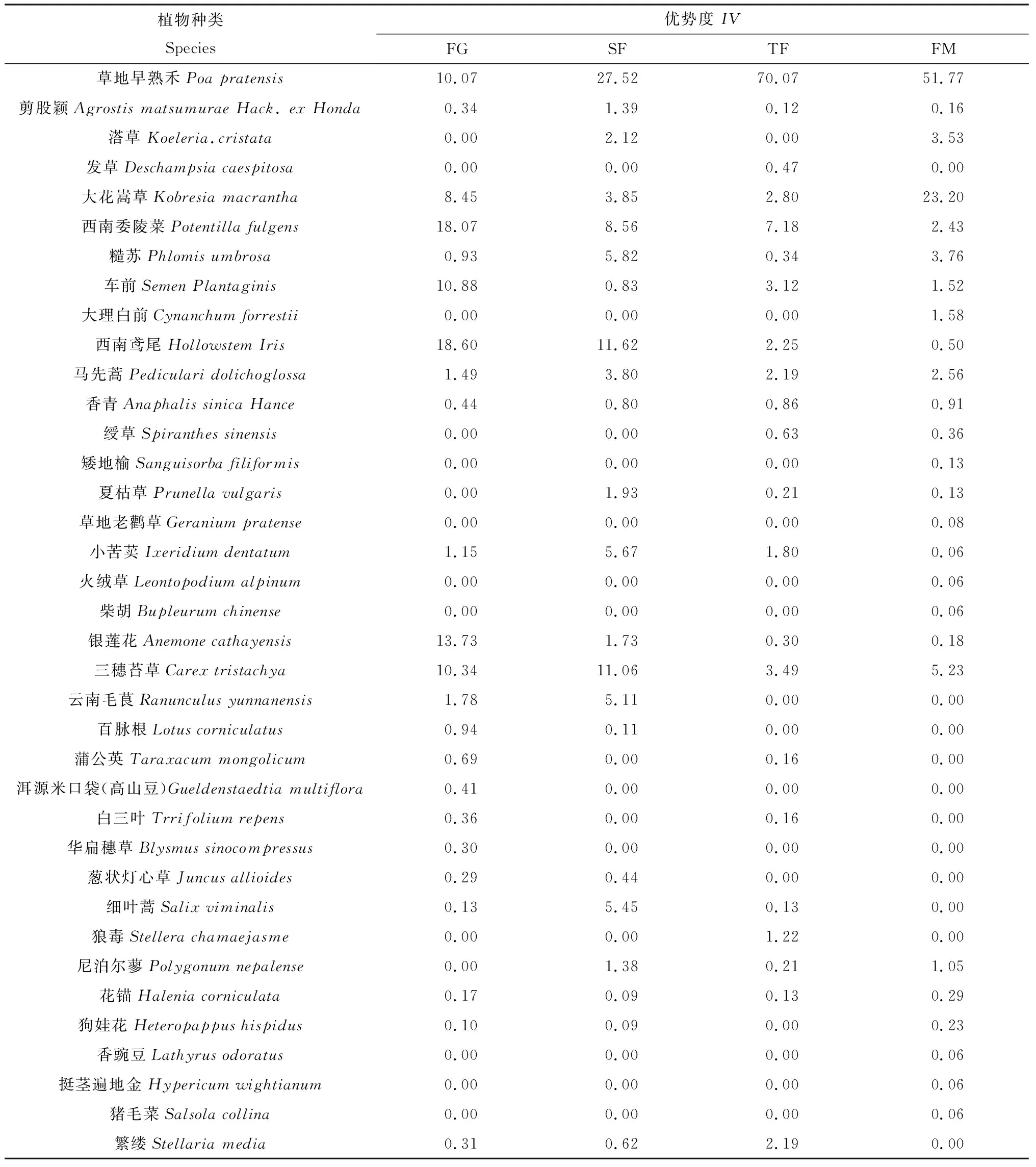
表2 放牧和封育对亚高山草甸群落物种组成及各物种优势度的影响Table 2 Effects of grazing and fencing on species composition and important values of subalpine meadow
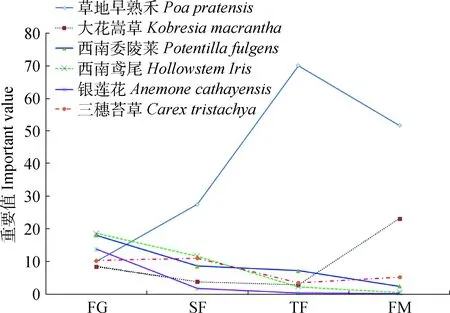
图1 放牧和封育对亚高山草甸主要物种优势度的影响Fig.1 Effect of grazing and fencing on important valuesof main species in subalpine meadow
2.3 放牧和封育对亚高山草甸地上现存量、盖度和密度的影响
图2为放牧和封育管理下亚高山草甸群落地上现存量、盖度和密度的测定结果。由图2可知,放牧和封育管理下亚高山草甸的地上现存量的高低顺序为TF>FW>SF>FG(P<0.05);盖度和密度以TF和FM显著高于SF和FG(P<0.05)。表明退化自由放牧草地经4年的生长季封育+非生长季放牧管理(SF),其地上现存量、盖度和密度无显著性增加;经4年的完全封育(TF)和20年的生长季封育+季末割草(FM)管理,其群落地上现存量、盖度和密度均显著增加,与自由放牧样地间差异显著(P<0.05)。
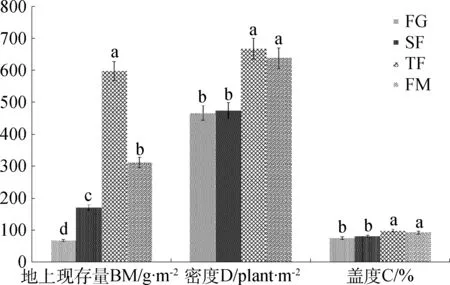
图2 放牧和封育对亚高山草甸群落地上现存量、盖度和密度的影响Fig.2 Effect of grazing and fencing on above-groundbiomass, coverage and density of subalpine meadow
2.4 放牧和封育对亚高山草甸物种多样性的影响
放牧和封育管理下亚高山草甸物种丰富度、多样性、均匀度和生态优势度的动态变化如图3、4所示。由图 3、4可知:自由放牧草地经4年的生长季封育+非生长季放牧(SF)管理,草地中物种的丰富度指数(Ma)和生态优势度(D)指数显著下降(P<0.05),多样性(H)和均匀度(Epi)指数变化不显著;经 4 年的全年封育(TF)管理,群落以单个物种(草地早熟禾)占绝对优势(单优势物种群落),群落生态优势度(D)急剧增加,而丰富度(Ma)、多样性(H)和均匀度(Epi)指数显著降低,与自由放牧草地差异显著(P<0.05);经20年的生长季封育+季末割草(FM)管理,群落丰富度指数(Ma)增加,但由于群落为寡优势物种群落,群落生态优势度指数(D)显著增加,多样性(H)和均匀度(Epi)指数显著降低(P<0.05)。

