
开口饵料对仔稚鱼相关生理影响研究进展
2017-07-31 18:44:54杜强张竹青周路于海波冯浪
中国饲料 2017年13期
杜强,张竹青*,周路,于海波,冯浪
(1.贵州省水产研究所,贵州贵阳550025;2.贵定县农村工作局渔业科,贵州贵定551300;3.罗甸县渔政渔港监督管理站,贵州罗甸550100)
开口饵料对仔稚鱼相关生理影响研究进展
杜强1,张竹青1*,周路1,于海波2,冯浪3
(1.贵州省水产研究所,贵州贵阳550025;2.贵定县农村工作局渔业科,贵州贵定551300;3.罗甸县渔政渔港监督管理站,贵州罗甸550100)
随着水产养殖业规模的扩大,苗种的成活和生长率变得至关重要。开口饵料作为水产苗种发展的决定性因素已成为近年来研究的热点。因此,系统地研究开口饵料对仔稚鱼消化生理的调控对于保证水产养殖业的快速发展具有重要作用。本文就开口饵料类型以及其对仔稚鱼消化生理、组织学及后期代谢的影响等方面进行了综述。
开口饵料;仔稚鱼;存活生长
我国作为水产品养殖业发展最快的国家之一,据中国渔业统计年鉴(2016)数据,2015年全国水产苗种产值617亿元,同比增长3.2%,而苗种作为养殖渔业发展的基础,对水产业发展具有重要作用。在育苗初期是否能及时提供营养均衡、大小适宜的饵料,对鱼类幼苗成活效率有很大影响。
本文将从仔稚鱼开口特征、开口饵料类型及开口饵料对仔稚鱼消化生理及系统组织学的影响等方面对仔稚鱼开口饵料的研究进行综述。
1 仔稚鱼开口特征
仔稚鱼消化生理研究是幼体开口饵料研究的基础。鱼类苗种开口摄食一般有两个重要的生理学特征:(1)开口与泄殖孔的孵化成型及消化器官的整体串联,不同鱼类完成顺序存在差异(表1)。(2)功能性鳔的生成。鳔的生成充气为鱼提供了浮力调控能力,因而鱼类能够漂浮在水体中,减少能耗为进食生长保留更多的能量。
2 仔稚鱼开口饵料类型
常见鱼类开口饵料类型及特点见表2,主要有生物饵料和人工开口饵料两大类。
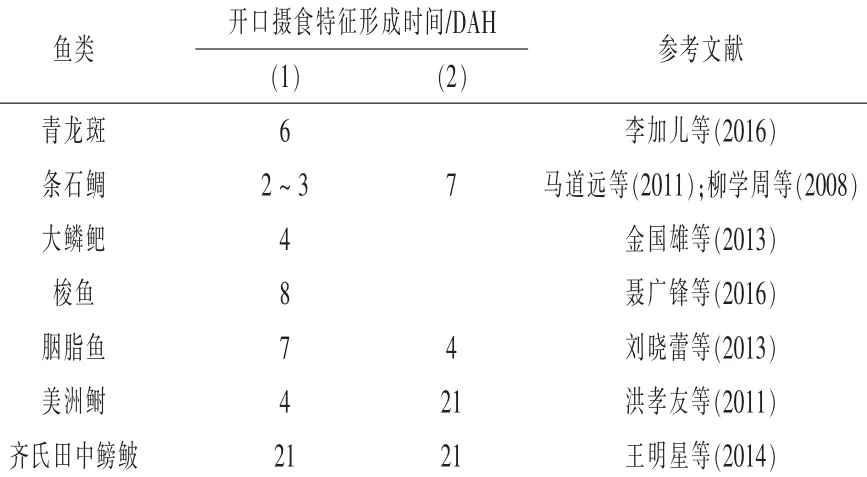
表1 不同鱼类开口摄食特征研究
由于生物饵料具有较好的诱食性,小规模的幼鱼培育有较好的成活率,所以生物饵料常被用于传统的苗种培育。然而随着集约化规模养殖的发展,生物饵料生产量受人力、物力和生产设施的限制,以及来自环境方面的影响,生物饵料价格居高不下。另外在营养组成方面也存在一定短板,如生物饵料中缺乏仔稚鱼大量需要的多不饱和脂肪酸(HUFA)。同时由于生物饵料活体容易携带病原菌进入育苗水体,增大育苗风险。因此生物饵料反而成为了仔稚鱼大规模培育的限制因素。

表2 常见鱼类开口饵料
人工微颗粒饲料最早开始用作水产动物早期的开口饵料是为了替代生物饵料,增加水产动物大规模培育的效果(吴格天,2006)。
由于水产饵料使用环境的特殊性,人工微颗粒饲料需要具备以下特点。(1)要在水体中能保持一定的稳定状态,以防止饵料在水中营养流失;(2)饵料营养组成能满足稚仔鱼生长所需(Langdon,2003);(3)饵料颗粒大小要适合稚仔鱼口径以便消化吸收。因此,平衡人工饲料中营养物质的浸出与饵料颗粒的消化性是一个有待解决的问题(Hamre等,2013)。所以到目前能完全使用人工微颗粒饵料开口的稚仔鱼相对较少。
3 开口饵料对仔稚鱼存活和生长的影响
