
山核桃种子生长发育过程油脂含量及脂肪酸的动态变化
2017-08-07 10:24:00徐亚楠叶晓明曾燕如赵国淼唐研耀杨珺雯
浙江林业科技 2017年2期
徐亚楠,周 伟,叶晓明,曾燕如,刘 力,赵国淼,贾 宁,唐研耀,杨珺雯
(浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
山核桃种子生长发育过程油脂含量及脂肪酸的动态变化
徐亚楠,周 伟,叶晓明,曾燕如,刘 力,赵国淼,贾 宁,唐研耀,杨珺雯
(浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
2014年7月23日、8月1日、16日和31日,采用索氏抽提法和气相色谱法(HPLC)测定了浙江省临安市山核桃Carya cathayensis产区同林地中30个单株种子种仁的油脂含量及脂肪酸的组分,研究山核桃种子生长发育过程中油脂含量及脂肪酸的动态变化。结果表明,随着种子的生长发育和不断成熟,油脂含量增加很快,4个时间点的平均值分别为9.87%,38.30%,72.06%,71.65%,各时间点及各单株间油脂含量差异显著;山核桃油脂中主要脂肪酸棕榈酸、硬脂酸、油酸、亚油酸和亚麻酸平均相对含量的变幅分别为15.48% ~ 4.57%,2.38% ~ 1.50%,20.45% ~ 66.01%,45.24% ~ 21.48%,8.49% ~ 1.54%,仅油酸相对含量逐步增加,成熟种子中饱和脂肪酸主要是棕榈酸,不饱和脂肪酸主要是油酸;各时间点主要不饱和脂肪酸含量高于饱和脂肪酸含量。
山核桃;种子;油脂;饱和脂肪酸;不饱和脂肪酸;动态变化
植物油脂是一种高分子天然有机化合物,它的主要成分包括甘油酯和脂肪酸。种子含油率、脂肪酸的组成和含量直接决定植物油脂的营养价值和用途[1-2]。国内外对多种油料作物的含油率差异和脂肪酸变化规律进行了研究,其中对油茶Camellia oleifera的研究多集中在不同成熟期、种源和品种间的含油率差异和脂肪酸变化分析[3-5];在花生Arachis hypogaea中,Onemli[6]对3个花生品种,Wang等[7]对花生不同品种间的含油率和脂肪酸的差异进行了研究。种子含油率和脂肪酸组分的变化受遗传因子与环境互作的影响。Uzun等对不同地理位置栽培的芝麻Sesamum indicum进行了研究,结果表明遗传因子和环境的交互作用对含油率和脂肪酸含量的影响极显著[8];西班牙杏仁Prunus amygdalus也不例外[9]。
山核桃Carya cathayensis系胡桃科Juglandaceae山核桃属Carya植物,是木本油料植物中含油率极高的经济树种,目前处于半野生状态,主要分布于浙皖交界的天目山区,30°18'30" ~ 30°24'55" N,119°23'47" ~ 119°28'27" E[10-11]。山核桃具有极高的营养价值和独特的口感,其种仁含有丰富的人体必需氨基酸和矿质营养元素,具有广阔的开发应用前景[12]。相对于油茶等其他木本油料植物,山核桃优株油脂含量变异系数小(1.67%),其不饱和脂肪酸含量高达95.38%[13]。目前有关山核桃种子的研究主要侧重于山核桃结实规律[14-15]、油脂的提取及成分分析[16-17]、营养价值[18-19]、油脂形成相关基因(种类)[20-22]等。就山核桃油脂含量和脂肪酸组成的分析,多以该物种混合种子样品[14,23]或基于优选优株的种子[13]为材料进行测定,或对不同地点山核桃混合种子样品进行分析[15],而同一天然群体中不同单株间山核桃种子发育过程中油脂含量的动态变化及其脂肪酸组成的变异分析则鲜有报道。