图3 放牧和封育对亚高山草甸物种丰富度和多样性的影响Fig.3 Effect of grazing and fencing on speciesrichness and diversity of subalpine meadow
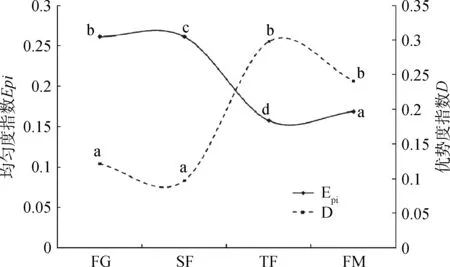
图4 放牧和封育对亚高山草甸群落均匀度和生态优势度的影响Fig.4 Effect of grazing and fencing on speciesevenness and dominance of subalpine meadow
3 讨论
3.1 放牧和封育对草地物种组成和群落结构的影响
关于放牧和封育对草地物种组成和群落结构的影响,多位学者研究指出,适度放牧可刺激草本牧草分蘖和灌木分枝,促进牧草再生,提高草地光合作用和固碳能力,但过度放牧则会导致草地退化[14-15]。重牧退化草地封育后,由于消除了放牧扰动,植被得以恢复,草地生产力、植被盖度增加,群落物种组成及各物种优势度发生改变[2-7,16-20],但恢复效果与封育年限密切相关[18,21-22]。如Wu等[18]研究表明,与过牧草地相比,围封3年、8年、20年和24年的羊草草原,其地上生物量分别增加71%,162%,167%和174%;苗福泓等[21]研究表明,青藏高原高寒草甸地上生物量随封育时间延长表现为先显著增加后显著降低,封育第4年和第5年地上生物量最大,封育第6年,草地初级生产力开始下降。单贵莲等[22]研究指出,与重度退化自由放牧草地相比,围封7年、10年、13年、20年,典型草原地上生物量和0~30 cm根系生物量先增加,封育13年左右达最大,之后下降。也有研究指出,封育限制了牧草在放牧条件下具有的超补偿性生长机制的发挥,其大量的凋落物和立枯物降低了植物生产的周转率,影响资源的利用效率,因此降低了草地净初级生产力[11-12]。本研究结果显示,与自由放牧相比,青藏高原南缘退化亚高山草甸经4年的生长季封育+非生长季放牧(SF)管理,草地中西南委陵菜、西南鸢尾、银莲花等有毒有害植物的优势地位降低,早地早熟禾等优良牧草的优势地位增加,但草地的生产力、密度和盖度无显著增加;经4年的全年封育(TF)和20年的生长季封育+季末割草(FM)管理,草地生产力、密度、盖度及优良禾本科牧草的优势度显著增加,说明封育能有效促进退化亚高山草甸植被的恢复,但恢复效果与封育措施及封育年限密切相关,与前人研究结论一致[2-7,16-22]。
3.2 放牧和封育对草地物种多样性的影响
