
棉铃虫发育起点温度和有效积温的地理变异
2017-08-02 01:39:31陈元生罗致迪钟平华
环境昆虫学报 2017年3期
陈元生,罗致迪,钟平华
棉铃虫发育起点温度和有效积温的地理变异
陈元生*,罗致迪,钟平华
(江西环境工程职业学院,江西赣州 341000)
为探明棉铃虫Helicoverpaarmigera不同地理种群发育起点温度和有效积温及其地理变异,推测该虫在不同地区可能发生的代数、为预测其发育发生期的差异提供依据,从而更有效地进行该虫的综合治理。以采集自中国的4个地理种群(广东广州(23.08° N,113.14° E)、江西永修(29.04° N,115.82° E)、山东泰安(36.15° N,116.59° E)、辽宁喀佐(41.34° N, 120.27° E)的幼虫在室内饲养繁育1代后为试材,采用在获得其在20℃、22℃、25℃和28℃恒温、L16∶D8的光周期条件下饲养的发育历期的基础上,比较研究这4个地理种群的幼虫、非滞育蛹和滞育蛹的发育起点温度和有效积温及其与栖息地纬度的关系。结果表明:在实验室人工饲料饲养的条件下,棉铃虫幼虫、非滞育蛹和滞育蛹发育起点温度分别为10.18℃-12.19℃、10.60℃-13.50℃、15.56℃-17.96℃,有效积温分别为254.91-300.89日度、126.21-204.43日度、181.03-363.39日度,滞育蛹的发育起点温度比非滞育蛹高3.39℃-5.50℃,其有效积温比非滞育蛹高35.56-165.89日度。这些说明棉铃虫发育起点温度和有效积温均存在明显地理变异,随栖息地纬度的升高,幼虫、非滞育蛹和滞育蛹的发育起点温度呈现逐渐下降趋势,幼虫的发育起点温度与栖息地纬度呈显著负相关关系(P<0.05),其余呈不显著负相关(P>0.05),而有效积温则相反,呈现上升趋势,滞育蛹有效积温与栖息地纬度升高呈显著正相关(P<0.05),其余呈不显著正相关(P>0.05)。
棉铃虫;发育起点温度;有效积温;地理变异;纬度;地理种群
昆虫是典型的变温动物,温度是影响昆虫生长发育的主要外因,发育起点温度(development threshold temperature, DTT)和有效积温(sum of effective temperatures, SET)是昆虫的基本生物学特性,是昆虫地理种群在当地能否完成发育的重要标志,也是预测昆虫发育进程和种群动态的重要指标。对昆虫的发育起点温度和有效积温的研究,有助于掌握其发育速度(Honěk, 1996; Jarosiketal., 2002),也有助于预测其发生期和发生代数(Bartekova and Praslicka, 2006)。
昆虫的热量需求变异与种群的地理起源(地理种群)有关,而且随地理纬度的增加,发育起点温度(LDT)降低、有效积温(SET)增加,且LDT与SET一般呈负相关(Trudgill, 1995)。Honěk(1996)报道了335种昆虫的发育起点温度和有效积温的地理变异,在热带(0-23°N or S),LDT较高且几乎无地理差异,而在亚热带(24-39°N or S)和温带地区(≥40°N or S),随地理纬度的升高,335种昆虫的LDT平均值显著降低,SET则随地理纬度升高而增加,且其变化幅度比LDT大(Honěk, 1996)。而同种昆虫不同地理种群的发育起点温度和有效积温的地理变异,已有不少报道。
多数情况下,同种昆虫不同地理种群的LDT同样也是随栖息地纬度升高而降低的,SET则相反。例如,小菜蛾Plutellaxylostella的发育起点温度与栖息地纬度呈负相关关系,而有效积温则呈正相关关系(Umeya and Yamada, 1973);甘蔗红粉蚧Saccharicoccussacchari菲律宾种群(低纬度)比埃及种群(高纬度)具有更高的LDT和更低的SET(Rae and De’ath, 1991);亚洲玉米螟一化性(黑龙江双鸭山,46.38°N)种群SET最高,二化性(吉林公主岭,43.31°N)种群次之,三化性(河北衡水,37.44°N)种群最低,卵、幼虫和蛹的LDT均为三化性的最高,二化性居中,一化性最低(夏新等,2007)。