
N-糖基化对β-甘露聚糖酶在毕赤酵母中异源表达的影响
2017-09-03 10:06姜风超马立娟杜丽平肖冬光
食品科学 2017年16期
马 清,蔡 瑞,姜风超,马立娟,2,*,杜丽平,2,肖冬光,2
N-糖基化对β-甘露聚糖酶在毕赤酵母中异源表达的影响
马 清1,蔡 瑞1,姜风超1,马立娟1,2,*,杜丽平1,2,肖冬光1,2
(1.天津科技大学生物工程学院,工业发酵微生物教育部重点实验室,天津 300457;2.天津食品安全低碳制造协同创新中心,天津 300457)
为研究N-糖基化对里氏木霉β-甘露聚糖酶(β-mannanase,Man1)在毕赤酵母GS115中异源表达的影响,采用定点突变的方法,将Man1上3 个N-糖基化修饰位点(N131、N158和N329)上的天冬酰胺用中性谷氨酰胺取代。结果发现,N-糖基化位点的突变对Man1的转录水平无明显影响,突变后获得的Man1表观分子质量有轻微下降,与Man1相比,3 个突变体N131、N158和N329的甘露聚糖酶活力分别降低了85.43%和79.48%和16.3%;而热稳定性分别提高了7.87%、13.5%和15.37%。由此可见,N-糖基化修饰对于β-甘露聚糖酶的高效表达是必需的,其中N131和N158糖基化位点尤为重要,但是会稍微降低β-甘露聚糖酶的热稳定性。
β-甘露聚糖酶;N-糖基化;定点突变;酶活力;热稳定性
β-甘露聚糖酶(β-1,4-mannanase,EC3.2.1.78)能够水解β-1,4-D-甘露糖苷键连接的葡萄甘露聚糖、甘露聚糖、半乳葡萄甘露聚糖及半乳甘露聚糖等底物[1],是一种仅次于木聚糖酶的第二大半纤维素酶类,在饲料、食品、造纸、制药等各个领域均具有广阔的应用前景[2-6],近年来对于它的研究也逐渐成为了国内外的研究热点。β-甘露聚糖酶在自然界的来源比较广泛,包括动物[7]、植物[8-10]和微生物[11-12],其中主要的来源是微生物。由于微生物具有生长周期短、易于培养和产酶能力强等优点[13],其已成为工业化生产β-甘露聚糖酶的主要“加工厂”。里氏木霉(Trichoderma reesei)作为生产纤维素酶与半纤维素酶的优良菌种[14],目前对其分泌的β-甘露聚糖酶Man1在毕赤酵母(Pichia pastoris)中表达的研究不多。
毕赤酵母表达系统作为一种高效的外源蛋白真核表达系统,通常存在蛋白质的糖基化修饰[15],N-糖基化是真核生物所特有的糖基化修饰。N-糖基化修饰是β-构型的N-乙酰葡糖胺异头C原子与天冬酰胺(Asn)的γ-酰胺N原子共价连接而成的N-糖苷键的修饰[16]。β-甘露聚糖酶在毕赤酵母中表达的主要翻译后修饰为N-糖基化修饰,氨基酸序列Asn-Xaa-Ser/Thr(Xaa是除了脯氨酸的任意氨基酸、Ser为丝氨酸、Thr为苏氨酸)被认为是N-糖基化的特征序列[17]。研究表明,N-糖基化修饰既可增加也可减少糖蛋白的稳定性,这取决于糖基化修饰发生的位置,当糖基化修饰发生在折叠有序的区域时,会减少其稳定性,而当糖基化修饰发生在混乱无序的位置时则会増加蛋白质的稳定性[18]。例如,Guo Meijin等[19]研究发现毕赤酵母重组肌醇六磷酸酶在90 ℃条件下处理10 min可维持40%的酶活力,而去糖基化后在40 ℃条件下处理10 min酶活力剧烈下降;Xi Hongxing等[20]也发现N-糖基化修饰的亮氨酸氨基肽酶具有较高的热稳定性和催化效率。此外,Wang Changqing等[21]的研究也表明糖蛋白的去N-糖基化修饰虽然不会改变蛋白质的二级结构,但会降低蛋白质热失活过程的可逆性,导致热变性过程中蛋白质的聚集,这也进一步阐明了N-糖基化修饰可提高蛋白质稳定性的原因。但是,也有文献报道糖基化修饰不会显著影响酶的热稳定性甚至会使其热稳定性降低。