
九宫山红椿种群结构和空间分布格局分析
2017-08-22 06:41张华香汪洋冷艳芝杨旭尹茜
湖北农业科学 2017年14期
张华香+汪洋+冷艳芝+杨旭+尹茜


摘要:对九宫山保护区惟一红椿(Toona ciliata Roem.)群落设立2个样地,研究了T1和T2样地种群空间分布格局。通过龄级划分研究种群年龄结构和动态量化分析,以探讨不同样地红椿种群空间结构与分布格局的成因。结果显示,2个样地的种群分布格局在16、25、50、100 m2时,均为泊松分布;取样面积越大,Lloyd平均拥挤度(m*)和丛生指数(I)越大;T2样地的Lloyd平均拥挤度(m*)和丛生指数(I)值均高于T1样地。T1和T2样地Ⅰ龄级数量分别占总量的41.86%和48.18%,Ⅳ、Ⅴ龄级环境筛与竞争作用加强。2个样地种群动态指数变化差异较大,T1样地为10个龄级,VⅣ、VⅤ、VⅥ为波动;T2样地12个龄级,VⅣ、VⅤ、 VⅨ、VⅩ为波动。2个样地种群Vpi1=40.83 %,V′pi1=2.041,Vpi2=49.72%,V′pi2=4.144,均属于增长性种群,天然更新较好,但种群老龄化趋势明显。因此,加强森林抚育,人为制造林窗,将更有利于红椿种群的更新和稳定发展。
关键词:红椿(Toona ciliata Roem.);种群结构;空间分布格局;九宫山
中图分类号:S792.33+5(634TS) 文献标识码:A 文章编号:0439-8114(2017)14-2685-05
DOI:10.14088/j.cnki.issn0439-8114.2017.14.022
Abstract: 2 sampling plots were set up to research the spatial distribution pattern of Toona ciliata Roem. population. The structure of age class and quantitive dynamic indexes were analyzed to identify the causes of structure and distribution patterns of T. ciliata populations in the Jiugong Mountain Region. The result revealed that the distribution pattern in 16 m2, 25 m2, 50 m2 and 100 m2 grid were all Poisson distribution. Both the David & Moore's cluster index(I) and mean crowding index(m*) increased with the sampling size; and the 2 indexes of T2 sampling plot were higher than those of T1. The quantities of the Ⅰ age class accounted for 41.86% and 48.18% of the 2 plots, respectively; while environmental sieves and competitive effect intensified in Ⅳ to Ⅴ age classes. Population dynamics index of the 2 plots changed a lot. T1 plot has only 10 age classes, fluctuated negatively in VⅣ, VⅤ and VⅥ, with Vpi1=40.83% and V′pi1=2.041; T2 plot has 12 age classes, waved negatively in VⅣ, VⅤ, VⅨ, and VⅩ, with Vpi2=49.72% and V′pi2=4.144. Both the plots were growing populations with good natural population regeneration; but the aging tendency was evident. Therefore, the intensification of woods tending, and canopy gaps interfered by human management are conducive to the regeneration and balanced development of the population.