另外,还有步甲Pterostihusnigrita(Ferenz, 1975)、欧洲玉米螟Ostrinianubilalis(Lee and Spence, 1987)等。显然,上述昆虫种群通过LDT和SET的变异来适用其当地的生态环境。此外,也有些种类的昆虫在整个分布区域内发育稳定,即LDT和SET无变异或者差异不显著,如金眼草蛉Chrysopaoculeata(Necholsetal., 1987; Tauberetal., 1987),七星瓢虫Coccinellaseptempunctata(Phoofolo and Obrycki, 1995), 竹桃蚜Aphisnerii(Groeters, 1992),豌豆蚜AcyrthosiphonpisumHarris(Lambetal., 1987),茶足柄瘤蚜茧蜂LysiphlebustestaceipesCresson(Royeretal., 2001),樱桃绕实蝇Rhagoletiscerasi(Baker and Miller, 1978),褐色雏蝗Chorthippusbrunneus(Telfer and Hassall, 1999)等。
棉铃虫HelicoverpaarmigeraHübner是一种危险性的世界性的害虫,属夜蛾科Noctuidae,主要为害玉米和棉花,随着综合防治的开展,需要我们对其基本生物学特性及环境因素对这些特性的影响有更深刻的了解。资料表明,棉铃虫在滞育诱导(吴孔明和郭予元,1997;Shimizu and Fujisaki, 2006)、发育历期(陈元生和涂小云,2016)等方面存在地理变异,但棉铃虫发育起点温度和有效积温的地理变异,尚未见报道。有关棉铃虫发育起点温度和有效积温的报道虽然有不少(吴坤君等,1980;李锦华和李兹新,1985;李超等,1987;吴孔明和郭予元,1994;牟吉元等,1995;吴孔明和郭予元,1996;Qureshietal., 1999; Jallow and Matsumura, 2001; Bartekova and Praslicka, 2006),但这些研究所采用的实验种群不同,且实验条件及测定方法不完全一致,故所得到的实验结果(DTT和SET)差异很大,很难有可比性,因为昆虫通过人工饲料饲养所获得LDT和SET,不同于寄主材料饲养的LDT和SET,人工饲料饲养的棉铃虫生长发育要比多数寄主植物饲养的快(吴孔明和郭予元,1994;Casimeroetal., 2000; Jallow and Matsumura, 2001; Jallowetal., 2001),即实验条件(营养、温度设置等)对LDT及SET影响极大,所以我们不能依据不同人在不同的实验条件下所得到的数据汇总来类推棉铃虫LDT和SET是否存在地理变异。鉴此,本试验在相同的实验条件下,系统比较了广东、江西、山东和辽宁4个地理种群棉铃虫幼虫、非滞育蛹和滞育蛹的LDT及SET,旨在探明其LDT和SET的真实地理变异及其特点,为推测该虫在不同地区可能发生的代数、预测其发育发生期的差异提供依据,从而更有效地进行该虫的综合治理。
1 材料与方法
1.1 供试虫源
供试棉铃虫分别于2010年6-8月采自广东省广州市(以下简写为“GZ”,23.08°N,113.14°E)、江西省永修县(以下简写为“YX”,29.04°N,115.82°E)、山东省泰安市(以下简写为“TA”,36.15°N,116.59°E)、辽宁省喀佐县(以下简写为“KZ”,41.34°N,120.27°E)。幼虫在室内用人工饲料(参照Wu and Gong(1997)的方法)、在25℃、L16:D8的光周期条件下饲养,3龄前在24孔板内群养,3龄后在21孔冰格板内单养,成虫用复合维生素糖水(梁革梅等,1999)饲喂,备用。
1.2 历期测定方法
实验在恒温下进行,设置20℃、22℃、25℃和28℃4个恒温;光照时间为:16 h/d。
将4个地理种群的初孵幼虫(室内第1代)分别移入相应温度和光周期的培养箱中,观察并记录各组幼虫的生长发育及化蛹情况,然后将滞育蛹与非滞育蛹分开,分别观察记录其羽化情况,统计求得幼虫历期、非滞育蛹历期和滞育蛹历期。以上实验均在新苗光照培养箱GZX-250BS-Ⅲ中进行,光照强度为500-700 lx,箱内的温度变化幅度为±0.5℃。
