
濒危植物暗紫贝母的开花物候和传粉方式
2017-07-30 11:29郭凤霞张荩凤
草地学报 2017年6期
郭凤霞, 陈 垣, 张荩凤, 白 刚
(1.甘肃农业大学甘肃省中药材规范化生产技术创新重点实验室,甘肃省药用植物栽培育种工程研究中心, 甘肃省干旱生境作物学重点实验室,生命科学技术学院,农学院,兰州 730070; 2.甘肃天士力中天药业有限责任公司, 甘肃省特色药用植物资源保护与利用工程实验室,甘肃省特色药材规范化可追溯栽培工程技术研究中心,定西 748100)
暗紫贝母(Fritillariaunibracteata)为药材川贝母的基原植物之一,以干燥鳞茎入药,夏、秋二季或积雪融化后采挖,清洗晒干或低温干燥后药用,味苦、甘,微寒,归肺、心经,具有清热润肺、散结消疽等功效,用于痰中带血、乳疽等症[1],其鳞茎中含有多种生物碱和微量元素等[2-3]。暗紫贝母野生于海拔3 100~4 500 m的高山或草原,是药材“松贝”的主要来源[4],我国主产于四川、甘肃、青海等高海拔草原地带[5-6]。甘肃主要分布在甘南州、渭源、漳县、岷县高寒山区,分布范围十分有限,野生资源极其稀少[7]。
川贝母基原植物属百合科(Liliaceae)贝母属(Fritillaria)多年生草本药用植物,适生环境气候冷凉,生长发育极其缓慢,年生长量很小,生长周期长,从种子播种至开花结果需5年及以上时间,生长4 年才可采挖药材,药材产量低[5-7]。药材主要依赖野生资源,现因采挖过度,加上人畜扰动加大,草地退化引起野生资源趋于枯竭[8-9],其中暗紫贝母、川贝母(F.cirrhosa)和甘肃贝母(F.przewalskii)均已成为国家二级濒危保护植物,由于药材市场对暗紫贝母认可度高,造成供不应求,价格昂贵,甘肃境内野生品已趋灭绝,资源的可持续性遭到威胁[7]。可持续主要指自然资源及其开发利用程度间的平衡[10]。自然条件下,暗紫贝母更新主要依赖地下根茎无性繁殖,因其种子具有较强的后熟和休眠特性,自然有性繁殖几率很低。野生驯化栽培时,多采用鳞茎繁殖,繁殖成本高,严重限制了规模化种植的推进,药材还主要依赖野生资源,远不能满足市场需求。利用种子驯化栽培无疑成为保护生态环境,提高野生资源再生性的有效途径[11]。国内外对百合科植物开花物候和繁育特性开展了大量工作。Duffyh和Johnson发现[12],雌雄同花百合科植物Bulbineabyssinica去除花药和花丝后蜜蜂很少访花,几乎不结种子。Shoichi等[13]调查显示,日本岛屿本州中心沿海岸线低洼区分布一种百合科贝母属植物(F.koidzumiana),属典型春季短命落叶林,多年多次开花,子房结实率达75%。我国研究发现海拔高度影响暗紫贝母生物量分配[4],也影响花朵和果实的大小及种子的质量[8-9],已通过研究探明了甘肃贝母种子灌浆和发芽特性[5-6],并探寻出破除休眠促进后熟,提高种子发芽率的关键技术[14-15]。陈垣等[7]研究发现,暗紫贝母花药开裂与强柱头可受性同步,柱头活力持续期更长,判断有性繁育系统应属于部分自交亲和异交需传粉者。
上述研究对选择川贝母驯化栽培适宜生态区和种子繁育策略提供了重要参考,但至今对暗紫贝母等川贝母基原植物的传粉结实特性的系统研究尚少见报道,野生环境下暗紫贝母零星分布在灌木丛或草丛中,很难找到足量的暗紫贝母开花居群定点进行相关研究。开花物候和传粉方式是反映植物有性生殖更新能力的重要依据[16-18],因此,在适宜生态区对野生驯化栽培的暗紫贝母物候及授粉结实特性的深入研究具有重要意义,可为制定川贝母基原植物野生驯化种子繁育规程提供科学依据,促进暗紫贝母资源的可持续化利用和发展。