以浙江省临安市太湖源镇指南村山核桃天然林的30棵山核桃单株的种子为对象,对其生长发育过程中4个时间点的种仁油脂含量和脂肪酸组成进行了分析,以便为后续山核桃成油机理研究打下基础。
1 材料与方法
1.1 材料
山核桃为风媒花,雌雄异花同株,雌雄花同时开放;雌花授粉后柱头即由紫红色变成紫黑色[24]。山核桃果实生长发育过程呈明显的阶段性,第1阶段为果实的增大期,从5月初至8月初,也是种子外部形态生长发育期,其中6月10日至7月10日为快速生长期;第2阶段为种仁填充期,从8月初至9月采收[14]。成熟种子于白露采摘,可食种仁为其胚,主要是膨大的子叶,无胚乳,无壁胚乳在种子生长发育的前期被发育的胚所吸收。
张通[25]于2012,2013 年采用索氏抽提法对浙江省临安市太湖源镇指南村同一单株山核桃(树龄30 a以上)测定了油脂含量,分别为7.35%(2012)/6.13%(2013),35.78%(2012)/29.95%(2013),60.45%(2012)/61.55%(2013),63.43%(2012)/71.33%(2013),其中前3个时间点油脂含量增量比较大,后一个时间点的值较前一个时间点的值几乎成倍增加,是油脂增加较快的时期,绘制了动态变化曲线。本试验在同一地点的山核桃林中随机选择30株正值结果旺盛期的健康大树,采样单株间距保持在50 m以上,分别编号Y1,Y2……Y30。2014年授粉后79 d(T1,7月23日),88 d(T2,8月1日),103 d(T3,8月16日),118 d(T4,8月31日)采集种子。第一次采样由于处于果实发育的初期,胚小,处于合子胚迅速发育的起始阶段[26],用于油脂重复测定的干基需求量大,因此每棵树采集1 000个左右的果实;后3次随着果实的进一步发育,种子逐渐充实,每棵树的采样量相继减少,最后一次每棵树采集50粒种子。采集的种子当天贮于冷库中,并及时取胚进行干燥处理。
1.2 方法
1.2.1 样品处理 每次从每棵树采得的种子样品剥取种胚后混合,并进行样品处理及分析。将12.5 cm×12.5 cm的脱脂双圈定性滤纸(英国Whatman®)折成滤纸包,放入65℃电热鼓风干燥箱(Xtemp,XT5118OV系列)中干燥至恒重M(最后2次称量的重量差异不超过0.05%,下同),放入磨口干燥器中备用。将剥取的山核桃胚置于培养皿中,放入65℃干燥箱干燥至恒重并用液氮充分混合研磨(A11分析研磨机,德国IKA)。取2 g样品放入滤纸包中,65℃干燥箱干燥至恒重M1,放入磨口干燥器中备用。
1.2.2 脂肪含量的测定 采用索氏抽提法(YG-2型索氏抽提器),用化学纯石油醚(沸程30 ~ 60℃,国药集团化学试剂有限公司)对样品进行抽提。抽提前每个时间点每株树3次重复的样品在同一抽提管中用石油醚浸泡过夜。抽提时温度控制在53 ~ 55℃,石油醚回流抽提至少10 h。抽提完毕后从抽提管中取出滤纸包,在水浴中蒸馏回收抽提瓶中的石油醚。当抽提瓶剩余大约30 mL石油醚时,取下抽提瓶,在通风柜中使残余的石油醚挥发,余下的同一时间点同一单株的混合油样用于后续脂肪酸的测定。抽提后的滤纸包放入65℃干燥箱内干燥至恒重M2。
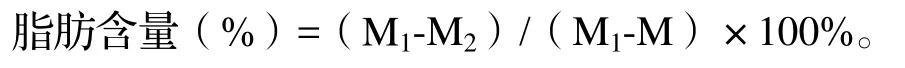