齐飞飞[22]发现里氏木霉纤维二糖水解酶Ⅰ的不同位点上的糖基化对酶分子的热稳定性的影响不同,N384位的糖基化能显著提高酶的热稳定性,N45和N270位点的糖基化对热稳定性无显著影响。Han Minghai等[23]通过定点突变用Asn-Xaa-Thr取代了Asn-Xaa-Ser氨基酸序列,增强了重组弹性蛋白酶的N-糖基化程度,结果发现N212和N280位点突变使弹性蛋白酶活力分别提高了43%和25%,而N36位点突变则降低了31%。而Vaquero等[24]发现在酯酶成熟肽的N-糖基化修饰对酶的活性没有任何影响。谢春芳等[25]利用生物信息学方法涉及并引入N-糖基化突变位点对假蜜环菌来源的β-甘露聚糖酶进行了分子改造,发现N-糖基化对该酶的热稳定性、酸碱稳定性和蛋白酶抗性均有改善。因此,有必要研究N-糖基化修饰对β-甘露聚糖酶在毕赤酵母中表达的影响,以便进一步优化重组β-甘露聚糖酶在毕赤酵母中的高效表达,满足其在各个领域的应用。
本研究采用NetNGlyc 1.0 Server软件在线分析里氏木霉β-甘露聚糖酶Man1中易发生N-糖基化修饰的位点,利用定点突变技术,将里氏木霉β-甘露聚糖酶N-糖基化修饰位点上的Asn用中性谷氨酰胺(Gln)取代。比较突变体表达的酶与非突变体表达的酶的转录水平,酶活力水平和热稳定性等特性,进而探讨N-糖基化对里氏木霉β-甘露聚糖酶在毕赤酵母中异源表达的影响,为β-甘露聚糖酶的定向改造提供理论依据从而有利于对其进一步研究。
1 材料与方法
1.1 菌株与载体
里氏木霉(T. reesei Rut)C30、大肠杆菌(Escherichia coli)DH5α、毕赤酵母GS115由天津科技大学天津市工业微生物重点实验室保藏;重组载体pPIC9K-Man1为本实验室构建并保存,其中Man1基因来源于里氏木霉。
1.2 试剂与培养基
3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)国药集团化学试剂有限公司;刺槐豆胶、十二磺基磺酸钠 美国Sigma公司;考马斯亮蓝R-250、山梨醇、生物素、无氨基酵母氮源(yeast nitrogen base without amino acids,yNB) 北京Solarbio公司;柠檬酸、柠檬酸钠、磷酸氢二钾、磷酸二氢钾、甲醇、甘油 天津市化学试剂一厂;咪唑 天津市化学试剂三厂;D-甘露糖 上海蓝季科技发展有限公司;酵母提取物、蛋白胨 英国Oxoid公司;聚合酶链式反应(polymerase chain reaction,PCR)试剂、限制性内切酶、Marker大连宝生物公司。
LB培养基:蛋白胨1%、酵母提取物0.5%、NaCl 1%;yPD培养基:酵母提取物1%、蛋白胨2%、葡萄糖2%;BMGy培养基:酵母提取物1%、蛋白胨2%、yNB 1.34%、甘油1%、生物素4×10-5%;BMMy培养基:酵母提取物1%、蛋白胨2%、yNB 1.34%、甲醇0.5%、生物素4×10-5%。
1.3 仪器与设备
HE-120多功能水平电泳槽、HE-120多功能垂直电泳槽 上海天能科技公司;hS-1300-V型超净台 苏州安泰空气技术有限公司;PCT-200型PCR基因扩增仪美国Bio-Rad公司;全自动凝胶成像仪 美国Syngene公司;H1650-W台式高速离心机 湖南湘仪实验室仪器开发有限公司;5430R型4 ℃离心机 德国艾本德股份公司;H2Q-C摇床 哈尔滨市东明医疗仪器厂;ECM830电转化仪 美国BTX公司;红外激光成像系统 美国LI-COR公司;StepOne型荧光定量PCR(quantitative real-time PCR,qRT-PCR)仪 美国ABI公司;DK-8D型电热恒温水槽 上海一恒科技有限公司。
1.4 方法
1.4.1 软件分析β-甘露聚糖酶Man1中易发生N-糖基化的位点
采用NetNGlyc 1.0 Server软件在线分析里氏木霉β-甘露聚糖酶Man1中易发生N-糖基化修饰的位点并选择其中阈值超过0.5的位点进行下一步研究。