1 材料与方法
1.1 试验区概况与研究材料
试验区位于甘肃省定西市漳县南部山区石川乡甘肃农业大学川贝母基地(34°35′29″ N,104°19′41″ E),东连岷县蒲麻镇,西靠四族乡,南与岷县禾驮乡接壤,北接草滩乡,属典型高寒阴湿气候。试区海拔3 100 m,无霜期126 d,年均降雨量500 mm,土层深厚、土壤肥沃,适宜暗紫贝母生长发育[7]。试验地位于坡度15°山坡上,占地面积2 500 m2。
暗紫贝母鳞茎2013年7月15-18日采自四川省红原县,分选出大鳞茎(经陈垣教授鉴定确认为4~5年生),带土保湿运回漳县。2013年7月20日在生荒地采用畦垄撒播,畦垄高5 cm,覆土厚度3 cm,播种后遮盖遮阳网。2014年返青后揭去遮阳网,仿野生栽培返青株经甘肃农业大学中草药栽培与鉴定系陈垣教授鉴定确认为暗紫贝母(Fritillariaunibracteata)。
1.2 开花物候动态观察
从鳞茎播种开始定期田间观察暗紫贝母返青及生长发育动态。2015年返青后从5月1日花蕾显现开始,田间选取暗紫贝母代表性植株20株挂牌标记,每隔3 d观察测定植株生长发育指标和花蕾大小、开花进程、开花顺序、单花结构、花粉散出和花朵萎蔫等生长发育动态,直到蒴果成熟采收为止,采用游标卡尺测定茎粗(stem diameter)、花冠长度(corolla length)和花冠直径(corolla diameter)等生长发育指标,茎粗测定部位为茎基部2 cm处,花冠直径测定部位为花冠上部开口处。每年返青后开花期观察地上部分植株形态变化和物候特征,田间观察统计株型结构和植株形态。
1.3 花朵操作及人工杂交授粉试验
暗紫贝母花蕾套袋杂交授粉操作试验于2015年5-7月在田间进行,花蕾操作处理试验采用单因素试验方案,完全随机设计,共设8个处理组,即(1)自然授粉对照(natural pollination CK):自然条件下不进行任何处理,用来检测自然结实率(rate of natural fruit-set),重复60朵花;(2)不去雄套纸袋(bagged without castration):自然条件下对花蕾采用硫酸纸袋套袋处理,以隔绝虫媒和风媒传粉机会,用于检测自动自交能力和自然自交亲和性(self-compatibility),重复30朵花;(3)不去雄套网袋(netted without castration):自然条件下对花蕾采用孔径1 mm的纱网套袋处理,以隔绝虫媒传粉机会,用于检测风媒传粉能力,重复30朵花;(4)去雄不套袋(non-bagged after castration):对花蕾去雄后不再进行其他处理,用以检测外来花粉的传粉效率,即自然异花授粉率,包括虫媒和风媒传粉成效,重复30朵花;(5)去雄套纸袋异株异花人工授粉(bagged and cross-pollinated after castration):对花蕾去雄后套纸袋,采集异株成熟花粉授粉,用于检测人工异花授粉率,重复75朵花;(6)去雄套网袋异株异花人工授粉(netted and cross-pollinated after castration):对花蕾去雄后套孔径1 mm的纱网,采集异株成熟花粉授粉,用于检测人工异花授粉率和风媒授粉能力,重复30朵花;(7)去雄套网袋(netted after castration):用于检测风媒传粉媒介效率,重复30朵花;(8)去雄套纸袋(bagged with paper after castration):用于检测是否存在单性结实或无融合生殖,重复30朵花。