1.2.3 脂肪酸的测定 参照GB/T 17376-2008方法对山核桃油进行甲酯化,取60 mg山核桃油样至5 mL的带塞试管中,用移液管移取4 mL异辛烷(色谱纯,上海国药试剂公司)溶解试样,用微量移液管加入200 µL氢氧化钾甲醇溶液(浓度为2 mol·L-1),盖上玻璃塞剧烈摇晃30 s后静置至澄清。然后向溶液中加入约1 g硫酸氢钠,剧烈摇晃,中和氢氧化钾,静置至溶液完全分层。取2 µL上清液直接注入GC-2014C型气相色谱测定仪(日本岛津公司)的色谱柱中进行脂肪酸测定分析。气相色谱条件:毛细色谱柱规格为30 m×0.32 mm×0.25 µm;起始温度为130℃,保持4 min,在4 min内升至210℃后保持10 min;载气为氮气;柱流速为1.52 mL·min-1;分流比为15:1;进样口温度为200℃;检测器温度为230℃。试验所用的标样共11种(美国AccuStandard公司),即肉豆蔻酸甲酯、棕榈酸甲酯、硬脂酸甲酯、油酸甲酯、亚油酸甲酯、亚麻酸甲酯、花生酸甲酯、甲基二十碳烯酸甲酯、山嵛酸甲酯、芥酸甲酯、木蜡酸甲酯。按照标样的保留时间对样品中各脂肪酸的种类进行定性。采用峰面积归一化法计算各脂肪酸的相对含量[27]。
1.2.4 数据处理 试验数据处理及作图采用Excel 2007,SPSS 18.0,Sigma plot 12.5软件。油脂含量测定结果为百分数,经反正弦转换后再进行方差分析;方差分析差异显著的则采用最小显著差数法(LSD)进行多重比较[28]。
2 结果与分析
2.1 不同单株间种子油脂含量的分析
供试各单株山核桃种仁油脂含量在种子生长发育与成熟过程4个时间点的平均值分别为9.87%,38.30%,72.06%,71.65%。随着山核桃种子不断地生长发育,油脂含量总体呈现逐步增加的趋势。T1至T3油脂积累迅速,从T1的平均9.87%增加到T3平均72.06%,高的单株达75.79%,且T3的油脂含量是T2油脂含量的近2倍;T3以后油脂含量增长缓慢,基本保持稳定(图1)。但研究发现,T1至T2各单株种子的油脂含量虽呈上升趋势,但彼此间还是有差异的,此后单株间的差异逐渐减小。
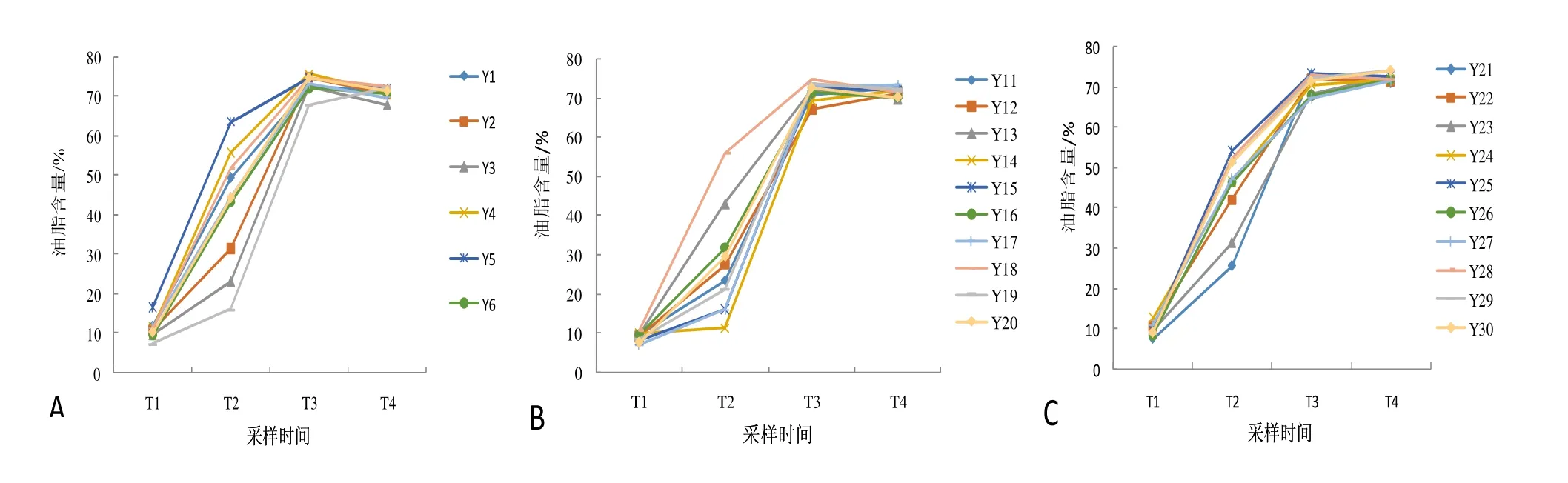
图1 不同采样时间30个单株山核桃种子油脂含量的动态变化Figure 1 Dynamic changes of oil content in kernels from 30 hickory trees at different sampling dates
种子油脂含量是由多因素控制的复杂数量性状。方差分析表明(表1),山核桃种子油脂含量在不同发育时间、不同单株间,以及种子发育时间与各单株交互作用之间差异极显著。多重比较发现(表2),种子油脂含量在T3与T4间差异不显著;除此之外,不同种子发育时间两两之间存在极显著的差异。这个结果与种子生长发育期种子油脂含量动态变化的曲线相吻合。

表1山核桃种子油脂含量的双因素方差分析Table 1 Two-way analysis of variance on oil content in kernels

表2山核桃种子油脂含量在不同种子发育时间的多重比较Table 2 Multiple comparison on oil content in kernels from 30 hickory trees at different sampling dates

表3山核桃种子不同发育时间不同单株间油脂含量两两比较差异显著组合分析Table 3 Pairwise comparison on oil content in kernels from 30 hickory trees at different sampling dates