1.4.2 突变载体的构建
利用定点突变试剂盒将N-糖基化位点上编码天冬酰胺Asn的位点(AAT或AAC)突变为编码中性Gln的位点(CAA)。以大肠杆菌扩增后的重组质粒pPIC9K-Man1为模板,通过上游引物MF和下游引物MR对质粒进行扩增。本实验所用引物见表1,均利用oligo7软件设计并由金唯智公司合成。PCR扩增程序为:95 ℃预变性30 s;95 ℃变性15 s,60 ℃退火15 s,72 ℃延伸9 min,30 个循环;72 ℃终延伸5 min。取扩增产物40~50 μL加入1 μL DpnⅠ,混匀后在37 ℃恒温反应1~2 h,以去除模板质粒,用琼脂糖凝胶电泳检测消化后的产物并进行切胶回收。

表1 PCR引物序列Table 1 Sequences of primers used in this study
重组反应体系(20 μL)为:ddH2O 2 μL、5×CEⅡ Buffer 4 μL、DpnⅠ消化产物12 μL、ExnaseⅡ 2 μL。将各组分混匀后置于37 ℃水浴锅中反应30 min,反应结束后立即置于冰浴中冷却5 min,所得到的产物即为突变质粒。用得到的质粒转化大肠杆菌DH5α感受态,用含有氨苄的LB培养基筛选阳性克隆,挑取阳性单克隆到液体LB培养基中,37 ℃、200 r/min摇床培养过夜后提取质粒并进行测序,将测序结果与pPIC9KMan1比对以确定是否突变成功。
1.4.3 毕赤酵母重组菌株的构建与筛选
将经过测序后确认构建成功的突变载体经限制性内切酶SacⅠ线性化后各取20 μL与80 μL毕赤酵母GS115感受态混合,混匀后,转入冰预冷的0.2 cm电转杯,冰上放置5 min后电击(1 500 V、25 μF、200 Ω、6 ms)转化,结束后迅速加入1 mL冰预冷的无菌1 mol/L山梨醇至杯中,于超净台内将所有溶液转至灭菌离心管中,30 ℃条件下静置培养1 h后取200 μL涂布于MD平板,30 ℃培养48~72 h至单克隆产生。
挑取MD板转化子依次点于含不同质量浓度G418(1、2、4 mg/mL)的yPD培养基平板上,30 ℃分别培养48 h,以筛选多拷贝转化子。将筛选到的阳性转化子挑取到5 mL yPD液体培养基中,30 ℃、240 r/min培养16~18 h后提取重组酵母基因组DNA进一步鉴定。
1.4.4 甲醇诱导培养
挑取筛选得到并验证正确的高拷贝单克隆,接种至含25 mL BMGy的250 mL摇瓶中,30 ℃、240 r/min培养至OD600nm为2~6(约16~18 h)。用灭菌离心管,4 ℃、3 000 r/min离心5 min收集细胞,用BMMy重悬细胞至OD600nm为1.0进行诱导。每24 h取样,4 ℃、5 000 r/min离心5 min,收集上清液进行酶活力测定。为补偿甲醇的挥发损失,每24 h补加甲醇至体积分数为0.5%,直至达到最佳的诱导时间。
1.4.5 突变体转录水平的测定
本实验通过qRT-PCR方法比较3 种重组突变体与重组pPIC9K-Man1产甘露聚糖酶转录水平的差异。
将3 种重组突变体与重组pPIC9K-Man1在同一条件下进行甲醇诱导培养,72 h后提取重组菌体,用RNA提取试剂盒和反转录试剂盒分别提取4 种菌体的RNA并反转录为cDNA,以此为模板通过qRT-PCR进行定量比较。
qRT-PCR体系(20 μL)为:SyBR Premix Ex TaqⅡ(2×)10 μL、10 μmol M F 0.8 μL、10 μmol M R 0.8 μL、50×ROX reference dye 0.4 μL、cDNA 2 μL、H2O 6 μL。以3-磷酸甘油醛脱氢酶基因GAP为内参基因,不加入模板为阴性对照以排除其他成分污染的影响,重组pPIC9KMan1(甲醇诱导2 h的甘露聚糖酶)表达量为阳性对照,即定义为1。所有的PCR样品进行3 次重复,重复水平的方差作为误差。PCR扩增程序为:95 ℃预变性3 min;95 ℃延伸30 s,60 ℃ 20 s,40 个循环;程序循环95 ℃ 15 s,60 ℃ 15 s,95 ℃ 15 s。