棉铃虫滞育蛹的判断依据蛹的眼点移动情况判断,化蛹后10-15 d眼点位置仍无变化的蛹判定为滞育个体(Cullen and Browning, 1978)。
1.3 发育起点温度和有效积温的计算
上述所得的各虫态发育历期(D)(陈元生和涂小云,2016)换算成发育速率(R=1/D)。依据有效积温法则,采用直线回归法(Honěk, 1996;Bartekova and Praslicka, 2006),按温度(T)与发育速率(R)的线性回归方程:R=a+bT [a,b均为线性回归参数],计算各虫态发育起点温度(DTT)与有效积温(SET):DTT=-a/b;SET=1/b。
1.4 地理变异分析方法
运用SPSS 13.0统计软件中线性回归分析法(Linear-regression)分析处理各虫态LDT和SET与栖息地纬度关系。
2 结果与分析
2.1 幼虫发育起点温度和有效积温
不同地理种群棉铃虫幼虫的发育起点温度和有效积温见表1。由表1可知,GZ、YX、TA和KZ种群幼虫的发育起点温度分别为12.14℃、10.89℃、10.98℃和10.24℃,随纬度升高,幼虫发育起点温度逐渐下降。采用线性回归分析得到,雌性幼虫发育起点温度与栖息地纬度的相关性方程为:y=-0.0984x+ 14.564(R2= 0.8496,P<0.05),雄幼虫的发育起点温度与栖息地纬度的相关性方程为:y=-0.0857x+ 13.689(R2=0.6245,P<0.05),这说明,雌、雄幼虫的发育起点温度均随栖息地纬度升高而下降,呈显著负相关关系,而且雌虫的相关性表现更显著、明显些。
从表1还可见,GZ、YX、TA和KZ种群幼虫的有效积温分别为231.00、290.19、263.64和264.48日度,雌、雄幼虫发育有效积温均随栖息地纬度升高而呈现出增加的趋势,呈不显著的正相关关系(P>0.05)。
2.2 非滞育蛹发育起点温度和有效积温
不同地理种群棉铃虫蛹的发育起点温度和有效积温见表2。由表2可见,GZ、YX、TA和KZ种群非滞育蛹的发育起点温度分别为12.11℃、12.46℃、13.11℃和11.24℃,随纬度升高,雌、雄非滞育蛹发育起点温度均呈下降趋势,但线性关系不明显(雌蛹相关性方程:y=-0.0281x+13.472,R2=0.0473,P>0.05;雄蛹相关性方程:y=-0.0251x+ 12.784,R2=0.0283,P>0.05)。雌性非滞育蛹DTT,中纬度YX、TA种群(13.36℃、13.36℃)明显分别高于低纬度GZ种群(12.11℃)和高纬度种群(11.24℃),差异显著;而雄性DTT只TA种群高于其它种群,其差异也达显著水平。
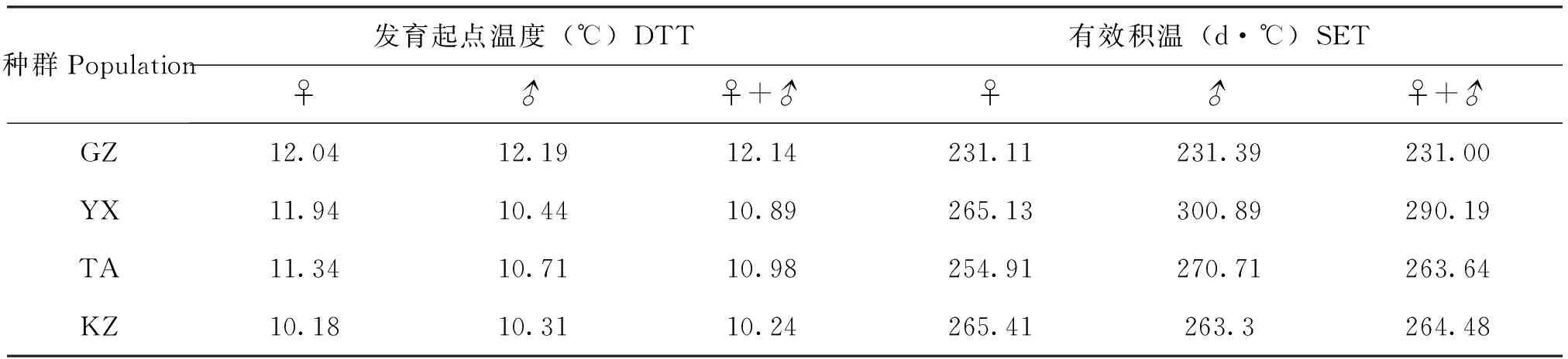
表1 不同地理种群棉铃虫幼虫发育起点温度和有效积温Table 1 DTT and SET of larva of different geographic populations of Helicoverpa armigera
从表2还可见,GZ、YX、TA和KZ种群非滞育蛹的有效积温分别为153.54、159.53、135.42和183.28日度,雌、雄非滞育蛹有效积温均随栖息地纬度升高而呈现出增加的趋势,呈不显著的正相关关系(雌蛹相关性方程:y=0.9061x+ 116.52,R2= 0.1398,P>0.05;雄蛹相关性方程:y=1.0335x+ 137.79,R2= 0.0864,P>0.05)。
除TA种群雌、雄非滞育蛹DTT基本相同外,其余种群非滞育蛹DTT均为雌性略高于雄性,而SET则所有种群均雄性高于雌性。
2.3 滞育蛹发育起点温度和有效积温
从表2可知,YX、TA和KZ种群滞育蛹的发育起点温度分别为17.51℃、16.50℃和16.74℃,均高于相应的非滞育蛹,分别高出5.05℃、3.39℃和5.50℃;随纬度升高,雌、雄滞育蛹发育起点温度均呈下降趋势,雌性呈不显著的负相关(y=-0.027x+ 17.387,R2=0.0486,P>0.05),雄性却呈显著负相关(y=-0.1096x+ 21.094,R2=0.9793,P<0.05)。