将各时间点不同单株间两两共435个组合进行多重比较,结果见表3。由表3可知,随着种子的生长发育,差异显著的组合所占比例逐渐增加,T2时比例最高,达97.01%,到T3时比例有所减少,但幅度还不是很大;种子近成熟采收时,差异显著的组合所占的比例减至38.85%,其中差异显著的组合中绝大多数组合差异极显著,说明同一地点山核桃种群内种子生长发育过程中,同一时间点不同单株的种子油脂含量存在差异。
2.2 种子脂肪酸的分析
2.2.1 种子中脂肪酸的种类 在供试样品中基于已有的标样共检出11种脂肪酸,但不同时间点不同样品中检出的种类并不完全相同,其中以种子内部胚发育初期(T1)最多,大多数样品都检出11种脂肪酸,到T3、T4时每个时间点仅有0 ~ 2个样品能够检出全部11种脂肪酸。山核桃油脂中主要的脂肪酸有5种,即棕榈酸(也即软脂酸)、硬脂酸、油酸、亚油酸、亚麻酸[16],其中棕榈酸、硬脂酸是饱和脂肪酸,其他3种为不饱和脂肪酸;标样中其余6种脂肪酸中既有饱和脂肪酸(肉豆蔻酸、花生酸、山嵛酸),也有不饱和脂肪酸(甲基二十碳烯酸、芥酸、木蜡酸)。在气相色谱图中,除了11种内标脂肪酸外,还存在其他未知的脂肪酸,但这部分脂肪酸峰面积非常小,可能存在山核桃特有的脂肪酸。
在种子生长发育期各时间点,5种主要脂肪酸相对含量总和占总脂肪酸的90%以上,在90% ~ 97%;其它6种脂肪酸(肉豆蔻酸、花生酸、甲基二十碳烯酸、山嵛酸、芥酸、木蜡酸)相对含量比较低,含量不到总脂肪酸10%。此外,影响植物油脂氧化稳定性的主要是棕榈酸、硬脂酸、油酸、亚油酸以及亚麻酸[29],且采用GC-MS法验证了这5种脂肪酸也是山核桃仁油的主要脂肪酸[30]。因此,对这5种脂肪酸进行动态变化的分析。
2.2.2 饱和脂肪酸相对含量的动态变化 由表4发现,山核桃种子生长发育过程中主要的饱和脂肪酸变异系数在13.21% ~ 34.99%,平均值的变幅为17.85% ~ 5.72%,呈下降趋势,近成熟时的值为初始值的三分之一左右,且T2变异最大。棕榈酸、硬脂酸的相对含量也逐渐下降,平均值的变幅分别为15.48% ~ 4.57%、2.38% ~ 1.50%,其中只有少数样品在T4未检测出硬脂酸;棕榈酸相对含量变异最大时间出现在T2,而硬脂酸相对含量变异最大时间出现在T1,且T1与T2的硬脂酸相对含量变异系数相差不大。

表4山核桃种子不同发育时间饱和脂肪酸含量分析Table 4 Saturated fatty acid content in kernels from 30 hickory trees at different sampling dates
2.2.3 不饱和脂肪酸相对含量的动态变化 从表5可知,山核桃不饱和脂肪酸主要有油酸、亚油酸和亚麻酸。在山核桃种子生长发育及成熟过程中,不饱和脂肪酸相对总量逐渐增加,平均值从74.197%增加到90.20%;T1时亚油酸相对含量高,其次为油酸;T4时主要是油酸,其次是亚油酸。油酸的相对含量逐渐增加,T3均值达到67.18%,T4略有下降,但相对含量仍然很高,较T1增加了约3倍。亚油酸平均相对含量从T1时的45.24%降低至T3时的16.34%,到T4时平均相对含量又上升至21.48%,其平均相对含量与油酸呈彼此消长的关系,推测这两种脂肪酸之间可能存在相互转化。亚麻酸的平均相对含量T1时最高也仅8.49%,到T2时下降约4倍,最后T4时只有1.54%。
此外,油酸在T1时的变异系数最大,而亚油酸和亚麻酸在T2时的变异系数最大,与油脂含量的差异变化相一致。不同时间点不饱和脂肪酸相对含量的变异程度比饱和脂肪酸要小得多,且随着种子的成熟,这种变异程度越来越小。油酸相对含量的变异系数从T1时38.21%逐渐减小至T4时的13.35%,下降幅度较大。