1.4.6 突变体酶活力的测定
将3 种重组突变体与重组pPIC9K-Man1在同一条件下进行甲醇诱导培养,每隔24 h补甲醇至0.5%并取样,5 000 r/min离心5 min取上清液作为粗酶液。用DNS法[26]测酶活力:取诱导上清液200 μL与1.8 mL底物0.5%刺槐豆胶溶液(50 mmol/L pH 5.0的柠檬酸钠缓冲液配制)于试管中混匀,以200 μL柠檬酸钠缓冲液作空白对照,置于60 ℃水浴反应5 min,流水冷却,加入3 mL DNS溶液,加热煮沸5 min。冷却后在540 nm波长处测吸光度,以D-甘露糖标准溶液做标准曲线,酶活力定义:在pH 5.0、60 ℃的条件下,以每分钟生成相当于1 μmol/L D-甘露糖所需的酶量为一个酶活力单位,用IU/mL表示。
1.4.7 突变体和非突变体重组甘露聚糖酶的纯化
在诱导168 h后,将4 种重组体的发酵液4 ℃、5 000 r/min离心5 min取上清液并用Ni-Agarose His标签蛋白纯化试剂盒纯化蛋白质。将上清液用0.22 μm滤膜过滤后加载到柱上,用含有pH 7.9 Tris-HCl和0.5 mmol/L NaCl的不同浓度咪唑溶液进行梯度洗脱,洗脱梯度为40、60、80、100、200、300、500 mmol/L。最后得到的纯化样品置于8~14 kD透析膜中用50 mmol/L柠檬酸缓冲液进行透析以置换出咪唑。将得到的蛋白质存于4 ℃条件下进行后续分析。
取部分纯化后的样品通过十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)[27]进行分析。
1.4.8 突变体热稳定性的测定
将纯化的4 种甘露聚糖酶适当稀释后,分别在70 ℃保温15、30、45、60 min,用DNS法测定3 种突变体和重组pPIC9K-Man1表达的β-甘露聚糖酶Man1的剩余酶活力,以最高甘露聚糖酶活力为100%,其他条件下的酶活力占最高酶活力的百分比为该酶在温度处理后的相对酶活力。
2 结果与分析
2.1 糖基化位点的预测
用NetNGlyc 1.0 Server软件在线分析里氏木霉β-甘露聚糖酶Man1中易发生N-糖基化修饰的位点。由图1、表2可知,Man1中共有5 个天然N-糖基化位点(Asn-Xaa-Ser/ Thr),其中阈值超过0.5的分别是N131、N158和N329,对其进一步研究。
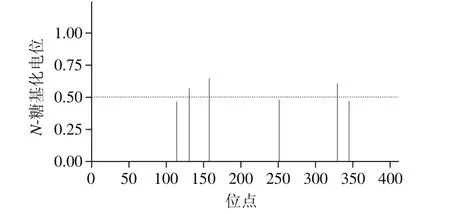
图1 N-糖基化位点预测Fig. 1 Predicted N-glycosylation sites
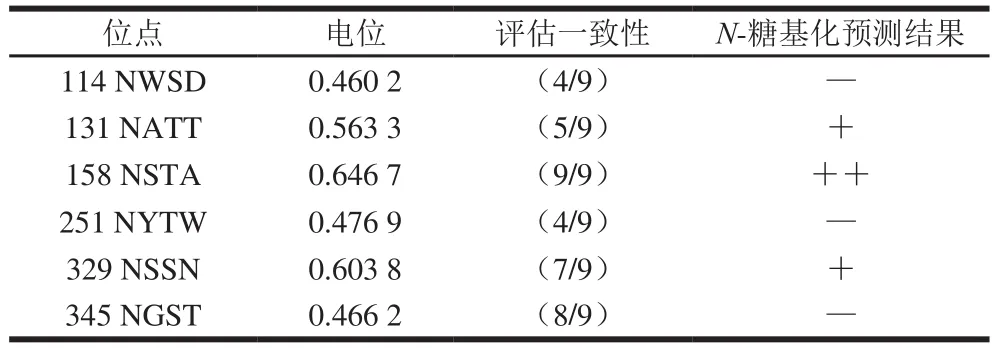
表2 Man1序列N-糖基化位点预测结果Table 2 Predicted N-glycosylation sites of Man1
2.2 突变载体的构建与鉴定
2.2.1 突变载体的构建
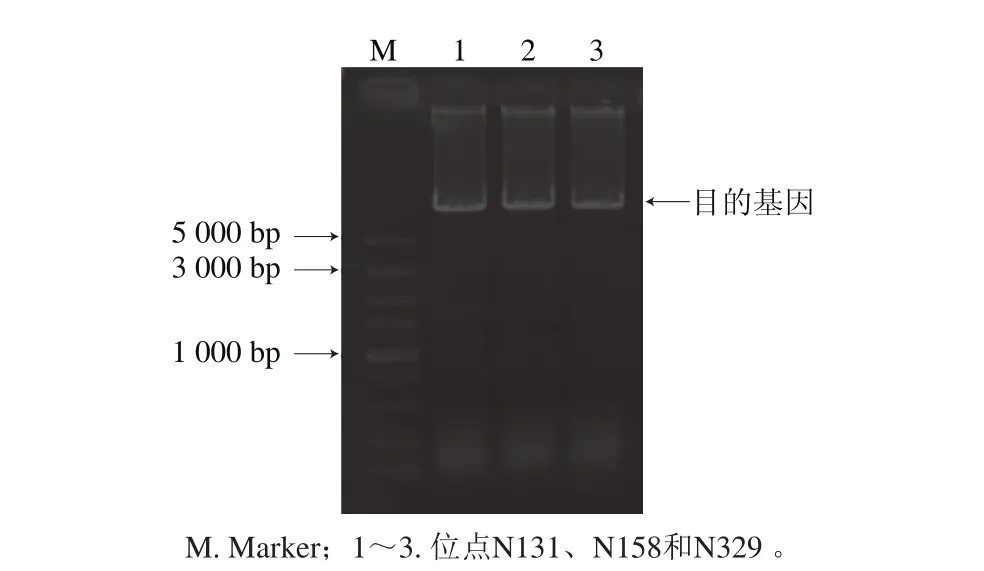
图2 定点突变PCR扩增Fig. 2 Agarose gel electrophoresis of PCR amplified products of mutant genes obtained by site-directed mutation