YX、TA和KZ种群滞育蛹的有效积温分别为195.05、269.46和349.17日度,大大高于相应非滞育蛹,分别高出35.56、134.04和165.89日度;雌、雄滞育蛹有效积温均随栖息地纬度升高而显著增加,呈显著正相关(雌滞育蛹回归方程:y=10.421x-83.107,R2=0.9303,P<0.05;雄滞育蛹回归方程:y=14.447x-250.99,R2=0.9331,P<0.05)。
滞育蛹DTT,YX、TA种群雄性高于雌性,KZ种群两性间几乎无差异;滞育蛹SET,YX、TA种群雌性高于雄性,KZ种群由相反。
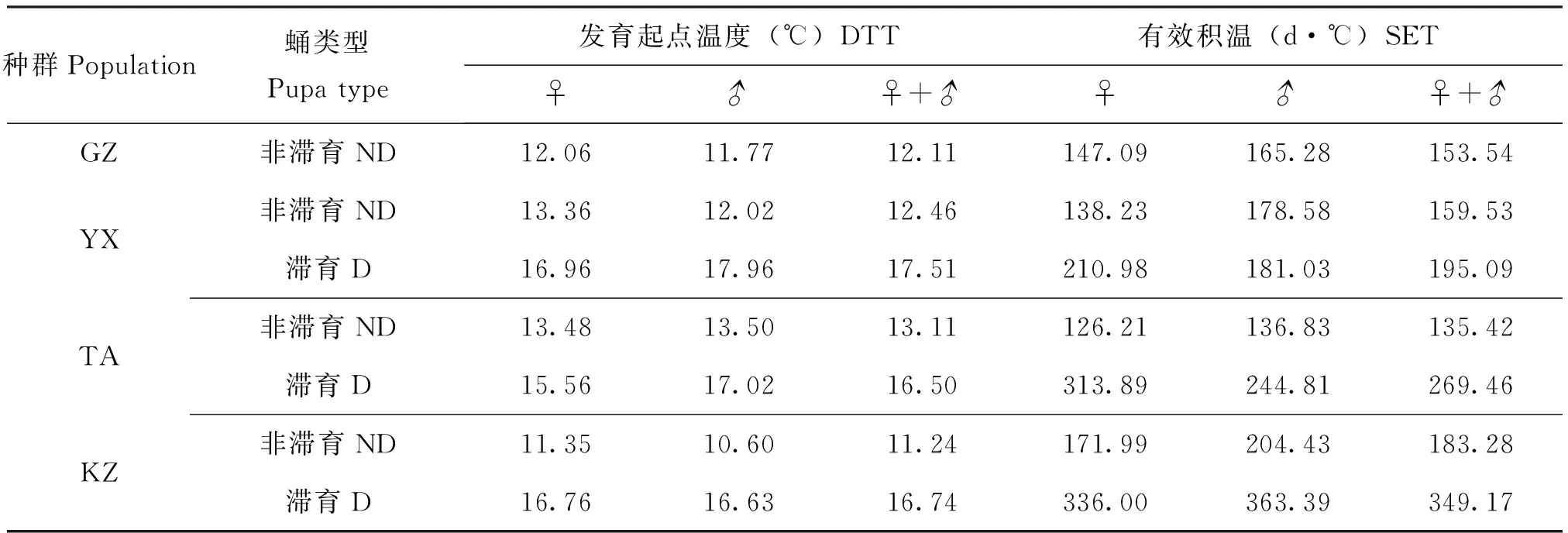
表2 不同地理种群棉铃虫蛹发育起点温度和有效积温Table 2 PupalDTT and SET of different geographic populations of Helicoverpa armigera
3 结论与讨论
本研究结果显示,棉铃虫发育起点温度和有效积温均存在地理变异,随栖息地纬度的升高,幼虫、非滞育蛹和滞育蛹的发育起点温度呈现逐渐下降趋势,幼虫的发育起点温度与栖息地纬度呈显著负相关关系(P<0.05),其余呈不显著负相关(P>0.05),而有效积温则相反,呈现上升趋势,滞育蛹有效积温与栖息地纬度升高呈显著正相关(P<0.05),其余呈不显著正相关(P>0.05)。棉铃虫发育起点温度和有效积温的地理变异,符合绝大多数昆虫的变异趋势,即LDT随栖息地纬度升高而降低,SET则相反(Honěk, 1996)。这说明高纬度北方种群幼虫、蛹的发育过程中温度较低也能开始并完成发育, 而低纬度南方种群幼虫、蛹的发育过程中要求温度较高才能开始并完成发育。
滞育蛹与非滞育蛹的发育特性有明显的差别,就发育起始温度来讲,滞育蛹发育起始温度高于非滞育蛹。Wilson等(1979)研究表明,滞育蛹的发育起始温度较非滞育蛹高5℃左右;蒋明星等(1997)发现,越冬蛹眼点移动前的发育特性明显不同于非滞育蛹,发育起始温度前者较后者高4℃-5℃,而在眼点移动后期,两类蛹则比较相似。吴孔明等(1996)的研究也发现滞育蛹发育起点温度比非滞育蛹高4℃以上。本试验与上述结果相似,滞育蛹的发育起点温度比非滞育蛹高3.39℃-5.50℃,其有效积温比非滞育蛹高35.56-165.89日度,在16℃以上滞育蛹才能启动发育,且北方种群需要累积的温度量要远高于南方种群,所以北方种群越冬代羽化时间远晚于南方种群。
有关棉铃虫发育起点温度和有效积温的研究已有一些零星报道,见表3。由表3可见,由于各人的研究方法、实验条件、食料以及虫源不同,所得结果差异很大。造成这些差异的主要原因,除了可能存在的棉铃虫地理变异外,还与食料及温度条件密切相关。如吴孔明等(1994)报道,同是河南新乡的棉铃虫且实验条件相同,幼虫分别饲于人工饲料和棉叶,得到的幼虫、蛹LDT和SET却存在很大差异(表3),说明幼虫期营养是影响棉铃虫有效积温的主要因素(吴孔明和郭予元,1994);日本熊本种群的LDT和SET与日本冈山种群的不同(Qureshietal., 1999),主要是由于所用的食料不同所致,而不是由于DTT的地理变异引起(Jallow and Matsumura, 2001);同是河北饶阳种群,由于食料和温度不同,吴坤君和李超所获得的结果却差异很大(表3);本实验的山东泰安种群(36.15° N)幼虫、非滞育蛹和滞育蛹DTT分别为10.98℃、13.11℃和16.50℃,SET分别为263.64、135.42和269.46日度,这与吴孔明等报道的饲于饲料的河南新乡种群(35° N)结果相近(滞育蛹DTT为16.52℃)(吴孔明和郭予元,1994,1996),而与纬度相近的却饲于棉花的山东临清种群(36.68° N)(牟吉元等,1995)差异较大,这进一步说明,食料及营养条件对对棉铃虫LDT及SET影响极大。此外,实验的温度范围不同,所得DTT估值可能也不同,应用线性回归方法推测DTT、SET时,应尽量选择4个以上的恒温,且温度范围要广(Bergant and Trdan, 2006)。本实验所采用的温度范围是20℃-28℃,设置20℃、22℃、25℃和28℃4个恒温,所有种群均在相同饲料等条件下进行的,故所得的结果是充分可信可靠的,完全可以以此比较种群的地理差异。