表5 山核桃种子不同发育时间不饱和脂肪酸含量分析Table 5 Analysis on unsaturated fatty acids in kernels from 30 hickory trees at different sampling dates
3 讨论
随着山核桃果实的生长发育,其油脂含量呈明显的“S”型增长,这一结果不仅与周秦[22]的相似,且与大多数油料作物的曲线一致[31-32]。7月下旬至9月初是山核桃种仁储藏物质积累和转化的关键时期[14]。研究发现,T1、T2间的种子既有形态生长,又有包括油脂在内的物质的形成、转化与积累;T2、T3间种子的油脂含量有所增加,且增长倍数近2倍(38.30% ~ 72.06%),同时也伴随着种子内部物质转化及油质的变化;T3到T4种子的油脂含量略有下降,但差异不大,主要是山核桃种子无休眠期,种子内部在种子采收前已在为种子萌发进行物质转化。
山核桃为高油经济树种,其种仁油脂含量是由多因素控制的复杂数量性状,不仅受种子成熟度、果实发育期气候条件和环境因素的影响,还受自身遗传因子的影响[1]。本试验材料采集于山核桃产区同一林地中,环境条件基本一致,但各时间点不同单株间油脂含量存在差异,尤其在T2时,但接近种子采收时,不同单株间的差异减小。由此推测,山核桃油脂含量的差异主要是由山核桃本身物质转化与积累速率的差异造成的,而环境条件对油脂含量差异的影响较小。
脂肪酸的组成是评定油脂品质的重要指标。从分析结果来看,山核桃2种主要饱和脂肪酸中棕榈酸所占比例大,尤其在种子发育初期,而油酸是成熟山核桃种子中主要的不饱和脂肪酸,这一结果与田荆祥等[15]的分析结果一致。在山核桃种子成熟过程中,主要饱和脂肪酸的相对含量在不断减少,种子成熟时主要饱和脂肪酸的含量极微,而主要不饱和脂肪酸的相对含量在逐渐增加,尤其是单不饱和脂肪酸油酸的相对含量最高,且饱和脂肪酸和不饱和脂肪酸呈显著负相关关系(r=-0.982,P<0.05)。由此推测山核桃不饱和脂肪酸主要是由饱和脂肪酸转化而来,并符合油料作物代谢途径的一般规律,即在质体中首先合成饱和脂肪酸,接着发生氧化脱氢反应生成单不饱和脂肪酸油酸,然后进入内质网脱饱和生成多不饱和脂肪酸,最后各脂肪酸与3磷酸甘油结合生成三酰甘油[33]。此外,各时间点主要不饱和脂肪酸含量要高于饱和脂肪酸含量,T1也不例外,此时合子胚刚发育,说明山核桃种子中饱和脂肪酸转化成不饱和脂肪酸很快,可能饱和脂肪酸形成与转化同时在进行,在成油机理研究中值得进一步研究。油酸在T4时相对含量有所下降,而亚油酸的相对含量有所增加,且两者之间呈显著负相关关系(r=-0.990,P<0.05),两者间的转化规律有待进一步研究。总不饱和脂肪酸的变异程度在整个种子生长发育过程中不到10%,表明山核桃油脂稳定性高、品质优良[16]。因此,山核桃油脂不仅具有很高的营养价值[19],且为油质较好的食用油。结合油脂含量在T3与T4间差异不显著的研究结果,说明T3 ~ T4期间主要是种子内脂肪酸的相互转化,使得检出11种脂肪酸的样品数极少。
综上所述,在山核桃种子生长发育的初期,油脂含量增加趋势比较快,在种子发育后期及成熟期,其变化趋势比较缓慢,单株间油脂含量保持相对平稳,主要是脂肪酸转换而导致的油质变化。
[1] Weselake R J,Taylor D C,Rahman M H,et al. Increasing the flow of carbon into seed oil [J]. Biotechnol Adv,2009,27(6):866-878.