以重组pPIC9K-Man1为模板进行PCR扩增后得到线性的目的质粒,由图2可知,条带1~3可能是所需的目的基因片段。然后通过琼脂糖凝胶回收试剂盒对目的条带进行纯化回收后,用DpnⅠ消化去除甲基化模板pPIC9K-Man1,用ExnaseⅡ催化使目的基因环化,即得到3 种突变重组质粒pPIC9K-N131、pPIC9K-N158和pPIC9K-N329。经大肠杆菌DH5α扩增后的重组质粒pPIC9K-Man1是经过甲基化修饰的,而PCR扩增产生的DNA片段不带甲基化修饰,DpnⅠ酶的识别位点被甲基化后才能被识别切割,即DpnⅠ酶只能识别甲基化的模板链而不能识别PCR产物链,因此通过DpnⅠ酶消化就能把模板DNA链和新合成的PCR产物链区分开[28]。然后在ExnaseⅡ催化下扩增产物5′末端和3′末端由于存在15~21 bp反向互补区域可以发生同源重组,完成扩增产物的环化过程,即得到了环状的突变重组载体。
2.2.2 重组质粒的验证
用构建成功的3 种突变重组载体转化大肠杆菌DH5α感受态,经氨苄抗性筛选后提取阳性转化子,挑取单克隆培养并提取质粒后将质粒进行测序。测序结果与pPIC9KMan1序列进行比对,结果如图3所示。N-糖基化修饰的特征氨基酸序列Asn-Xaa-Ser/Thr中编码Asn的位点(AAT或AAC)均已突变为编码中性的谷氨酰胺的位点(CAA),其他序列均与模板保持一致,这一结果说明3 个糖基化位点均已成功突变,3 种突变重组载体已构建成功。
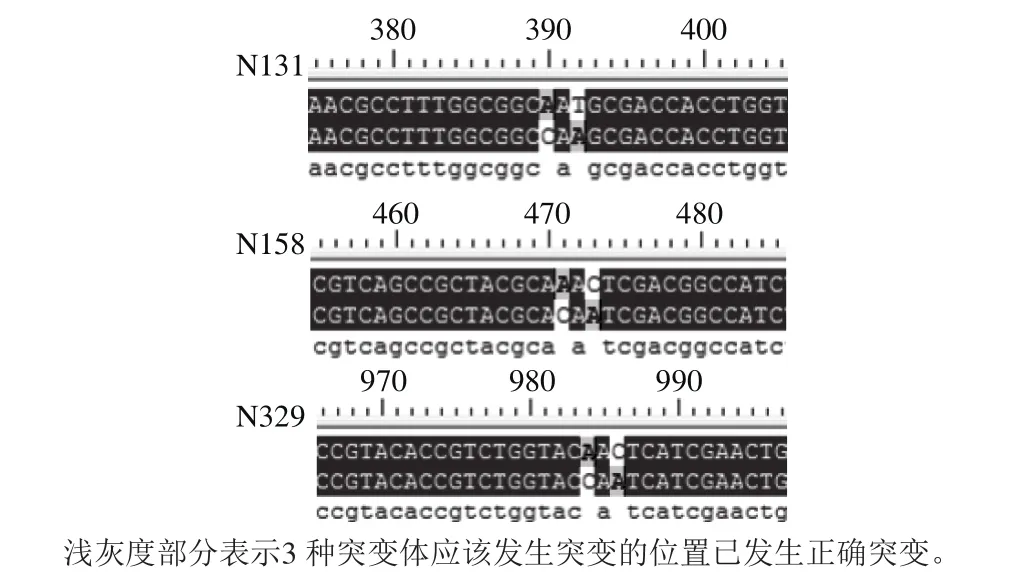
图3 突变序列的比对Fig. 3 Sequence alignment between mutant strains and pPIC9K-Man1
2.3 3 种重组突变体与重组Man1的比较
2.3.1 转录水平的比较
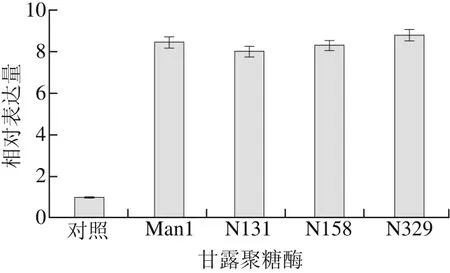
图4 甘露聚糖酶转录水平的比较Fig. 4 Comparison of transcriptional expression levels of recombinant mannanases
通过进行qRT-PCR比较3 种毕赤酵母重组突变体与重组Man1在甲醇诱导72 h后产甘露聚糖酶转录水平的差异,结果如图4所示。4 种重组菌体甘露聚糖酶的转录水平无明显差异,却均比重组Man1甲醇诱导2 h后产甘露聚糖酶转录水平高7~9 倍,说明N-糖基化修饰对重组甘露聚糖酶的转录水平表达无明显影响,而甲醇诱导时间对重组甘露聚糖酶的影响较大。
2.3.2 酶活力水平的比较
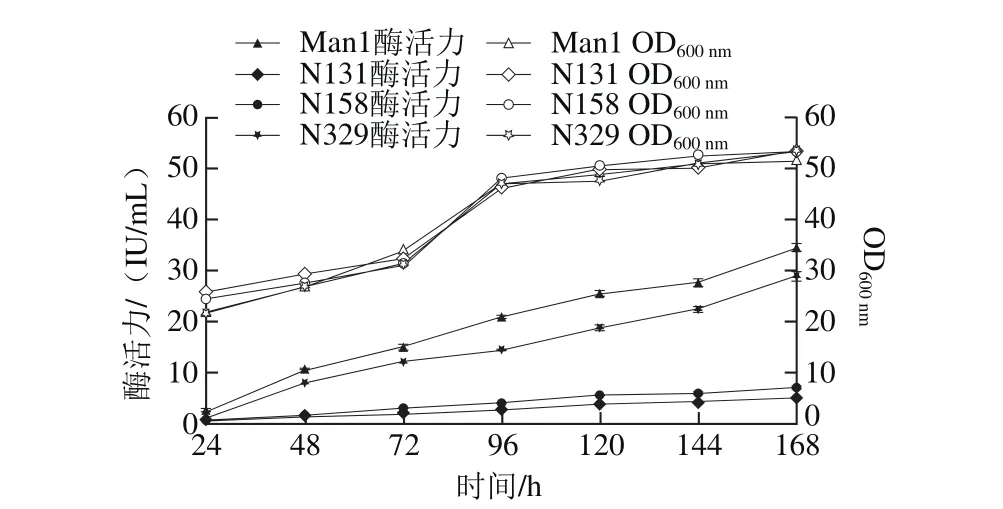
图5 4 种重组体产甘露聚糖酶活力和菌体生长曲线Fig. 5 Time-courses for mannanase production and the growth of recombinant strains
将筛选正确的3 种毕赤酵母重组突变体pPIC9K-N131、pPIC9K-N158和pPIC9K-N329与重组pPIC9K-Man1在同等条件下进行甲醇诱导,结果如图5所示。随着诱导时间的延长,甘露聚糖酶活力和菌体OD600nm也逐渐升高,然而在4 种重组体的菌体量与上清液中总蛋白质量浓度(1.2 mg/mL左右)均无显著差异的条件下,重组甘露聚糖酶活力却有很大不同,即去N-糖基化修饰的3 种突变体分泌的甘露聚糖酶活力与重组Man1相比均有所下降,其中N131、N158分泌的重组酶活力下降幅度比较大,分别为85.43%、79.48%,而N329与Man1相比酶活力下降幅度不大,仅为16.3%。突变后造成酶活力降低的原因可能是因为糖基化修饰可以通过改变蛋白质的整体构象从而影响构象决定的所有功能,如蛋白质的正确折叠,如果糖基化受到抑制,那么蛋白质不能正确折叠因而达不到应有的功能状态[29]。
2.3.3 纯化后的SDS-PAGE分析
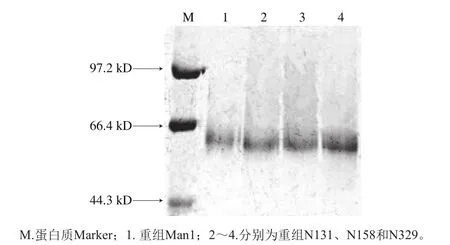