对发育起点温度DTT的估计有不同的方法。林郁等(1959)曾用实测法、图测法和回归直线法估计了三化螟SchoenobiusincertellusWalker的发育起点温度DTT和有效积温SET,发现计算所得数值都与实测的有一定差异,而且有些计算数值例如卵的LDT是没有实际意义的。有关棉铃虫LDT和SET的估算,国内学者多是根据有效积温法则公式和最小二乘方求系数公式求得LDT和SET常数(牟吉元等,1995),或者依据有效积温的“直接最优法”(吴孔明和郭予元,1994),而国外学者多采用温度与发育速率(发育历期的倒数)的线性回归法(Qureshietal., 1999; Jallow and Matsumura, 2001; Bartekova and Praslicka, 2006),这是一种简便的分析温度对昆虫发育的影响的方法,在昆虫生理温度范围内,这种方法适合于绝大多数昆虫,具有广泛实用性(Lamb, 1992;Honěk, 1996;Trudgilletal., 2005)。本实验所采用的是后面一种方法。
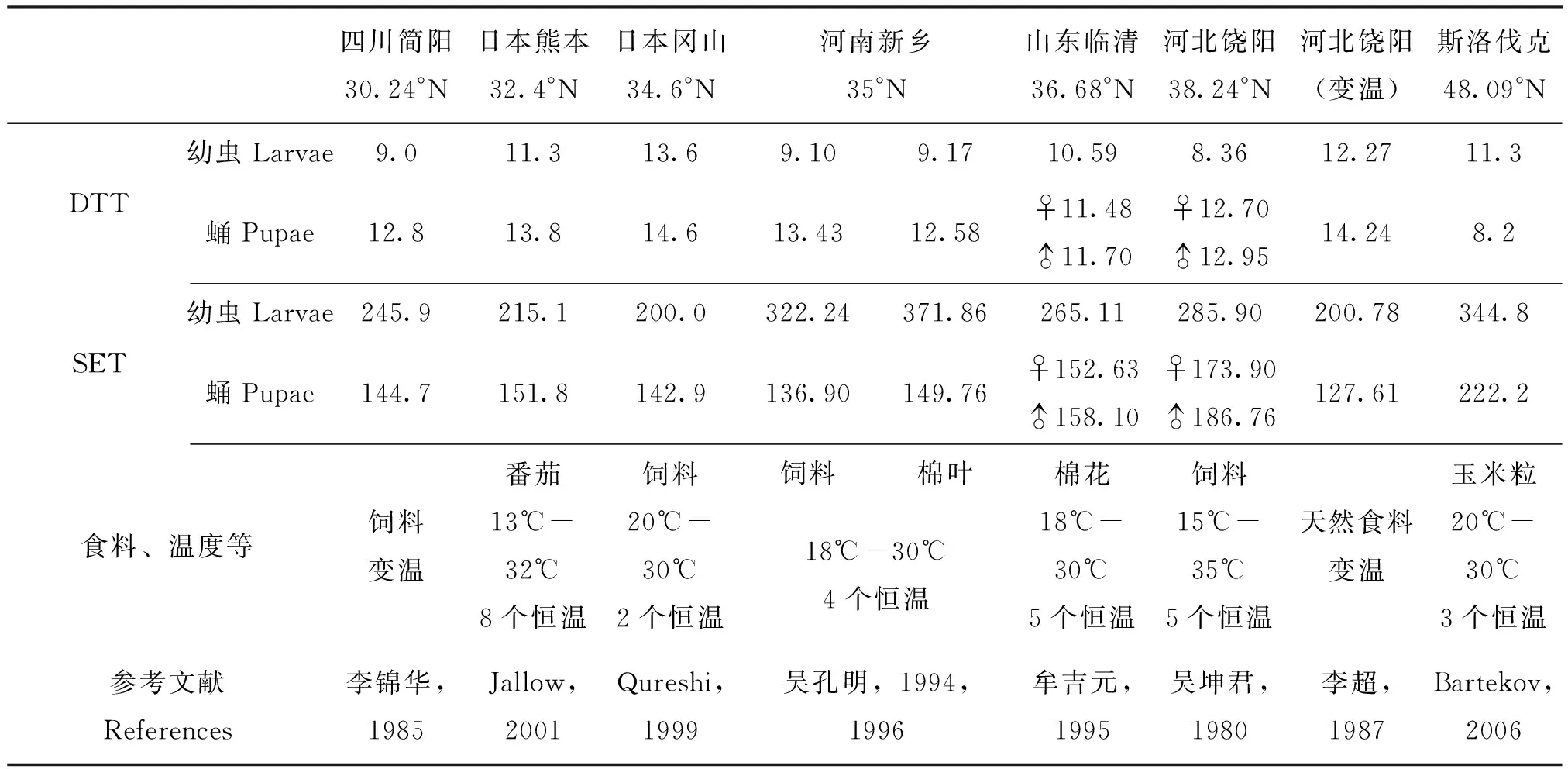
表3 棉铃虫的发育起点温度和有效积温(文献资料)Table 3 DTT and SET of Helicoverpa armigera (Literatures)
References)
Baker CRB, Miller GW.The effect of temperature on the post-diapause development of four geographical populations of the European cherry fruit fly (Rhagoletiscerasi) [J].EntomologiaExperimentalisetApplicata, 1978, 23: 1-13.
Bartekova A, Praslicka J.The effect of ambient temperature on the development of cotton bollworm (HelicoverpaarmigeraHübner, 1808) [J].PlantProtecionScience, 2016, 42: 135-138.
Bergant K, Trdan S.How reliable are thermal constants for insect development when estimated fromlaboratory experiments ? [J].EntomologiaExperimentalisetApplicata, 2006, 120: 251-256.
Casimero VR, Tsukuda F, Nakasuji,etal.Effect of larval diets on the survival and development of larvae in the cotton bollworm,HelicoverpaarmigeraHübner (Lepidoptera: Noctuidae) [J].AppliedEntomologyandZoology, 2000, 35: 69-74.
Chen YS, Tu XY.Geographic variation in developmental duration of the cotton bollworm,HelicoverpaarmigeraHübner (Lepidoptera: Noctuidae) [J].JournalofEnvironmentalEntomology, 2016, 38 (4): 771-778.[陈元生, 涂小云.棉铃虫发育历期的地理变异[J].环境昆虫学报, 2016, 38 (4): 771-778]