[2] Aruna R,Nigam S N. Inheritance of fatty acid content and related quality traits in groundnut (Arachis hypogaea L.) [J]. Oilseed Res,2009(26):10-17.
[3] Zarringhalami S,Sahari M A,Barzegar M,et al. Changes in oil content, chemical properties, fatty acid composition and triacylglycerol species of tea seed oil during maturity period [J]. J Food Biochem,2011,35(4):1161-1169.
[4] 郭夏丽,肖萍,杜尚广,等. 不同品系油茶种子脂肪酸组成分析[J]. 南昌大学学报(理科版),2013,37(1):43-46.
[5] 姚小华,王开良,黄勇,等. 小果油茶不同居群种仁含油率及脂肪酸组分变异特征分析及评价[J]. 林业科学研究,2013,26(5):533-541.
[6] Onemli F. Impact of climate change on oil fatty acid composition of peanut (Arachis hypogaea L.) in three market classes[J]. Chil J Agr Res,2012,72(4):483-488.
[7] Wang M L,Raymer P,Chinnan M,et al. Screening of the USDA peanut germplasm for oil content and fatty acid composition [J]. Biomass Bioenerg,2012,39:336-343.
[8] Uzun B,Arslan C,Furat S.Variation in fatty acid compositions,oil content and oil yield in a germplasm collection of sesame (Sesamum indicum L.) [J]. J Am Oil Chem Soc,2008,85:1135-1142.
[9] Kodad O,Estopanán G,Juan T,et al. Oil content, fatty acid composition and tocopherol concentration in the Spanish almond genebank collection[J]. Sci Hor,2014(177):99-107.
[10] 吕惠进,刘伯根. 浙西皖南山核桃资源及其开发利用[J]. 生态经济,2005(5):97-101.
[11] 吕芳德,黄菁. 山核桃属植物研究进展[J]. 经济林研究,2005,23(2):72-75.
[12] 郭传友,黄坚钦 方炎明. 山核桃研究综述及展望[J]. 经济林研究,2004,22(1):61-63.
[13] 黄坚钦,郑炳松,黄有军,等. 木本油料植物山核桃新品种的选育[J]. 生物质化学工程,2006,40(z1):178 -181.
[14] 解红恩,黄有军,薛霞铭,等. 山核桃果实生长发育规律[J]. 浙江林学院学报,2008,25(4):527-531.
[15] 田荆祥,吴美春,仲山民. 山核桃成熟过程中油脂变化的研究[J]. 浙江林学院学报,1986,3(2):53-57.
[16] 陆浩,杨会芳,毕艳兰,等. 山核桃油的理化性质及脂肪酸组成分析[J]. 中国油脂,2010,35(5):73-76.
[17] 周明亮,王鸿飞,赵丹. 山核桃油的提取工艺及其特性研究[J]. 农业机械学报,2007,38(3):95-98.
[18] 王冀平,李亚南. 山核桃仁中主要营养成分的研究[J]. 食品科学,1998,19(4):44-46.
[19] Zhu C,Deng X,Shi F. Evaluation of the antioxidant activity of Chinese Hickory (Carya cathayensis) kernel ethanol extraction[J]. Afr J Biotechnol,2008,7(13).
[20] 黄银芝,周秦,黄友军,等. 山核桃脂肪代谢相关cDNA文库的构建[J]. 浙江农林大学学报,2011,28(1):80-85.
[21] 黄银芝,曾燕如,周秦,等. 山核桃种子脂肪代谢期EST序列的初步分析[J]. 浙江农林大学学报,2015,32(2):229-236.
[22] 周秦. 山核桃果实成熟期间基因表达的初步分析[D]. 临安:浙江农林大学,2010.