关于放牧和封育对草地物种多样性的影响,有研究表明,适度放牧利用,草地处在“中度干扰”阶段,中度干扰下群落优势种的竞争力消弱从而使稀有种增加[23],或者干扰产生多种生境斑块从而使不同演替阶段的种共存[24],因此适度放牧有利于草地物种多样性的增加,但过度放牧往往导致草地中伴生种减少,群落物种多样性降低[25-26]。退化草地采取封育管理措施后,由于外界干扰源的消除和生境的改善,为群落中减少的和一些已消退的物种再度“侵入”创造了条件,因此物种丰富度和多样性增加[7-9,22]。也有研究指出,封育尤其是封育年限过长会降低物种多样性,如Xiong等[3]研究指出,6年的封育降低了青藏高原北部高山草甸的物种多样性;Altesor等[11]研究显示,在乌拉圭温带草地,由于中度放牧导致暖季生长的植物取代了一些冷季生长的杂草,所以中度放牧地的其物种多样性显著高于封育9 年的草地。Abella等[13]研究指出,与中度放牧相比,封育减少了莫哈韦沙地群落中本地物种的数量,因而降低其物种丰富度;Matthew等[27]指出,封育因排除了干扰使生境趋于均一化而导致生物多样性的降低。本研究结果显示,与自由放牧(FG)相比,青藏高原南缘退化亚高山草甸经4年的生长季封育+非生长季放牧(SF)管理,草地中物种的丰富度显著下降,多样性、均匀度变化不大;经 4 年的完全封育(TF)管理,草地的丰富度、多样性和均匀度显著降低;经20年的生长季封育+季末割草(FM)管理,群落的丰富度指数增加,但由于群落为寡优势物种群落,群落的多样性和均匀度显著低于自由放牧草地。概括而言,与放牧利用相比,青藏高原南缘亚高山草甸采用封育管理措施后,草地的丰富度、均匀度和多样性降低,与前人研究结论一致[3,11,13,27]。
4 结论
与自由放牧利用相比,生长季封育+非生长季放牧、全年封育、生长季封育+季末割草管理均可增加滇西北退化亚高山草甸优良牧草草地早熟禾、大花嵩草、三穗苔草等的优势度,降低草地中西南委陵菜、西南鸢尾、银莲花等有毒有害植物的优势地位。同时,与自由放牧利用相比,封育后草地生产力、密度、盖度增加,但物种丰富度、均匀度和多样性降低,说明封育可促进退化亚高山草甸植被的恢复,但封育后因排除了放牧干扰,群落中单个或少数物种在群落中的优势地位大大增加,草地中伴生种减少,群落物种丰富度和多样性降低。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
绿色科技(2021年10期)2021-06-23 03:13:06
活力(2019年21期)2019-04-01 12:17:10
草业学报(2019年2期)2019-02-25 01:57:16
绿色科技(2017年6期)2017-04-20 08:13:04
河北林业科技(2016年5期)2016-11-08 03:13:30