Cullen JM, Browning TO.The influence of photoperiod and temperature on the induction of diapause in pupae ofHeliothispunctigera[J].JournalofInsectPhysiology, 1978, 24 (8-9): 595-601.
Ferenz H.Anpassungen vonPterostichusnigritaF.(Col., Carab.) an Subarctische Begingungen [J].Oecologia, 1975, 19: 49-57.
Groeters FR.Geographic conservatism of development rate in the milkweed oleander aphid,Aphisnerii[J].ActaOecologica, 1992, 13: 649-661.
Honěk A.Geographical variation in thermal requirements for insect development [J].EuropeanJournalofEntomology, 1996, 93: 303-312.
Jallow MFA, Matsumura M.Influence of temperature on the rate of development ofHelicoverpaarmigera(Hübner) (Lepidoptera : Noctuidae) [J].AppliedEntomologyandZoology, 2001, 36 (4): 427-430.
Jallow MFA, Matsumura M, Suzuki Y.Oviposition preference and reproductive performance of JapaneseHelicoverpaarmigera(Hübner) (Lepidoptera: Noctuidae) [J].AppliedEntomologyandZoology, 2001, 36: 419-426.
Jarosik V, Honěk A, Dixon AFG.Developmental rate isomorphy in insects and mites [J].TheAmericanNaturalist, 2002, 160: 497-510.
Jiang MX, Zhang XX.Diapause termination and postdiapause development of overwintering pupae of cotton bollworm in Nanjing [J].ActaEntomologicaSinica, 1997, 40 (4): 366-373.[蒋明星, 张孝羲.南京地区棉铃虫越冬蛹滞育的解除与发育[J].昆虫学报, 1997, 40 (4): 366-373]
Lamb RJ.Developmental rate ofAcyrthosiphonpisum(Homoptera: Aphididae) at low temperatures: Implications for estimating rate parameters for insects [J].EnvironmentalEntomology, 1992, 21: 10-19.
Lamb RJ, Mackay PA, Gerber GH.Are development and growth of pea aphids,Acyrthosiphonpisum, in North America adapted to local temperatures ? [J].Oecologia, 1987, 72: 170-177.
Lee D,Spence J.Developmental adaptation of European corn borer (OstrinianubilialisHebner) in Alberta [J].CanadianEntomologis, 1987, 119: 371-380.