[23] 张鹏,钟海雁,姚小华,等. 四种山核桃种仁含油率及脂肪酸组成比较分析[J]. 江西农业大学学报,2012,34(3):499-504.
[24] 黎章矩. 山核桃栽培与加工[M]. 北京:中国农业科技出版社,2003:43.
[25] 张通. 山核桃油脂合成和基因表达变化分析[D]. 临安:浙江农林大学,2015.
[26] 胡恒康. 山核桃合子胚发育、体胚再生及其生物学特性研究[D].南昌:江西农业大学,2011.
[27] 俞春莲,王正加,夏国华,等. 10个不同品种的薄壳山核桃脂肪含量及脂肪酸组成分析[J]. 浙江农林大学学报,2013,30(5):714 -718.
[28] 明道绪. 生物统计附试验设计[M]. 北京:中国农业出版社,2008:87-92.
[29] Chu Y H,Kung Y L. A study on vegetable oil blends [J] Food Chem,1998,62(2):191-195.
[30] 赵志刚. 山核桃仁油中脂肪酸组成的分析及评价[J]. 安徽农业科学,2009,37(6):2473-2474.
[31] 齐季,熊彪, 琚煜熙,等. 山胡椒果实生长发育规律及油脂变化研究[J]. 中国农学通报,2015,31(4):29-33.
[32] 石从广,李因刚,朱光权,等. 白花树种子成熟期含油率和脂肪酸变化规律[J]. 浙江农林大学学报,2013,30(3):372-378.
[33] 赵翠格,刘頔,李凤兰,等. 植物种子油脂的生物合成及代谢基础研究进展[J]. 种子,2010,29(4):56-62.
Dynamic Changes of Oil Content and Fatty Acids in Seeds of Carya cathayensis at Different Growth Stage
XU Ya-nan,ZHOU Wei,YE Xiao-ming,,ZENG Yan-ru,LIU Li,ZHAO Guo-miao,JIA Ning,TANG Yan-yao,YANG Jun-wen
(The Nurturing Station for the State Key Laboratory of Subtropical Silviculture, Zhejiang A & F University, Lin’an 311300, China)
Determinations by Soxhlet extraction and GC (gas chromatography)were implemented on oil content and components of fatty acids from nut kernels of 30 Carya cathayensis trees in the same stand located in Lin’an, Zhejiang province, sampled on July 23, August 1, August 16 and 31 of 2014. Results showed that oil content increased significantly along with maturation of seeds, the average content was 9.87%, 38.30%, 72.06% and 71.65% of samples with different dates. There was significant difference of oil content either among sampling dates or among individual trees. The mean content of main fatty acids in kernel, such as palmitic acid, stearic acid, oleic acid, linoleic acid, and linolenic acid ranged from 15.48% to 4.57%, 2.38% to 1.50%, 20.45% to 66.01%, 45.24% to 21.48% and 8.49% to 1.54% respectively. Only the average content of oleic acid increased gradually. The main saturated fatty acid in kernel was palmitic acid, and the main unsaturated one was oleic acid. The content of unsaturated fatty acids was always higher than that of saturated ones.
Carya cathayensis; seed; oil; saturated fatty acid; unsaturated fatty acid; dynamic change
S664.1
A
1001-3776(2017)02-0010-07
10.3969/j.issn.1001-3776.2017.02.002
2016-11-09;
2017-01-14
浙江省自然科学基金重点项目(Z13C160012)
徐亚楠,硕士研究生,主要从事经济林培育与利用研究;E-mail:xuyanan102@163.com。通信作者:曾燕如,教授,硕士生导师,主要从事经济林培育与利用研究,E-mail: yrzeng@zafu.edu.cn。
猜你喜欢
数学大王·中高年级(2021年3期)2021-03-10 11:19:30
新农业(2020年18期)2021-01-07 02:17:28
天然产物研究与开发(2018年9期)2018-10-08 03:26:00
中国民族医药杂志(2016年4期)2016-05-09 07:41:11
合成化学(2015年10期)2016-01-17 08:56:06
分析测试学报(2015年8期)2016-01-13 06:19:29
精细石油化工(2015年3期)2015-12-14 09:07:40
作文与考试·小学高年级版(2015年5期)2015-05-30 10:48:04
西安交通大学学报(医学版)(2015年2期)2015-02-28 17:59:15
食品科学(2013年14期)2013-03-11 18:25:06