上述套袋处理花蕾均选择花被长度小于1.5 cm且花药尚未成熟的绿色花蕾,经显微镜检测后确定其花粉尚未成熟,每次操作前后用75%乙醇彻底消毒,以防杂花粉污染。授粉处理第7 d的18:00去除套袋,第10 d统计坐果数,种子成熟时采收统计结种子蒴果数和种子数,最后计算各处理结实率。
结实果率(%)=结种子蒴果数×100/总花数
1.4 数据分析
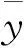
2 结果与分析
2.1 暗紫贝母开花物候和田间植株形态观测结果
图1为甘肃漳县栽培的暗紫贝母田间花蕾期套袋(图1A)和植株形态(图1B-H)。2013年7月采用野生大鳞茎播种的暗紫贝母次年3月下旬返青,90.0%以上植株单叶无茎,叶似飘带,俗称“一匹叶” (图1B-C),9.0%单株有叶2~5片,茎短缩,俗称“树儿子” (图1D),开花株稀少,单花顶生,7月上旬地上部分全部枯萎倒苗。仿野生农田环境条件下,随着栽培年限的延长,田间株形多样化趋势愈加明显(图2),2015年返青后,田间有大量“一匹叶”株(图1 B-C)和多叶未开花“树儿子”株(图1 D),“灯笼花”株明显增多(图1 B和图2),开花株5月上旬现蕾,下旬进入盛花期,6月中旬蒴果成形,7月中下旬种子成熟。开花株叶片线状披针形,底部叶对生,上部叶互生,无叶柄,叶片在5片及以上(图1 E-G)。开花株中,单株花朵数的年分配比例趋势更为明显,95.5%单株顶生1花(图1 F),4.5%单株抽生2花(图1 G)。2016年返青后,盛花期6月4日统计(图2),田间株形多样化,“一匹叶”株仍占据优势(1/2),“树儿子”株占1/3,开花株即“灯笼花”株占1/5,三者差异性达显著水平(P<0.05)。在开花株中,单株开花数存在极显著差异(P<0.01),2/3植株顶生1花,近1/3植株着生2花,1.0%的植株着生3花,花自上到下从叶腋处抽出,顶花柄也可分支花蕾,茎易折断。“一匹叶”株地下根茎部着生1枚鳞茎,多叶开花株可见2枚鳞茎,茎杆中空,鳞茎下部着生数量不等的根(图1 H)。“一匹叶”株和“树儿子”株7月上旬全部落黄倒苗,开花株7月中下旬蒴果成熟后地上部枯萎倒苗,茎秆中空。群体开花期从5月中旬持续至6月中旬,开花期田间观察到访花昆虫有蜂类、蜘蛛、蝇类、蝶类和蚂蚁,常在花被内外穿梭访花,但种类和数量较少。峰类主要是蜜蜂,初步鉴定为石蜜,体外黑色(图1F),活动频繁,传粉效率最高。蜘蛛主要是红蜘蛛,体小呈红色,穿行于花器官中。

图1 漳县驯化栽培的暗紫贝母试验田生境及开花期植株形态Fig.1 Experimental field surrounding and plant shape during anthesis for F. unibracteata domesticated in Zhang County(A) 试验地生境和套袋株Test field surrounding and the bagged plants;(B) 田间植株形态Plant shape in field; (C) 单叶株One-leaf plant;(D)多叶株Multiple-leaf plant; (E)现蕾株Budding plant; (F)单花株Single flower plant; (G) 多花株Multiple-flower plant; (H) 全株Full plant