Li C, Li SQ, Guo BF.Studies on the temperature threshold of cotton bollworm development in varying temperature environments [J].ActaEntomologicaSinica, 1987, 30 (3): 253-258.[李超, 李树清, 果保芬.棉铃虫在变温环境中发育起点温度的研究[J].昆虫学报, 1987, 30 (3): 253-258]
Li JH, Li ZX.A study on the cotton bollworm,Heliothisarmigera(Hübner), in Sichuan[J].ActaPhytophylacicaSinica, 1985, 12(1): 17-24.[李锦华, 李兹新.四川棉区棉铃虫发生规律的研究[J].植物保护学报, 1985, 12 (1): 17-24]
Liang GM, Tan WJ, Guo YY.Technical improvement of cotton bollworm in artificial rearing [J].PlantProtection, 1999, 25 (2): 15-17.[梁革梅, 谭维嘉, 郭予元.人工饲养棉铃虫技术的改进[J].植物保护, 1999, 25 (2): 15-17]
Lin Y, Zhu ZQ, Hu JQ,etal.Studies on the predication of the outbreak of the paddy borer,SchoenobiusincertellusWLK., with the effective thermal summation [J].ActaEntomologicaSinica, 1995, 9 (5): 423-435.[林郁,祝兆麒, 胡家骐, 等.三化螟有效积温的研究Ⅰ: 各虫态期的发育零点和其有效积温[J].昆虫学报, 1995, 9 (5): 423-435]
Mu JY, Ciu L, Chen T.Testing biological constants of generation of the cotton bollworm,HeliothisarmigeraHübner, by the method of “thermal sums” [J].JournalofShandongAgriculturalUniversity, 1995, 26 (3): 280-284.[牟吉元, 崔龙, 陈天业.棉铃虫发育始点、有效积温的测定和发生检验[J].山东农业大学学报, 1995, 26 (3): 280-284]
Nechols JR, Tauber MJ, Tauber CA.Geographical variability in ecophysiological traits controlling bormancy inChrysopaoculata(Neuroptera: Chrysopidae) [J].JournalofInsectPhysiology, 1987, 33: 627-633.
Phoofolo MW, Obrycki JJ.Comparative life history studies of neartic and palearctic populations ofCoccinellaseptempunctata(Coleoptera: Coccinelliadae) [J].EnvironmentalEntomology, 1995, 24: 581-587.
Qureshi MH, Murai T, Yoshida H,etal.Effects of photoperiod and temperature on development and diapause induction in the Okayama population ofHelicoverpaarmigera(Hb.) (Lepidoptera Noctuidae) [J].AppliedEntomologyandZoology, 1999, 34 (3): 327-331.
Rae DJ, De’ath G.Influence of constant temperature on development, survival and fecundity of sugarcane mealybug,Saccharicoccussacchari(Cockerell) (Hemiptera: Pseudococcidae) [J].AustralianJournalofZoology, 1991, 39 (1): 105-122.
Royer TA, Giles KL, Kindler SD,etal.Developmental response of three geographic isolates ofLysiphlebustestaceipes(Hymenoptera: Aphididae) to temperature [J].EnvironmentalEntomology, 2001, 30: 637-641.
Shimizu K, Fujisaki K.Geographic variation in diapause induction under constant and changing conditions inHelicoverpaarmigera[J].EntomologiaExperimentalisetApplicata, 2006, 121 (3): 253-260.
Tauber CA, Tauber MJ, Nechols JR.Thermal requirements for development inChrysopaoculata: A geographically stable trait [J].Ecology, 1987, 68: 1479-1487.
Telfer MG, Hassall M.Ecotypic differentiation in the grasshopperChorthippusbrunneus: Life history varies in relation to climate [J].Oecologia, 1999, 121: 245-254.
Trudgill DL.Why do tropical poikilothermic organisms tend to have higher threshold temperature for development than temperate ones? [J].FunctionalEcology, 1995, 9: 136-137.
Umeya K, Yamada H.Threshold temperature and thermal constants for development of the diamond-back moth,PlutellaxylostellaL., with reference to their local differences [J].JapaneseJournalofAppliedEntomologyandZoology, 1973, 17: 19-24.[梅谷献二, 山田偉雄.コナガの発育零点と発育有効積算温量,およびその地理的差異 [J].日本応用動物昆虫学会誌(応動昆), 1973, 17 (1): 19-24]
Wilson AGL, Lewis T, Cunningham RB.Overwintering and spring emergence ofHeliothisarmigera(Hübner)(Lepidoptera: Noctuidae) in the Namoi Vallay, Newsouth Wales [J].BulletinofEntomologicalResearch, 1979, 69: 97-109.
Wu KJ, Gong PY.A new and practical artificial diet for the cotton bollworm[J].EntomologiaSinica, 1997, 14 (3): 277-282.