Key words: Toona ciliata Roem.; population structure; spatial distribution pattern; Jiugong Mountain
種群是构成群落的基本单位,种群分布格局能够反映种群空间变化规律,体现种群动态及其群落的演替趋势[1],研究种群空间格局对于分析和发现濒危植物的致危因素具有重要价值。由于种群分布格局是种群生物学特性、种内种间关系及环境条件综合作用的结果,是种群的基本数量特征之一[2],因此研究种群分布格局的目的不仅是对种群水平结构进行定量描述,更重要的是用以揭示格局的成因,阐述种群及群落的动态变化,为种群保护措施的制定提供一定的科学依据[3,4]。然而种群格局研究受取样面积的影响,在不同尺度上考察所得到的结论不同[5]。设置不同取样方式,是准确获取濒危植物种群空间分布格局的关键;研究种群结构不仅可以了解种群现在的状态,而且有助于分析种群未来的动态[6]。种群大小、结构不仅直接影响着群落结构,还能客观反映种群乃至整个群落的发展、演变趋势,且对种群动态变化有预测作用,是种群生态学研究的热点之一[7]。红椿(Toona ciliata Roem.)是楝科(Meliaceae)香椿属(Toona Roem.)落叶或半常绿大乔木,又名红楝子,为国家II级重点保护野生植物。其干形通直,树姿挺秀,自然整枝明显,属强阳性树种,是中国热带、亚热带地区珍贵速生用材树种,其木材为上等家具用材,素有“中国桃花心木”之称[8]。红椿天然分布范围从印度经缅甸、老挝、巴基斯坦、泰国、马来西亚、印度尼西亚至澳大利亚,在中国分布不广,仅呈天然零星分布,但发展潜力很大;不过由于人类的过度开发以及天然更新较慢,其数量不断减少[9]。鉴于红椿优良的用材特性和濒危现状,众多学者对其在种质资源、遗传育种、植物生理、栽培造林以及药用价值开发和化学成分分析、资源保护等方面进行了广泛研究,其中在种群生态学方面,国内已有冯立新等[10]研究了桂西北红椿种群年龄结构与空间分布格局及其濒危因素。但由于不同地理环境下红椿分布格局表现差异较大[11],需要进一步研究不同地理种源红椿的种群结构和空间分布格局,从而为红椿群落的保护和开发利用提供生态学基础依据。
1 材料与方法
1.1 研究区概况
湖北省通山县九宫山国家级自然保护区地处北纬29°19′27″-29°27′08″,东经114°23′35″-114°39′48″,海拔高度在117~1 656.7 m[12,13],属中亚热带季风型气候,并且山地气候特征明显。当地全年雨量充沛,年均降水量1 400~2 000 mm,雨热同季,多暴雨[14];年均日照时间为1 600 h,无霜期长,年均温度14.4 ℃,7月平均气温为26.5 ℃,1月平均气温为1.9 ℃[12,13]。研究样地位于保护区石龙沟,海拔519~545 m,土壤为山地黄棕壤[15]。植被类型为常绿阔叶林,主要乔木植物为红椿、青冈栎[Cyclobalanopsis glauca (Thunb.)Oerst.]、柃木(Eurya japonica Thunb.)、苦槠[Castanopsis sclerophylla(Lindl.) Schott.],色木槭(Acer mono Maxim.)、乌药[Lindera aggregata(Sims)Kosterm.]等,红椿为群落优势种和建群种。
1.2 野外调查方法
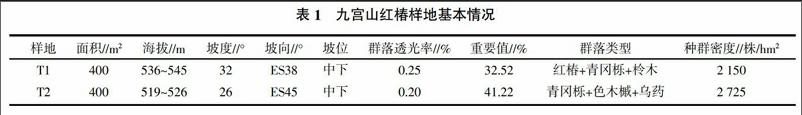
九宫山天然红椿群落为该地仅存的红椿群落,群落沿西南向东北走向的溪流分布,坡向东南,群落在溪边,长约60 m,宽约26~32 m,呈不规则长条状,总面积约1 800 m2。鉴于样地的不规则形状,同时为便于研究,取样覆盖了全部红椿存活植株,在群落中设置2个20 m×20 m的样地,样地基本情况见表1。运用相邻格子法在每块样地设置5 m×5 m的样方,每个样地各包含16个样方,共计32个。对样方内的红椿进行每木调查,对胸径(DBH)≤2.5 cm的红椿植株测定地径、高度;对DBH≥2.5 cm的记录树高、胸径、冠幅、枝下高、分枝角等指标。记录环境因子,对林下乔灌草藤本等植物也调查统计。