图2 漳县驯化栽培暗紫贝母2015-2016年开花期田间单株叶片数和开花数植株分布Fig.2 Distribution of Leaf and flower numbers for single plant in F. unibracteata anthesis in Zhang County domesticated field during 2015-2016注:不同小写字母表示P<0.05差异显著,不同大写字母表示P<0.01差异极显著Note:Different lowercase letters indicate significant difference at the 0.05 level and the capital letters indicate significant difference at the 0.01 level
2.2 暗紫贝母现蕾开花期植株发育动态观测
2015年现蕾期(5月17日)选择20株定期观测显示(图3),进入开花结果期,暗紫贝母营养生长与生殖生长并进,随发育进程的递进植株呈极显著增高加粗趋势(P<0.001)。5月19日现蕾期较6月7日结果期植株平均增高5.13 cm(t=5.177,P<0.001) (图3A),增粗0.025 cm(t=4.387,P<0.001) (图3B)。叶片极显著加宽和加长(P<0.001)(图3C-D),并表现出个体间的极显著差异性(P<0.001)。5月19日田间株高和茎粗变幅分别为10.18~19.26 cm和0.164~0.242 cm,变异系数分别17.50%和9.13%。叶长和叶宽变幅分别为2.110~3.850 cm和0.160~0.550 cm,变异系数分别为17.56%和34.14%。至6月7日株高和茎粗变幅达15.06~26.72 cm和0.192~0.268 cm,叶长和叶宽变幅分别为2.680~4.920 cm和0.216~0.680 cm。现蕾开花期,株高、茎粗、叶长和叶宽等植株形态指标均随发育时间延长呈直线增长趋势,直线方程拟合指数R2(相关指数)均达极显著水平(P<0.01),直线方程拟合点(y)与实测点极其接近(图3)。
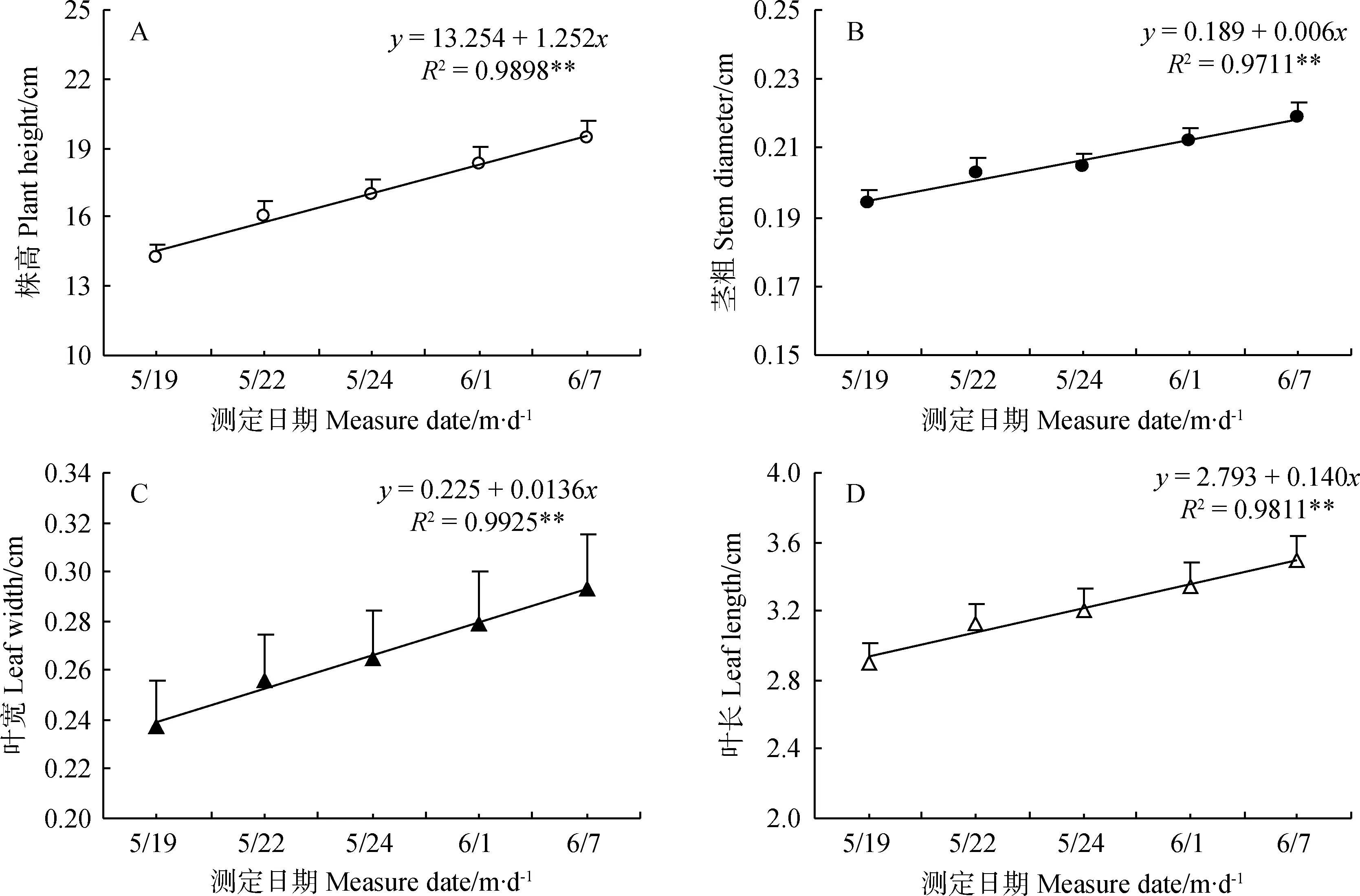
图3 漳县驯化栽培的暗紫贝母开花株发育动态及直线回归拟合(n=20)Fig.3 Dynamic of abloom plant development and lineal-regressive simulation for F. unibracteata domesticated in Zhang County(n=20)(A) 株高Plant heigh;(B) 株幅Plant diameter;(C) 叶宽Leaf width;(D) 叶长Leaf length。R2为相关指数, **表示在P≤0.01达极显著水平(下同)。R2 indicate correlation index and **indicate significant difference at the 0.01 level (the same below).