由于仔稚鱼口裂对饵料颗粒的限制(Russ等,2009),仔稚鱼可能因为无法摄食较大饵料而发生饥饿胁迫。因此饵料大小是鱼类幼苗选择的主要因素(刘建国等,2007)。
不同种类稚仔鱼对饵料大小的选择也存在一定差异。金钱鱼(Scatophagus argus)在5DAH时的口裂宽仅有0.17 mm,32DAH时发育到0.5 mm(蔡泽平,2015);而斑鳜(Siniperca scherzeri)经过91 h的培育后开口即可摄食6 mm长的饵料鱼(胡振禧,2014)。同时稚仔鱼摄食饵料大小范围随着日龄增长而增大,而快速生长则意味着需要获取更有营养的饵料,以在饵料中获取的能量和捕获、消化饵料耗能之间维持稳定状态(Pradhan等,2014)。因此仔稚鱼通常会选择更大的的饵料。在蛇头纹鳢(Ophiocephalus)(Qin,1996)、非洲鲶鱼(Silurus asotus)(Baras,2014)仔鱼中也有类似的报道。而同种鱼类在同一时间也会有不同特性,如黄峰等(2000)大口鲇在2.5 DAH时口裂宽1.2 mm,而乔志刚等(2007)却发现,同一时间的大口鲇口裂宽仅0.837 mm,二者相差30%。这可能是同种鱼类幼苗由于孵化环境、营养条件等差异导致。
饵料密度对仔鱼摄食也有很大的影响,研究发现,在一定范围内仔鱼的摄食强度会随着饵料密度增加而增大。如南方鲶(Silurus meridionali)7~30DAH仔鱼在浮游生物密度2000个/L时生长最快,残食率最低(杨凯等,2010)。黄尾石首鱼(Seriola lalandi)的4日龄仔鱼摄食轮虫密度为22 ind/mL/h,11日龄时增大到22 ind/mL/h(Lindsey,2016)。而当轮虫密度超过30 ind./mL时,孵化后70 h花尾胡椒鲷(Plectorhynchus cinctus)仔鱼的摄食强度明显下降(张雅芝,2000)。因此饵料密度的控制是仔鱼能保持较高摄食效率的重要因素。
相较于幼鱼和成鱼阶段,稚仔鱼对饵料的营养要求更高(郭文学等,2014)。由于仔稚鱼对蛋白质、氨基酸、脂类、矿物质、维生素等营养物质的需求量随生长出现较大变化,所以在配制人工饲料时常采用鱼粉、豆粕、菜籽粕、畜禽粉等混合蛋白源来弥补单一蛋白源的营养不足。因此,仔稚鱼阶段饵料的选择会直接影响早期鱼类的存活和生长率。
4 开口饵料对仔稚鱼消化生理的影响
Ronnestad等(2013)根据胃出现的情况将早期仔鱼分为无胃鱼、早熟鱼类、晚成鱼三种。无胃鱼是像常见鲤科鱼如鲤、鲢、鳙、草鱼等整个生活史中都没有形成胃的鱼类(毕冰等,2011)。在初次摄食便发育出功能性胃的是早熟鱼类,如鲑鳟鱼类和罗非鱼属。晚成型鱼类的胃是在初次摄食后开始发育的,大部分海洋鱼类都是晚成型鱼类,由于胃的较晚发育成型,大多数淡水鱼较海水鱼能更好的接受人工开口饵料(宋青春,2010)。
肝胰脏作为仔鱼初孵阶段消化生理的重要器官,通过饵料刺激胰脏所分泌的酶类可以判断鱼体营养吸收状况(骆豫江等,2010;张永泉等,2010)。特别是在还没有胃的仔稚鱼期,鱼类所摄食的饵料主要通过肠道内胰腺酶类消化。