图6 突变体和原重组酶的SDS-PAGE分析Fig. 6 SDS-PAGE analysis of recombinant mannanases
pPIC9K-N131、pPIC9K-N158和pPIC9K-N329与重组pPIC9K-Man1分泌的4 种β-甘露聚糖酶经镍柱纯化后,进行进一步SDS-PAGE分析,结果如图6所示,4 种重组甘露聚糖酶均呈单一条带,再一次验证了3 种毕赤酵母突变体已成功构建并成功表达了目的蛋白。3 种重组突变体与重组pPIC9K-Man1表达的蛋白质相比表观分子质量略有下降,这可能是因为毕赤酵母分泌的蛋白质大多数为N-连接糖基化高甘露糖型,蛋白质转录后会一定程度上增加寡糖链长度。而增加的长度约为平均每个支链8~14 个甘露糖残基,即去N-糖基化修饰后分子质量约降低1~3 kD[30]。因此从图中可以看出3 种突变体与原重组体Man1相比,表观分子质量仅有轻微的下降。
2.3.4 热稳定性的比较
将3 种重组突变体和原重组体Man1在70 ℃分别处理不同时间,DNS法测相对酶活力,结果如图7所示,4 种重组甘露聚糖酶(Man1、N131、N158和N329)的相对酶活力均随处理时间的延长而剧烈下降,处理60 min后分别下降至13.82%、21.69%、27.32%和29.19%。而与原重组体Man1相比,3 种突变体(N131、N158和N329)热稳定性分别提高了7.87%、13.5%和15.37%。酶在某一温度条件下的半衰期是指将酶在某个温度保存不同时间,测其在不同时间的剩余酶活力,当剩余酶活力为初始酶活力50%时对应的时间。从图7可知,4 种重组甘露聚糖酶(Man1、N131、N158和N329)在70 ℃的半衰期的大小顺序为:N329>N158>Man1>N131,但是在相对酶活力下降到50%之后Man1的下降速率明显高于N131,所以1 h后Man1的剩余活力最小。由此可见,这3 个位点的N-糖基化位点突变可以在一定程度上增强毕赤酵母中异源表达的β-甘露聚糖酶的热稳定性,即这3 个位点的糖基化会降低该重组酶的热稳定性,这可能是因为这3 个位点的糖基化影响了酶的局部结构的改变,使其变为相对不耐热的结构,因而降低了它的热稳定性。
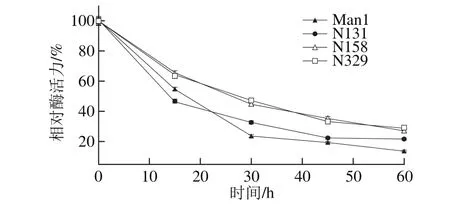
图7 突变体和原重组酶的热稳定性比较Fig. 7 Comparison of thermal stability of recombinant mannanases
3 结 论
本实验成功构建了毕赤酵母突变体N131、N158和N329,并且将它们所表达的β-甘露聚糖酶在表观分子质量、酶活力、转录水平表达及热稳定性等方面与重组Man1分泌的甘露聚糖酶进行了比较,结果发现,3 种突变重组甘露聚糖酶均能在毕赤酵母GS115中成功表达,且3 种突变体分泌的甘露聚糖酶纯化后测得的表观分子质量因其位点突变而有轻微的下降。突变后重组甘露聚糖酶在转录水平没发生明显变化,而在同一条件下3 种突变体(N131、N158和N329)的酶活力与重组Man1相比分别下降了85.43%、79.48%和16.3%,但热稳定性却分别提高了7.87%、13.5%和15.37%。可见N-糖基化修饰会在一定程度上增强毕赤酵母中异源表达的β-甘露聚糖酶活力,轻微降低它的稳定性,使表观分子质量轻微增大,对酶转录水平表达无明显影响。
[1] STÅLBRAND H, SIIKA-AHO M, TENKANEN M, et al. Purification and characterization of two β-mannanases from Trichoderma reesei[J]. Journal of Biotechnology, 1993, 29(3): 229-242. DOI:10.1016/0168-1656(93)90055-R.