2.3 暗紫贝母花被发育和开花动态观测结果
暗紫贝母初现花蕾绿色,发育过程中逐渐加深转变为暗紫色(图4),花被弯曲下垂,吊钟形(图1B、E-H)。在花朵发育过程中,未出现花色分离现象,只是转色程度不同,花被转紫过程中极显著增长和加宽(P<0.01)(图4)。单花寿命6~8 d,最初3 d花被显著加长(t=2.467,P<0.05),花径极显著加宽(t=5.837,P<0.01),后3d加长趋势不显著,但加宽趋势仍极显著(t=2.958,P<0.01),开花10d后蒴果显现。回归分析显示,花被长随发育时间呈“S型”曲线增加,符合Logistic曲线方程y=2.782/(1+e-1.262-0.795x)(R2=0.9875,P<0.01)(图5A)。花径与发育时间呈直线关系 (R2=0.9859,P<0.01),方程拟合点与实际点极其接近(图5)。
暗紫贝母晴天开花集中在11:00-15:00,这一时间段温度高,风力大,适宜传粉。阴雨天开花时间一般滞后或者不开花。单花开花后第1 d花瓣微开(图4C),花药尚未开裂;开花后第2 d雄蕊逐渐分离,花药开裂,花粉散出(图4D);开花后第3 d(图4E)花药完全开裂,大量花粉散出;开花后第4 d花药散出花粉粒明显减少;开花后第5 d花药开始萎缩,花被逐渐脱落,开花第10 d后蒴果成形(图4F和图6A)。

图4 漳县驯化栽培的暗紫贝母花朵发育过程中花蕾开放及转色变化Fig.4 Changes of bud opening and coloring for F. unibracteata domesticated in Zhang County(A)绿色花蕾Green bud;(B) 紫色花蕾Purple bud;(C)花朵微开Tiny-opening flower;(D)朵半开Half-opening flower;(E) 花朵全开Fully opening flower;(F) 坐果Fruit setting
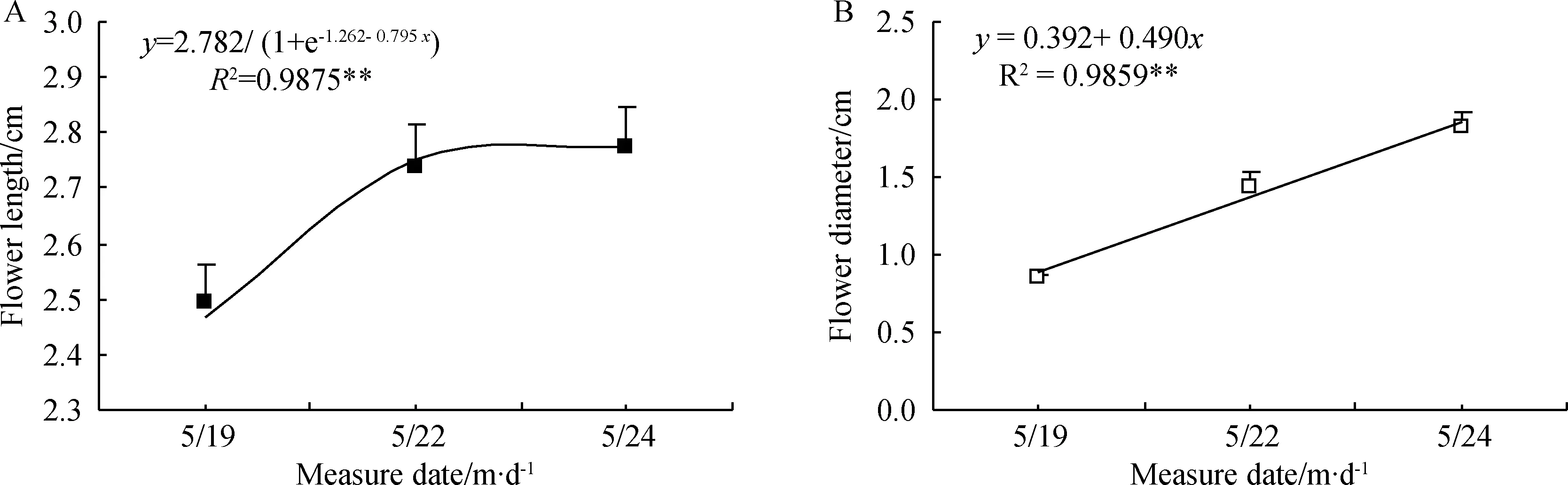
图5 漳县暗紫贝母发育过程中花朵长度(A)和花朵直径(B)变化动态及其线性回归拟合(n=20)Fig.5 Dynamics and Linear regressive simulation of flower length (A) and diameter (B) during development for F.unibracteata flowers in Zhang County (n=20)
2.4 暗紫贝母传粉结实特性