1.3 种群空间分布格局判定方法
为准确研究红椿种群分布格局与聚集强度,在样地内分别设定4 m×4 m、5 m×5 m、5 m×10 m、10 m×10 m的样方。同时采用7个分布格局数学模型进行分析,以避免各模型的片面性。7个分布格局数学模型分别是扩散系数(Cx)、负二项参数(K)、Cassie指标(Ca)、Lloyd平均拥挤度(m*)、聚块性指数(PAI)、David & Moore的丛生指数(I)、Morisita扩散型指数(Iδ),以此进行分布格局与聚集强度的判定[4,16,17]。
1.4 龄级划分及动态特征分析方法
由于红椿树龄与胸径之间具有较高的相关性[18],所以试验在红椿平均胸径与连年生长的过程分析中,参考曲仲湘[19]的立木分级标准,采用“空间替代时间法”对林木依胸径大小进行分级,以龄级结构代替种群径级结构分析种群的动态[20-22]。具体操作是从第二径级开始,以胸径5 cm為步长依次增加一级,则各龄级分别是:Ⅰ(幼苗级,2.5 cm 采用Leak[23]和陈晓德[24]的数学模型,推导种群年龄结构的动态指数,对红椿种群动态进行定量描述,公式如下: 式中,Vn表示种群从n到n+1级的个体数量变化动态指数;Vpi表示整个种群结构的数量变化动态指数;Sn和Sn+1分别表示第n到第n+1龄级种群的个体数,当考虑外部干扰时, 式中,K为种群龄级数量,Vpi与Vn取正、负、零值的意义分别反映种群或相邻龄级个体数量的增长、衰退、稳定的动态关系。P为种群对外界干扰所承担的风险概率,当P值为最大时,对种群动态Vpi构成最大的影响[20]。 2 结果与分析 2.1 红椿种群空间分布格局 种群分布格局指个体在水平空间的分布方式或者在水平空间上种群个体之间的相互关系[25];种群在一定的空间内都会呈现出特有的分布格局,它能够反映出种内或种间关系、适应环境机制、空间异质性等群落生态特征[26]。因此,课题组在九宫山天然红椿群落中取2个样地,分别研究空间分布格局,以揭示九宫山红椿不同样地空间分布格局的特征。为避免取样面积对空间分布格局结论的影响,采用7个聚集度指标对红椿种群实施空间分布格局分析,同时变换取样面积,分别设置若干小样方(4 m×4 m, 5 m×5 m,5 m×10 m,10 m×10 m)。对于扩散系数采用t检验;而Morisita扩散型指数则采用F检验。2个样地在不同样方面积时,df分别对应24、15、7、3;由此,九宫山天然红椿群落空间分布格局的判定结果见表2。从表2分析可见,扩散系数(Cx)的t检验结果表明,T1样地在16 m2取样面积时,Cx=1.407,>1,但采用Cx实测值与理论值1的差异显著程度在t检验后,t=1.410,
2.2 红椿种群年龄结构
九宫山红椿种群的结构与鄂西地区的红椿种群结构相似[11],但苗期植株数量明显较多,这可以保证种群的延续。T1样地红椿各龄级存活数量统计情况见图1,T2样地红椿各龄级存活数量统计情况见图2。从图1可以看出,在样地T1里红椿种群有10个龄级,属于增长型种群,其中Ⅰ龄级幼苗占有比率最大,达41.86%;Ⅱ龄级幼树为16.28%,Ⅲ~X龄级各类树总和为41.86%,以Ⅳ齡级幼树的存活数量最少,说明Ⅳ、Ⅴ龄级的环境筛与竞争作用加强。从图2可见,样地T2的红椿种群有12个龄级,属于衰退型种群,其中Ⅰ龄级幼苗占比达48.18%,Ⅱ龄级幼树为17.27%,其他龄级总计占比为34.55%,说明幼树到成年树经历了环境筛的选择或人为干扰;对比图1发现,在Ⅵ龄级前,T2与T1样地的龄级数量分布趋势一致,Ⅶ龄级后,种群损失数量很大。如果没有幼苗库的更新,种群必将衰退。从图1与图2还可见,2个样地的龄级数量分布预测方程y对应x的4次方程,为对应龄级植株数量的函数关系。
