
氮和磷添加对草原群落特征及有关生态过程的影响
2017-09-13 05:19王金枝姜丽丽李博文崔树娟孟凡栋汪诗平
草地学报 2017年5期
王 奇, 王金枝, 姜丽丽, 周 阳, 李博文, 崔树娟, 孟凡栋, 汪诗平
(1. 中国科学院青藏高原研究所, 北京 100101; 2. 中国科学院大学, 北京 100049; 3. 中国林业科学研究院湿地研究所 湿地生态功能与恢复北京市重点实验室, 北京 100091;4. 中国科学院青藏高原地球科学创新卓越中心, 北京100101)
草地是我国重要的陆地生态系统,占国土总面积的41%左右。由于气候变化和不合理放牧等原因导致了天然草地不同程度的退化。因此,如何维持较高的生物多样性和提高草地植物生产力,已成为各级政府和科研人员普遍关心的问题。许多研究表明,影响植物生产力的因素很多,在较大的地理尺度上,气候等环境因子(如气温、降水和土壤类型等)是决定植物生产力的关键因子[1-2],而在较小的地理单元上,生物和资源有效性等可能是植物生产力大小的主导因子[3]。因此,植物生产力受到环境和生物因子互作的影响,其各自影响程度依据不同草地生态系统状态而异。有学者从其他角度综述了影响草地生产力的主要因素[4],本文拟从养分添加的角度扼要综述氮(N)和磷(P)添加对植物多样性和物种组成影响,特别是P添加对主要植物和群落N利用形态的生态位分离以及利用效率的影响,并从植物根系的形态和周转对N和P添加的响应等方面,系统分析了N、P添加对草地植物生产力影响的过程和机理。在此基础上,提出可能存在的问题并提出未来的研究重点,为退化草地生态系统的恢复和草地生产力的提高提供借鉴。
1 N和P添加对物种组成及生产力的影响
大多数研究表明N添加能够提高草地生产力[5-8]。对于高寒草甸而言,有些研究发现N添加提高了高寒草甸生产力[9-11],但也有研究显示N添加并没有提高其生产力[12],或者N添加仅仅提高了植物地上净初级生产力而对植物总的生产力没有显著影响[13-14], 只有在N、P同时添加后植物生产力才显著增加[14]。这些结果表明不同生态系统的第一限制因子可能不同,高寒草甸植物的生长极有可能更受土壤中可利用P含量的限制,或者是N和P共同限制的[14]。但是N和P哪个是高寒草甸植物生长的第一限制因子目前尚不清楚。
N添加后群落中一些植物能够快速对新增加的外源养分做出反应,通过直接或间接的资源获取和后代的繁殖再生而排斥其它物种,从而成为群落的主要优势种,进而影响生态系统生产力[15]。例如,在高寒草甸生态系统中添加N后,禾草类植物生物量的增加显著高于杂类草植物[16-17],表明不同植物对N添加的响应不尽相同。或者N和P同时添加显著降低了青藏高原亚高寒草甸生态系统的物种多样性,单独P添加对各功能群物种丰富度和总丰富度无显著性影响[7],表明P添加可能影响了植物N的吸收利用。而内蒙古典型草原一、二年生杂类草随N添加增加显著,但多年生禾草生物量却随N添加显著下降[18]或者多年生禾草占优势的群落转变为半灌木占优势的群落[19-20],但是当N和P同时添加时,仅菊科植物生物量显著增加[21]。这表明植物物种对N和P添加的响应不同,甚至同一功能群的植物(例如禾草)在不同生态系统中对N和P添加的响应也不尽相同。植物物种对N和P的需求量以及N利用策略不同可能是导致植物对N和P添加响应不同的原因[19,22-23]。但是N和P同时添加如何影响植物多样性和群落物种组成进而影响生产力形成的过程和机制目前还不十分清楚。
2 N和P添加对植物N利用策略的影响
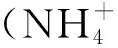
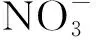
3 N和P添加对植物叶片N、P含量及其回收量的影响