暗紫贝母坐果后,弯曲下垂的花果茎端逐渐向上生长,使蒴果与茎秆呈直立状,开花50 d后蒴果种子陆续成熟。成熟蒴果黄褐色,种子褐色,心形(图6A-H)。经选取单花植株花蕾套袋操作试验结果显示(表1),花朵套袋操作处理后暗紫贝母单花结实率变幅为0%~82.7%。去雄异株异花人工授粉套纸袋结实率最高,自然授粉次之,两者相差7.7%。去雄异株异花人工授粉后套网袋结实率明显下降,较自然授粉降低18.3%,较套纸袋降低26.0%。不去雄套纸袋较不去雄套网袋单花结实率提高6.7%,去雄后不套袋结实率较自然授粉下降5.0%。去雄套纸袋不结实,而去雄套网袋结实率较去雄套纸袋结实率提高30.0%,不去雄套纸袋较去雄套纸袋结实率提高26.7%,不去雄套网袋较去雄套网袋结实率降低10.0%(表1)。
花朵操作处理对暗紫贝母单蒴果结种子数具有极显著影响(F=84.148,P<0.01) (表1),去雄套纸袋不结蒴果和种子,其余处理组均有蒴果,蒴果均结有种子。在结蒴果的7个处理组中,单果种子数和百粒重差异性均不显著。单果平均种子数从多到少依次为去雄套纸袋异花人工授粉组>自然授粉对照组>去雄套网袋异花人工授粉组>去雄套网袋组>去雄不套袋组>不去雄套网袋组>不去雄套纸袋组。含有种子蒴果处理组中,单果种子数均在100粒以上。种子百粒鲜重单果间差异性较大,但处理间差异性不显著,从高到低依次为去雄不套袋>去雄套纸袋异花人工授粉组>自然授粉对照组>去雄套网袋异花人工授粉>不去雄套纸袋>不去雄套网袋>去雄套网袋组,种子平均百粒鲜重为(94.53±53.62) mg。
经对各处理获得的168枚蒴果统计显示,暗紫贝母中轴胎座,3心皮将子房分隔成6室,每室15~30粒种子,层叠式排列,成熟种子黄褐色(图6),单果结种子70~179粒,平均(110±22)粒,变异系数20.8%。单果种子数为80~140粒的蒴果占81.1%,种子在140粒以上的蒴果数占10.0%,8.9%单果种子数少于70粒。单果内种子重较种子数变异更大。单果百粒鲜重为23.37~528.00 mg,平均94.53±53.62 mg,百粒鲜重为60~120 mg的蒴果数占82.1%,12.5%的单果种子百粒鲜重≤60 mg,百粒鲜重>120 mg的单果数仅占5.4%,变异系数56.7%。

图6 漳县驯化栽培的暗紫贝母自然授粉蒴果和种子形态Fig.6 Shape of capsules and seeds of nature pollinated F. unibracteata plants domesticated in Zhang County(A)坐果Fruit setting;(B) 鲜果底面Fresh capsule fundus;(C) 1室种子One chamber of seeds;(D) 2室种子Two chambers of seeds;(E)蒴果及枯花被Capsule and dried perianth;(F) 果顶开裂Split capsule on top;(G)裂果底部Split capsule bottom;(H) 种子Seeds