[2] CAI H y, SHI P J, LUO H y, et al. Acidic β-mannanase from Penicillium pinophilum C1: cloning, characterization and assessment of its potential for animal feed application[J]. Journal of Bioscience and Bioengineering, 2011, 112(6): 551-557. DOI:10.1016/j.jbiosc.2011.08.018.
[3] PAN X, ZHOU J G, TIAN A, et al. High level expression of a truncated β-mannanase from alkaliphilic Bacillus sp. N16-5 in Kluyveromyces cicerisporus[J]. Biotechnology Letters, 2011, 33(3): 565-570. DOI:10.1007/s10529-010-0457-8.
[4] VÁRNAI A, HUIKKO L, PERE J, et al. Synergistic action of xylanase and mannanase improves the total hydrolysis of softwood[J]. Bioresource Technology, 2011, 102(19): 9096-9104. DOI:10.1016/ j.biortech.2011.06.059.
[5] CHAUHAN P S, PURI N, SHARMA P, et al. Mannanases: microbial sources, production, properties and potential biotechnological applications[J]. Applied Microbiology Biotechnology, 2012, 93(5): 1817-1830. DOI:10.1007/s00253-012-3887-5.
[6] VAN ZyL W H, ROSE S H, TROLLOPE K, et al. Fungal β-mannanases: mannan hydrolysis, heterologous production and biotechnological applications[J]. Process Biochemistry, 2010, 45(8): 1203-1213. DOI:10.1016/j.procbio.2010.05.011.
[7] ZAHURA U A, RAHMAN M M, INOUE A, et al. An endo-β-1,4-mannanase, AkMan, from the common sea hare Aplysia kurodai[J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2010, 157(1): 137-143. DOI:10.1016/ j.cbpb.2010.05.012.
[8] PRAKASH R, JOHNSTON S L, BOLDINGH H L, et al. Mannans in tomato fruit are not depolymerized during ripening despite the presence of endo-β-mannanase[J]. Journal of Plant Physiology, 2012, 169(12): 1125-1133. DOI:10.1016/j.jplph.2012.03.017.
[9] AGUILERA-GÁLVEZ C, VÁSQUEZ-OSPINA J J, GUTIÉRREZSANCHEZ P, et al. Cloning and biochemical characterization of an endo-β-1,4-mannanase from the coffee berry borer Hypothenemus hampei[J]. BMC Research Notes, 2013, 6: 333. DOI:10.1186/1756-0500-6-333.
[10] 周海燕, 赵文魁, 吴永尧. 甘露聚糖酶的多样性分析[J]. 生物技术通报, 2008(2): 60-67.
[11] 陈小兵, 乔宇, 丁宏标, 等. 里氏木霉RUT-C30内切β-甘露聚糖酶基因在毕赤酵母中分泌表达[J]. 农业生物技术学报, 2007, 15(1): 142-146.
[12] 高宏斌. 里氏木霉RUT-C30 β-甘露聚糖酶的诱导表达及其cDNA的克隆[D]. 呼和浩特: 内蒙古农业大学, 2003.
[13] 李剑芳. 黑曲霉酸性β-甘露聚糖酶及制备魔芋低聚糖研究[D].无锡: 江南大学, 2007.
[14] 郭雨刚. 毕赤酵母表达系统及其应用[D]. 合肥: 中国科学技术大学, 2012.
[15] 韩铭海. 弹性蛋白酶在毕赤酵母中表达及N-糖基化位点突变对重组酶的影响[D]. 无锡: 江南大学, 2013.
[16] KOWARIK M, NUMAO S, FELDMAN M F, et al. N-linked glycosylation of folded protein by the bacterial oligosaccharyltransferase[J]. Science, 2006, 314: 1148-1150. DOI:10.1126/science.1134351.
[17] ARISAN-ATAC I, HODITS R, KRISTUFEK D, et al. Purification, and characterization of a β-mannanase of Trichoderma reesei C-30[J]. Applied Microbiology Biotechnology, 1993, 39(1): 58-62. DOI:10.1007/BF00166849.
[18] SHENTAL-BECHOR D, LEVy y. Folding of glycoproteins: toward understanding the biophysics of the glycosylation code[J]. Current Opinion in Structural Biology, 2009, 19(5): 524-533. DOI:10.1016/ j.sbi.2009.07.002.
[19] GUO M J, HANG H F, ZHU T C, et al. Effect of glycosylation on biochemical characterization of recombinant phytase expressed in Pichia pastoris[J]. Enzyme Microbial Technology, 2008, 42(4): 340-345. DOI:10.1016/j.enzmictec.2007.10.013.
[20] XI H X, TIAN y P, ZHOU N D, et al. Characterization of an N-glycosylated Bacillus subtilis leucine aminopeptidase expressed in Pichia pastoris[J]. Journal of Basic Microbiology, 2015, 55(2): 236-246. DOI:10.1002/jobm.201400368.
[21] WANG C Q, EUFEMI M, TURANO C, et al. Influence of the carbohydrate moiety on the stability of glycoproteins[J]. Biochemistry, 1996, 35(23): 7299-7307. DOI:10.1021/bi9517704.