陆地生态系统的多个结构和功能特征是受N和P限制的[9,41],因此植物叶片N和P含量对于基本生态学过程有重要的预示作用[42],如初级生产力[9,41]、凋落物分解[43]以及土壤营养的可利用性变化等[44-45]。特别是植物N和P含量与植物代谢过程是紧密耦合在一起的[46],用植物N和P的含量可以表征植物的N和P利用效率,同时,利用成熟叶片N和P含量与立枯中N和P的含量可以估算植物N和P的回收率[47]。提高养分利用效率,以最小的养分吸收量或最大的养分回收量来生产最多的新生物量是植物适应贫瘠环境的重要策略之一[47]。研究发现大尺度上气候因素对植物N∶P的影响仅为2%,系统发生学决定高寒植物叶片N∶P的大小[48-49]。同样,吕晓涛等也在内蒙古典型草原发现植物叶片,甚至非叶器官(例如茎)的N含量和回收量在不同物种和功能群之间存在较大的差异[50-52]。在高寒草甸发现高的N、P回收效率可能正是莎草在群落组成中占优势的原因[11]。吕晓涛等对内蒙古典型草原不同物种植物N、P含量和在枯落物中的回收量对N和P添加及放牧的响应做了比较细致的研究,发现N和P添加增加优势植物羊草(Leymuschinensis)和大针茅(Stipagrandis)的绿叶和枯落叶N含量,对P含量没影响,降低了植物的N和P回收量,而P添加却对植物N和P含量均没有显著影响[50-53]。有研究发现P添加对叶片N浓度的影响会在不同物种间表现出差异,如Milla等[54]发现,Pistacialentiscus叶片N含量在P肥沃的生境和P贫瘠的生境之间无显著差异;Kozovits等[55]则发现,施P肥可以提高Blepharocalyxsalicifolius叶片N浓度。另有学者发现在低P水平下,绿叶N浓度也与施P量呈正相关[56]。
邓建明等指出高寒草甸植物N素吸收与土壤N素相关性不显著[57],而宾振钧等[58]却发现高寒草甸N添加导致5种优势植物叶片N含量增加,但是黄花棘豆的N含量不受影响,并且随着N添加,高寒草甸优势植物叶片N∶P升高,意味着优势物种可能更多地受到P的限制[58]。可见,高寒草甸优势植物叶片N和P含量低于我国草地植物N和P含量的平均值的前提下,植物叶片N和P含量以及回收量对N和P添加的反应与典型草原很可能不一致,但这方面的研究还很少。
4 N和P添加对植物根系形态与周转的影响
植物根系是构成生态系统生产力的重要部分,在植物营养吸收的过程中起着决定性的作用[59]。根系形态(根长、根表面积、根直径等)和周转(寿命)的变化与土壤养分密切相关[23,60]。草地植物较粗的根系(直径≥0.5 mm的根系)对养分的吸收贡献不大,而细根(直径≤0.2 mm的根系)在吸收土壤养分的过程中起到了重要作用,并且会随着土壤养分浓度的变化而改变[60-62]。由于土壤限制了根系的可观察性,以及刻画根系形态性状的科学量化方法不完善等因素,使得根系研究相对困难[63]。国内关于N和P添加对根的形态的影响研究目前多集中在农田[23,60,64]或典型草原植物[19],高寒草甸生态系统中植物根的形态研究仅在自然条件下做了初步探讨[59]。早期的研究发现N添加降低了植物细根生产[65-66]。郭大立和范萍萍[62]推测N添加后,土壤中植物可利用性N增加,植物细根有机会吸收更多的N,细根寿命延长,但生物量并没有增加,甚至还会减少。另有研究发现农田P添加却能通过调控植物细根的产生,改变植物的根表面积和长度,减缓根的枯萎等方式提高植物生产力[64]。
5 存在的问题与建议
目前国内外关于施肥对草地生产力、群落结构和多样性等方面已有大量研究,而高寒草甸由于有机质矿化率低,再加上长期过度放牧,致使土壤养分供给不足,已成为限制植物生长的关键因子[67]。如嵩草草甸地处青藏高原的高海拔地区,尽管土壤中储存着大量的有机氮(N)和磷(P),由于受低温限制,有机N和P矿化作用比较弱,因此可利用N、P含量仅占土壤全N、全P含量的1%左右,植物的生长极有可能受到可利用N和P的限制[68]。