2.3 红椿种群年龄结构量化分析
种群相邻龄级间个体数量的变化动态分析结果见表3。从表3可见,2个不同样地的种群动态指数变化差异较大。T1样地的VⅠ、VⅡ、VⅢ数值较高,表明T1红椿种群呈增长动态;其后出现过3次波动(VⅣ、VⅤ、VⅥ),即种群在Ⅴ龄级、Ⅵ龄级和Ⅶ龄级转变为衰退动态;对应VⅧ=0,说明到Ⅷ龄级时,种群处于稳定动态;VⅨ=50.00%,表明种群再一次进入增长动态。Vpi1=40.83%,V′pi1=2.041,种群为增长型。T2样地的VⅠ、VⅡ、VⅢ也为增长动态,其幼苗丰富,幼苗到幼树阶段增长较好。虽然在不同龄级间均出现4次衰退动态(VⅣ、VⅤ、VⅨ、VⅩ),但总体变化指数Vpi2=49.72%,V′pi2=4.144,说明种群为增长型种群。种群动态增长指数(Vpi)是T2>T1,说明不同样地种群增长能力有大小之分;干扰条件下的动态指数(V′pi)T2>T1,表明T2样地种群对外界风险的调控能力优于T1。由于丰富的幼苗和萌蘖枝维系了种群整体稳定,所以总体上看,九宫山红椿种群幼苗丰富,幼苗到幼树阶段增长较好,天然更新较强。
3 小结与讨论
自然界中的濒危植物种群其年龄结构多呈衰退型,但在适合的生境条件下呈增长型,并且在自然条件下能够实现自我更新[27]。红椿的年龄结构及其分布格局主要由其生物学特性和环境条件共同决定,由于红椿为强阳性树种,生长喜光,除了幼苗能耐一定的阴蔽环境外,成年红椿植株对光的需求很高,因此在郁闭度较高的天然林中更新较差。九宫山红椿虽然所处的群落郁闭度较大,但群落所在的坡向与坡位使得群落光照条件得到满足。因此,环境筛的选择向高龄级推移,种群在低龄级时属于增长型。该特征与鄂西地区红椿种群动态存在差异。但鄂西地区所发现的天然红椿种群结构与桂西北地区红椿种群结构差异明显,主要表现在桂西北红椿种群整体上呈现幼龄期和老龄期的个体数量较少,中龄期的个体数量较多[10];鄂西地区红椿种群结构特征为幼龄期个体数量较多,幼龄期与中龄期期间个体数量损失严重,老龄期的个体数量较少,但种群基本呈增长或稳定型,种群衰老较突出,形成典型小种群。这种特定结构与环境因子相结合,制约了种群的空间分布格局。
种群空间分布格局分析表明,九宫山红椿种群在不同尺度(16、25、50、100 m2)上均属于泊松分布,最为典型表现泊松分布的取样尺度为25 m2。聚集强度最强的取样尺度为50 m2和16 m2,聚集强度变化无明显趋势,但与扩散系数变化趋势较为一致,这说明了取样尺度在研究分布格局上的重要性;当取样面积为25 m2时,可能为红椿种群自然更新的最佳尺度。Lloyd平均拥挤度随取样面积的增大而增大,表明种群个体相对拥挤程度增大,导致种内种间竞争加强,红椿聚集性分布转变为相对较为均匀的泊松分布,形成了红椿天然种群自身分布特点。T2样地的Lloyd平均拥挤度和丛生指数绝对值均高于T1样地。T1和T2样地Ⅰ龄级数量分别占总量的41.86%和48.18%,Ⅳ、Ⅴ龄级环境筛与竞争作用加强。2样地种群动态指数变化差异较大,T1样地为10个龄级,VⅣ、VⅤ、VⅥ为波动;T2样地12个龄级,VⅣ、VⅤ、VⅨ、VⅩ为波动。2样地种群Vpi1=40.83%,V′pi1=2.041,Vpi2=49.72%,V′pi2=4.144,均属于增长性种群,天然更新较好,但种群老龄化趋势明显。
自然保护区的建立对于红椿等濒危植物起到了良好的保护作用,然而濒危植物囿于自身生理特点对生境条件适应的局限性仍然很大。因此,在森林抚育中,人为制造林窗可以尽量接近聚集性最强的面积尺度,这样就能够将有利的自然因素通过耦合效应与人为活动效应结合起来[16],从而更有利于红椿种群的更新和稳定发展。
参考文献:
[1] 张 超,黄清麟,普布顿珠,等.西藏灌木林种群分布格局[J].山地学报,2011,29(1):123-128.