虽然仔稚鱼消化系统功能在开口初期尚未完善,但大多数消化酶已经在孵出鱼类的早期发育中逐渐产生(郑凯迪等,2010)。然而对于仔稚鱼如何控制其消化酶活性以满足不同条件需要仍存在分歧。一些研究认为,活饵刺激对仔稚鱼消化酶活性的影响几乎可以忽略不计(Martinez等,2014;Lazo等,2007),但也有试验表明,在仔稚鱼缺乏足够的消化酶时,可通过活饵中酶类物质帮助摄食鱼类消化食物,如刘忠优等(2014)发现投喂小球藻强化后的天然饵料对龙虎斑仔稚鱼的胃、肠蛋白酶活性有显著提高作用。
5 开口饵料对仔稚鱼消化系统组织学的影响
鱼类初次摄食外源性营养时,某些消化器官细胞会作出不同程度的应答反应,可通过组织学方法将这些应答反应作为评判稚仔鱼阶段生理和营养状态的指标(Pradhan等,2014)。在仔稚鱼阶段,胰腺、肝脏和肠道等消化组织和器官处于发育未完全阶段,它们的形态结构对突然改变的摄食条件和营养变化会产生应激反应(Cahu等,2009)。因此,组织学是用作研究营养缺乏或饥饿导致不可逆损伤的方法之一(张永泉等,2009)。如组织学观察发现对于乌苏里拟鲿(Pseudobagrus ussuriensis)仔鱼,人工饵料比天然饵料更易消化吸收(车建芳等,2016)。大黄鱼幼苗饲喂微颗粒饲料条件下肠道膜褶皱变深变多,而投喂桡足类的鱼肠道黏膜褶皱更易变平(于海瑞等,2012)。Gisbert等(2004)研究发现,在以生物饵料作为开口饵料的牙鲆(Paralichthys californicus)肠道中有大量脂肪的积累,同样的现象也出现在Bolasina(2006)对牙鲷幼鱼的研究中。这表明稚仔鱼类自身消化器官的发育会受到开口饵料的影响,而这种影响主要是通过长时间作用于消化器官的发育及代谢来共同完成的。
综上所述,开口饵料是稚仔期鱼类成活率及生长率的重要决定因素。因此,了解不同鱼类开口特征、消化生理和组织结构特点,选用和开发针对性的开口饵料是水产鱼苗系统工程的重中之重。
[1]毕冰,孙中武,肖晓文,等.鲤、鲢、鳙、草鱼消化道消化酶种类和活性的比较研究[J].水产学杂志,2011,24(2):17~20
[2]蔡泽平,胡家玮,王毅金.钱鱼早期发育的观察[J].热带海洋学报,2014,33(4):20~25.
[3]车建芳,卜宪勇,苏宝辉,等.不同饲料对乌苏里拟鲿仔鱼生长及饲料利用率的影响[J].中国饲料,2016,10:30~32.
[4]郭文学,尹家胜,佟广香,等.养殖方式、光照强度对哲罗鱼稚鱼生长与存活的影响[J].海洋与湖沼,2014,45(2):265~273.
[5]洪孝友,朱新平,陈昆慈,等.美洲鮒胚胎及仔稚鱼的发育[J].水生生物学报,35,1:153~162.
[6]胡振禧.斑嫩胚胎与仔鱼早期发育观察:[硕士学位论文][D].福建福州:福建农林大学,2014.
[7]黄峰,严安生,张桂蓉,等.大口鲇仔鱼消化道的组织学观察[J].华中农业大学学报,2000,19(1):59~63.
[8]金国雄,徐伟,耿龙武,等.大鳞鲃消化系统早期发育的组织学观察[J].水产科学,2013,32(6):311~315.
[9]李加儿,吴水清,区又君,等.斜带石斑鱼(♀)×鞍带石斑鱼(♂)杂交子代(青龙斑)消化系统的早期发育[J].动物学杂志,2016,51(1):73~83.
[10]刘建国,殷明焱,张京浦,等.微拟球藻的水产饵料效果研究[J].海洋科学,2007,31(5):4~9.