由于人类活动所引起的气候变化包括全球气温上升、N沉降增加等,青藏高原近年来正经历着明显高于全球平均水平的温暖化过程,土壤温度的升高促进了土壤中微生物的活性,同时N沉降有逐年增加的趋势,这些环境变化均促使土壤中可利用营养元素增加[14,69-70],很有可能改变高寒草甸的物种组成和生物量以及其他相关生态过程,进而可能对正在发生的气候变化产生巨大的反馈作用。目前对高寒草甸很多研究证明不同植物对N可利用形态存在明显的生态位分化[22,30-31],且施N肥对其产生了显著影响[36]。杨晓霞等研究发现N、P添加均促进青藏高原高寒草甸植物生物量的增加,同时P的促进作用要高于N,表明土壤有效P对植被的限制作用可能大于N的限制作用[14]。
N添加对不同草地生态系统植物生产力的影响不尽相同,甚至只有同时添加N、P时植物生产力才显著提高[14],说明不同草地生态系统植物生产力的第一限制因子可能不同,特别是对于高寒草甸生态系统而言,目前尚不清楚N和P哪个是高寒草甸植物生产力的首要限制因子。同时,植物细根生产决定了植被地下净初级生产力[59],早期的研究发现N、P添加改变了细根的寿命、根面积、长度及其周转速率[65-66]。当土壤N含量发生变化时,植物可能通过改变N的利用形态从而影响其在群落中的优势度,进而改变群落物种组成并影响生态系统生产力,但是土壤N含量变化后导致的P限制的变化或者N和P交互作用如何影响上述有关过程的研究却很少。青藏高原由于特殊的地理位置,对西北乃至整个中国的生态环境都有极大的影响[71]。而高寒草甸作为青藏高原的主要草场,由于近几十年来气候变化以及不合理的土地利用方式导致大片草地严重退化[72],因此,对于草地生态系统,特别是高寒草甸生态系统而言,建议优先开展或加强以下过程和机理的研究:
5.1 N和P添加对草地植物生产力的相对限制作用及其互作效应
有研究发现只有N和P同时添加才可显著提高高寒草甸植物生产力,或P添加增强了N的作用效果[6-7,14],表明高寒草甸可能是P限制或者N和P共同限制的。所以,开展N和P添加对草地生产力相对影响及其潜在互作效应的研究,将有利于回答不同草地生态系统N、P对生产力的第一限制性作用及其主要影响因素等科学问题,从而为生产实践提供科学依据。
5.2 N和P添加对植物根系形态与周转的影响
郭大立和范萍萍推测N添加细根寿命延长可能是植物地下生物量维持的机制[62]。栽培试验发现P添加可增加植物侧根生长[64]。表明植物细根对N和P添加的响应方式可能不同,N和P添加可通过改变植物细根的形态和周转从而影响生态系统生产力,然而这方面的研究相当缺乏。由于草地生态系统地下生物量占总生物量的60%~90%,该研究的缺乏影响了我们对N、P添加对植物生产的地下过程的了解,制约了通过N、P添加提高草地生态系统生产力的认识。
5.3 N和P添加如何通过影响植物对N的利用形态而影响群落组成和生态系统生产力
许多研究表明草地植物不同物种的N利用形态(铵态氮、硝态氮和氨基酸小分子有机氮)不同,并且植物的N利用形态因季节和土壤剖面不同而异。然而,关于P添加如何影响N添加的效应,以及如何影响植物的N利用形态的研究相对较少,进一步制约了N和P添加如何通过影响植物N形态利用及时空变化从而改变植物在群落中的优势度,从而影响植物物种组成和生态系统生产力的理解。
5.4 N和P添加对植物N、P利用效率或回收效率的影响
植物叶片的N、P含量和回收量在不同植物物种和功能群之间存在较大的差异。然而,对于这些的影响过程和机理尚不清楚,特别是N和P添加条件下植物如何通过改变N和P利用效率或回收效率进而影响植物物种组成和生产力的过程和机理有待进一步研究。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
河南科学(2020年3期)2020-06-02
活力(2019年21期)2019-04-01
红土地(2016年3期)2017-01-15
河北林业科技(2016年5期)2016-11-08
草业科学(2015年9期)2015-12-11
环球市场信息导报(2015年15期)2015-12-03