表1 花朵操作对暗紫贝母田间结实特性的影响Table 1 Effect of flower operations on seed-set traits of F. unibracteata Hsiao et K. C. Hsia plants
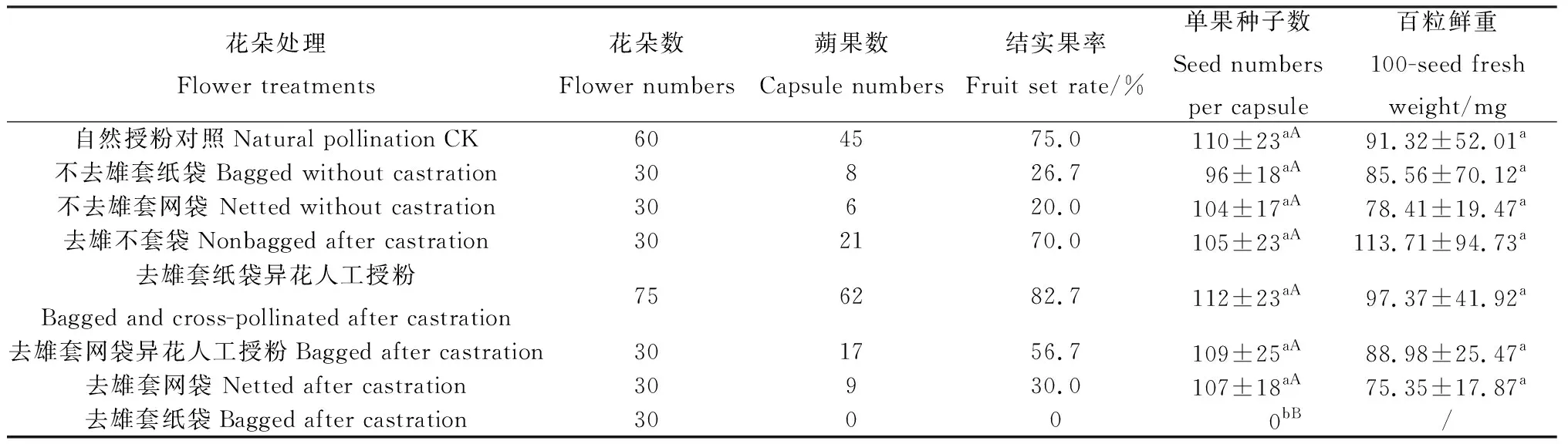
花朵处理Flower treatments花朵数Flower numbers蒴果数Capsule numbers结实果率Fruit set rate/%单果种子数Seed numbers per capsule百粒鲜重100-seed fresh weight/mg自然授粉对照Natural pollination CK604575.0110±23aA91.32±52.01a不去雄套纸袋 Bagged without castration30826.796±18aA85.56±70.12a不去雄套网袋 Netted without castration30620.0104±17aA78.41±19.47a去雄不套袋Nonbagged after castration302170.0105±23aA113.71±94.73a去雄套纸袋异花人工授粉Bagged and cross-pollinated after castration756282.7112±23aA97.37±41.92a去雄套网袋异花人工授粉Bagged after castration301756.7109±25aA88.98±25.47a去雄套网袋 Netted after castration30930.0107±18aA75.35±17.87a去雄套纸袋Bagged after castration30000bB/
注:表中数据为平均数±SD,同列数字后不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)
Note:The data in the
Table indicate means±SD, different small letters after data in the same column indicate significant difference at the 0.05 level, and the capital letters mean significant difference at the 0.01 level.