[22] 齐飞飞. N-糖基化修饰对瑞氏木霉纤维二糖水解酶Ⅰ分泌、酶活和蛋白稳定性的影响及影响机制的研究[D]. 济南: 山东大学, 2014.
[23] HAN M H, WANG W X, WANG X F, et al. Enhanced expression of recombinant elastase in Pichia pastoris through the substitution of Thr for Ser in Asn-Xaa-Ser sequons[J]. Applied Biochemistry Biotechnology, 2015, 175(1): 428-435. DOI:10.1007/s12010-014-1284-5.
[24] VAQUERO M E, BARRIUSO J, MEDRANO F J, et al. Heterologous expression of a fungal sterol esterase/lipase in different hosts: effect on solubility, glycosylation and production[J]. Journal of Bioscience Bioengineering, 2015, 120(6): 637-643. DOI:10.1016/ j.jbiosc.2015.04.005.
[25] 谢春芳, 黎玉凤, 刘大岭, 等. 利用N-糖基化修饰对β-甘露聚糖酶Man47的稳定性改造[J]. 中国生物工程杂志, 2013, 33(12): 79-85.
[26] MILLER G L. Use of dinitrosalicylic acid reagent for determination of reducing sugar[J]. Analytical Chemistry, 1959, 31(3): 426-428. DOI:10.1021/ac60147a030.
[27] 江小华. 里氏木霉木聚糖酶的分离纯化、酶学特性及降解机理的研究[D]. 南京: 南京林业大学, 2006.
[28] 樊艳爽. β-葡萄糖醛酸苷酶的N-糖基化设计及热稳定性研究[D].北京: 北京理工大学, 2015.
[29] TROMETTA E S. The contribution of N-glycans and their processing in the endoplasmic reticulum to glycoprotein biosynthesis[J]. Glycobiology, 2003, 13(9): 77R-91R. DOI:10.1093/glycob/cwg075.
[30] MACAULEY-PATRICK S, FAZENDA M L, MCNEIL B, et al. Heterologous protein production using the Pichia Pastoris expression system[J]. Yeast, 2005, 22(4): 249-270. DOI:10.1002/yea.1208.
Effect of N-Glycosylation on the Heterologous Expression of β-Mannanase in Pichia pastoris
MA Qing1, CAI Rui1, JIANG Fengchao1, MA Lijuan1,2,*, DU Liping1,2, XIAO Dongguang1,2
(1. Key Laboratory of Industrial Microbiology, Ministry of Education, College of Biotechnology, Tianjin University of Science and Technology, Tianjin 300457, China; 2. Tianjin Food Safety & Low Carbon Manufacturing Collaborative Innovation Center, Tianjin 300457, China)
In order to investigate the influence of N-glycosylation on the expression of β-mannanase from Trichoderma reesei (Man1) in Pichia pastoris GS115, the asparagine (Asn) residues at three N-linked glycosylation sites (N131, N158 and N329) of Man1 were substituted by neutral glutamine (Gln) through site-directed mutagenesis. The results showed that mutations of the N-glycosylated sites had no significant effects on the expression of Man1 at the transcriptional level.
Compared with Man1, the apparent molecular mass of the mutant Man1 decreased slightly, and the activities of the mutants (N131, N158 and N329) decreased by 85.43%, 79.48% and 16.3%, respectively. However, the thermal stability of mutant N131, N158 and N329 increased by 7.87%, 13.5% and 15.37% compared to that of Man1. Therefore, N-glycosylation especially at N131 and N158 was essential for the high-level expression of Man1 in P. pastoris GS115, but led to a slight reduction in the thermal stability of Man1.
β-mannanase; N-glycosylation; site-directed mutagenesis; enzyme activity; thermal stability
10.7506/spkx1002-6630-201716013
Q55
A
1002-6630(2017)16-0086-06
马清, 蔡瑞, 姜风超, 等. N-糖基化对β-甘露聚糖酶在毕赤酵母中异源表达的影响[J]. 食品科学, 2017, 38(16): 86-91. DOI:10.7506/spkx1002-6630-201716013. http://www.spkx.net.cn
MA Qing, CAI Rui, JIANG Fengchao, et al. Effect of N-glycosylation on the heterologous expression of β-mannanase in Pichia pastoris[J]. Food Science, 2017, 38(16): 86-91. (in Chinese with English abstract)
10.7506/spkx1002-6630-201716013. http://www.spkx.net.cn
2016-10-20
天津科技大学青年教师创新基金项目(2014CXLG10);工业发酵微生物教育部重点实验室暨天津市工业微生物重点实验室(天津科技大学)开放基金项目(2014IM101);天津市自然科学基金重点项目(16JCZDJC31800)
马清(1993—),女,硕士研究生,研究方向为酶与分子生物学。E-mail:399491057@qq.com
*通信作者:马立娟(1981—),女,助理研究员,博士,研究方向为生物质资源综合利用、食品微生物发酵。
E-mail:malj@tust.edu.cn
猜你喜欢
中国特种设备安全(2021年7期)2022-01-19
现代畜牧科技(2021年6期)2021-07-16
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与机械(2019年1期)2019-03-30
医学研究杂志(2015年12期)2015-06-10
中国酿造(2014年9期)2014-03-11
湖南农业科学(2014年5期)2014-02-27
中国粮油学报(2014年7期)2014-02-06
现代检验医学杂志(2014年1期)2014-02-06