[11]刘晓蕾,余必先,蒲德永,等.胭脂鱼仔稚鱼的发育及生长研究[EB/OL].中国科技论文在线:http://www.paper.edu.cn/releasepaper/content/ 201303410.
[12]刘忠优,张健东,周晖,等.不同饵料强化剂对龙虎斑仔稚鱼生长、存活率、消化酶活力及体成分的影响[J].广东海洋大学学报,2014,34(4):27~32.
[13]柳学周,徐永江,王妍妍,等.条石鲷的早期生长发育特征[J].动物学报,2008,54(2):332~341.
[14]骆豫江,陈昆慈,朱新平,等.高体革(鱼刺)消化道发生的组织学观察[J].水产学报,2010,34(2):264~270.
[15]马道远,于道德,肖永双,等.条石鲷消化道发育的组织学观察[J].海洋科学,2011,35(12):28~35.
[16]梅高峰.卤虫的营养价值及营养强化[J].养殖技术顾问,2014,1:35.
[17]聂广锋,李加儿,区又君,等.梭鱼仔、稚、幼鱼消化系统胚后发育的组织学观察[J].中国水产科学,2016,23(1):90~103.
[18]乔志刚,石灵,常国亮,等.鲇胚胎及其仔鱼发育的连续观察[J].水产科学,2007,26(8):431~435.
[19]宋青春,齐遵利,水产动物营养与配合词料学[M]:中国农业大学出版社,2010.
[20]王明星,钟俊生,石锅寿宽,等.齐氏田中鳑鲏的早期发育研究[J].上海海洋大学学报,2014,23(3):382~387.
[21]吴格天,徐子伟.水产动物开口饲料的研究进展[J].饲料工业2006,27(4):28~31.
[22]许友卿,李伟峰,丁兆坤.多不饱和脂肪酸对鱼类免疫与成活的影响及机理[J].动物营养学报,2010,22(3):551~556.
[23]许友卿,庄丽,丁兆坤.多不饱和脂肪酸对海水仔稚鱼生长发育的影响及机理[J].饲料工业,2010,31(14):13~18.
[24]杨凯,樊启学,周华,等.饵料密度对南方鲶仔稚鱼生长、成活、残食的影响[J].应用与环境生物学报,2010,16(2):235~239.
[25]于海瑞,艾庆辉,麦康森,等.微颗粒饲料与冷藏桡足类对大黄鱼稚鱼消化酶活力、肠道肝脏显微结构的影响[J].水生生物学报,2012,36(6):1087~1096.
[26]俞爱萍.微生态制剂及在水产养殖上应用研究[J].中国水产,2015,12:76~78.
[27]张金平,刘远高,杨军,等.齐口裂腹鱼人工育苗技术的初步研究[J].浙江海洋学院学报(自然科学版),2015,34(4):318~322.
[28]张雅芝,谢仰杰,张文生.花尾胡椒鲷早期发育阶段的摄食与生长特性[J].台湾海峡,2000,19(1):27~35.
[29]张永泉,刘奕,尹家胜,等.哲罗鱼消化系统胚后发育的形态与组织学的研究[J].海洋与湖沼,2010,41(3):422~428.
With the expansion of aquaculture,the survival and growth rate of seedling become critical.Initial feeding as decisive factors of aquatic fingerlings were paid more attention.Therefore,systematic research on the initial feeding regulation of digestive physiology plays an important role for the fast-growing aquaculture.In this paper,the initial feeding type,the effects of initial feeding on the larval digestive physiology,tissue and the metabolism in later life of fish were reviewed.
initial feeding;larval fish;the survival and growth
S963
A
1004-3314(2017)13-0042-04
10.15906/j.cnki.cn11-2975/s.20171311
*通讯作者
猜你喜欢
渔业研究(2022年5期)2022-10-28 01:56:14
水产科学(2022年1期)2022-01-26 08:43:00
渔业科学进展(2021年6期)2022-01-04 06:26:42
当代水产(2021年1期)2021-03-19 05:17:14
当代水产(2019年9期)2019-10-08 08:02:44
甘肃畜牧兽医(2019年2期)2019-03-20 01:49:42
海洋渔业(2016年6期)2016-04-16 03:00:32
广东海洋大学学报(2015年4期)2016-01-13 08:39:40
应用海洋学学报(2015年3期)2015-11-22 07:39:22
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:34