Wu KM, Guo YY.Investigations on the migration and diapause inHelicoverpaarmigera-Diapause responses of different geographical populations of cotton bollwormHelicoverpaarmigeraHübner [J].AgriculturalSciencesinChina, 1997, 30 (3): 1-6.[吴孔明, 郭予元.棉铃虫迁飞与滞育的研究-我国各棉区棉铃虫滞育诱导的光温反应特点[J].中国农业科学, 1997, 30 (3):1-6]
Wu KM, Guo YY.Investigations on the migration and diapause inHelicoverpaarmigera-Diapausetermination and emergence pattern inHelicoverpaarmigera[J].ScientiaAgriculturaSinica, 1996, 29 (1): 15-20.[吴孔明, 郭予元.棉铃虫迁飞与滞育的研究-棉铃虫滞育的解除与羽化形式[J].中国农业科学, 1996, 29 (1): 15-20]
Wu KM, Guo YY.The influence of nutrition on thermal constants for development of cotton bollworm [J].PlantProtection, 1994, 20 (4): 16-17.[吴孔明, 郭予元.营养对棉铃虫发育积温的影响.植物保护, 1994, 20 (4): 16-17]
Wu KQ, Chen YP, Li MH.Influence of temperature on the growth of laboratory population of the cotton bollworm,Heliothisarmigera(Hübner) [J].ActaEntomologicaSinica, 1980, 23 (4): 358-367.[吴坤君, 陈玉平, 李明辉.温度对棉铃虫实验种群生长的影响 [J].昆虫学报, 1980, 23(4): 358-367]
Xia X, Cong B, Song LQ,etal.Studies on the effective accumulated temperature and survival rate of Asian corn borer,Ostriniafurnacalis(Guenée) with different geographical populations and different voltinism [J].JournalofAnhuiAgriculturalSciences, 2007, 35 (19): 5795, 5797.[夏新, 丛斌, 宋立秋, 等.不同地理种群和不同化性亚洲玉米螟的有效积温和存活率的研究[J].安徽农业科学, 2007, 35 (19): 5795, 5797]
Geographic variation indevelopment threshold temperature and sum of effective temperatures of the cotton bollworm,HelicoverpaarmigeraHübner (Lepidoptera: Noctuidae)
CHEN Yuan-Sheng*, LUO Zhi-Di, ZHONG Ping-Hua
(Jiangxi Environmental Engineering Vocational College, Ganzhou 341000, Jiangxi, Province, China)
The purpose of the research is to understanding geographic variation in development threshold temperature (DTT) and sum of effective temperatures (SET) of different populations of the cotton bollworm,HelicoverpaarmigeraHübner, in order to suppose its possible occurrence generations in different regions as the basis for predicting the occurrence period of its development., thus to establish IPM method to control this pest effectively.After breeding the larvae ofH.armigerafor one generation from 4 different geographic populations (Guangzhou population, 23.08°N, 113.14°E; Yongxiu population, 29.04°N, 115.82°E; Taian population, 36.15°N, 116.59° E; Kazuo, 41.34°N, 120.27°E), on the basis of its developmental durations under constant temperature 20℃, 22℃, 25℃, and 28℃, and light period L16:D8, the DTT and SET of larvae, non-diapaused pupae and diapaused pupae ofH.armigerawas systematically investigated, and the relationship between the DTT, SET and latitude was analyzed.The results showed that the DTT of larvae, non-diapaused pupae and diapaused pupae was 10.18℃-12.19℃, 10.60℃-13.50℃ and 15.56℃-17.96 ℃, respectively.Their SET were 254.91-300.89, 126.21-204.43 and 181.03-363.39 day-degrees, respectively, reared in the laboratory with artificial feed.The DTT of diapaused pupae was 3.39℃-5.50℃ more than that of non-diapaused pupae, and their SET were 35.56-165.89 day-degrees more than that of non-diapaused pupae.There was clear geographic variation in the DTT and SET ofH.armigera, with the increase of latitude habitat.The DTT of larvae, non-diapaused pupae and diapaused pupae was decreased, showing negative correlation, but the SET were the opposite, SET increased with increasing geographical latitude.
Helicoverpaarmigera; development threshold temperature; sum of effective temperatures; geographic variation; latitude; geographic population
陈元生,罗致迪,钟平华.棉铃虫发育起点温度和有效积温的地理变异[J].环境昆虫学报,2017,39(3):573-579.
国家自然科学基金项目(31060243);江西省科技支撑计划重点项目(20151BBF60069)
陈元生,男,1967年生,江西信丰人,教授,博士,从事昆虫生物学和滞育生理生态研究,E-mail: cys0061@163.com
Received: 2016-03-14; 接受日期Accepted: 2016-10-24
Q965;S433.3
A
1674-0858(2017)03-0573-07
猜你喜欢
中国农业科学(2022年15期)2022-08-09 02:23:48
中国马铃薯(2022年2期)2022-07-05 00:12:08
四川农业科技(2019年10期)2020-01-06 03:02:02
黑龙江粮食(2019年10期)2019-11-23 02:09:32
齐鲁周刊(2017年29期)2017-08-08 06:28:15
新疆农垦科技(2016年2期)2016-08-21 13:50:22
现代农业(2016年5期)2016-02-28 18:42:35
现代农业(2016年4期)2016-02-28 18:42:09
现代农业(2015年5期)2015-02-28 18:40:42
技术经济与管理研究(2014年11期)2014-03-11 17:02:54