[2] 周纪纶,郑师章,杨 持.植物种群生态学[M].北京:高等教育出版社,1992.32-42.
[3] GREIG-SMITH P J. The use of random and contiguous quadrats in the study of the structure of plant communities[J]. Annals of Botany,New Series,1952,16:293-316.
[4] 康华靖,陈子林,刘 鹏,等.大盘山自然保护区香果树种群结构与分布格局[J]. 生态学报,2007,27(1):389-396.
[5] 李 博.现代生态学讲座[M].北京:科学出版社,1995.1-9.
[6] 龚 滨,夏洋洁,张光富,等.中国特有珍稀濒危树种银缕梅种群结构和空间格局[J].生态与农村环境学报,2012,28(6):638-646.
[7] WU X P,ZHENG Y,MA K P. Population distribution and dynamics of Quercus liaotungensis,Fraxinus rhynchophylla and Acer mono in Dongling Mountain,Beijing[J].Journal of Integrative Plant Biology,2002,44(2):212-223.
[8] 鄭万钧.中国树木志 第一卷[M].北京:中国林业出版社,1983.
[9] 汪 洋,冉勇军,冷艳芝,等.红椿和毛红椿遗传与生理生态相关研究[J].安徽农业科学,2014,42(20):6910-6912.
[10] 冯立新,陈 荣,朱昌叁,等.桂西北红椿种群年龄结构与空间分布格局研究[J].西北林学院学报,2015,30(1):46-50.
[11] 汪 洋,冷艳芝,宋丛文,等.恩施天然红椿种群结构及空间分布格局[J].浙江农林大学学报,2016,33(1):17-25.
[12] 杨新国,王 建.湖北九宫山自然保护区植被景观类型划分及景观多样性差异原因探析[J].广东农业科学,2011,38(23):145-148.
[13] 胡 静.基于生物多样性保护条件下的旅游资源开发研究[J].经济地理,2005,25(3):426-429.
[14] 陈进明,刘 星,杨春锋,等.九宫山自然保护区珍稀濒危植物调查研究[J].武汉植物学研究,2002,20(1):62-65.
[15] 张光远.湖北省九宫山土壤的垂直分布及其性状[J].华中农学院学报,1984,3(4):58-67.
[16] 张文辉,许晓波,周建云,等.濒危植物秦岭冷杉种群空间分布格局及动态[J].西北植物学报,2005,25(9):1840-1847.
[17] MICHAEL J C. Plant Ecology[M]. Oxford London Edinburgh: Blackwell Scientific Publications,1983.473-524.
[18] 龙汉利,冯 毅,向 青,等.四川盆周山地红椿生长特性研究[J].四川林业科技,2011,32(3):37-41.
[19] 曲仲湘.植物生态学[M].北京:高等教育出版社,1982.102-118.
[20] 吕海英,王孝安,李 进,等.珍稀植物银砂槐中国分布区的种群结构与动态分析[J].西北植物学报,2014,34(1):177-183.
[21] 杨 宁,邹冬生,杨满元,等.贵州雷公山秃杉的种群结构和空间分布格局[J].西北植物学报,2011,31(10):2100-2105.
[22] 韩 路,王海珍,彭 杰.塔里木河上游天然胡杨林种群空间分布格局与动态研究[J].西北植物学报,2007,27(8):1668-1673.
[23] LEAK W B. Age distribution in virgin red spruce and Northern hard woods[J]. Ecology,1975,56:1451-1454.
[24] 陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.
[25] 李先琨,黄玉清,苏宗明.元宝山南方红豆杉种群分布格局及动态[J].应用生态学报,2000,11(2):169-172.
[26] 张育新,马克明,祁 建,等.北京东灵山海拔梯度上辽东栎种群结构和空间分布[J].生态学报,2009,29(6):2789-2796.
[27] 张文辉,祖元刚,刘国彬.十种濒危植物的种群生态学特征及致危因素分析[J].生态学报,2002,22(9):1512-1520.