3 讨论
3.1 暗紫贝母田间植株形态多样化,物候特征明显
种质是指亲代传递给子代的遗传物质,对种质资源或遗传资源的有效保护可以通过对生态环境的保护、建立自然保护区、植物园和生物种质库来实现[17]。人工驯化不仅可以有效保护珍稀濒危药用植物物种,甚至能使其不断繁衍而解除濒危状态[17]。徐波等[4]调查提出,野生状态下暗紫贝母种子萌发后第1年地上部分仅长出1片叶,俗称“一根针”;第2年长出一片叶子,俗称“一匹叶”;第3年才开始抽出茎,长出多片叶,俗称“树儿子”;第4年地上茎长出花,进入有性繁殖阶段,俗称“灯笼花”。李西文和陈士林[19]观察分析,川贝母存在明显的繁育退化现象,当出现极端气候时,从灯笼花发育阶段可直接退回到一匹叶阶段。陈士林等[20]通过数值分析显示,暗紫贝母植株随生态分化株形差异较大,花期“灯笼花”植株高约17~35 cm,“一匹叶”植株(营养生长期)高约7~15 cm。本研究发现,暗紫贝母自然更新速度很慢,2013年经采用大鳞茎(4~5年生)播种,次年返青后田间以“一匹叶”居群为主,开花株稀少。随栽培年限延长,物候有提早增强趋势,“树儿子”和“灯笼花”株率提高,但“一匹叶”仍占据优势,地下鳞茎数也因株形不同出现差异,单株开花数年分配比例逐年提高,“灯笼花”株高15~27 cm,茎粗0.192~0.268 cm,而野生条件下因人畜扰动和见花采挖,多年生大鳞茎稀少,很难见到多花单株,单花株较为普遍,开花株茎表观健壮,实易折断,这可能是其保护地下繁殖体的应对策略,这与Shoichi对日本贝母的观察描述一致[13]。
暗紫贝母植株形态差异和开花株比率的年分配是环境和遗传共同作用的结果,在其生长发育过程中,植株个体间出现的这种差异是其在极端环境条件下得以生存繁衍的一种本能反应,也是稀有物种得以繁衍的暗示[21]。田间株形多样化也充分显示出暗紫贝母无性和有性繁殖的叠加可再生性能。尽管暗紫贝母在长期应对多变的高山极端环境过程中形成了独特的无性繁殖能力,种子还有油脂易被蚂蚁扩散[13],但自然有性繁殖几率很低,因为2015年返青后有一定比例的结果株种果开裂落地,但2016年返青后田间少见“一根针”苗,说明成熟蒴果种子自然出苗繁殖几率很低。野生环境下,动物踩踏啃食和鸟啄造成种子的流失,加上种子一旦落地后不能持续保湿则难以完成其后熟,而是进入深休眠,这也印证了常彦莉[14]的研究结果,即川贝母种子具有明显的后熟和休眠特性。
本研究还发现,暗紫贝母抽薹开花期营养生长与生殖生长并进,即植株进入生殖生长阶段后,营养生长还继续,7月中下旬高温到来之前地上部分枯萎倒苗,说明暗紫贝母能充分利用极其有限的适温资源完成其地上年生物量积累分配,温度急剧升高不利于暗紫贝母地上部分的生长发育,因为气候升温越快,暗紫贝母倒苗越早,杂草丛中植株倒苗期延后,但倒苗可能更有利于地下鳞茎生物量的积累分配和有效成分的转化积累,茎杆的中空结构为地上部分营养成分向根茎部转运提供了畅通无阻的通道,倒苗后地上部分逐渐分解也印证了暗紫贝母植株营养物质各器官转运分配的能力。
3.2 暗紫贝母属异花授粉类型,部分自交亲和,不存在无融合生殖现象
植物花开放后由花粉囊散出的花粉借助一定的媒介力量被传送到同一花或另一花的柱头上的传递过程称为传粉作用,传送花粉的外力有风、动物、水等,传粉在植物繁殖进化和性状的遗传变异等方面具有重要的意义[18]。传粉受精过程与自然界中的传粉者和传粉行为相结合构成了整个植物有性生殖纽带。有性繁殖药用植物根据花器构造、开花习性及授粉方式的不同可分为自花授粉、异花授粉和常异花授粉植物3种类型。育种中,一般根据异交率判断繁育类型,异交率<5%归为自花授粉,异交率>50%以上为异花授粉,而介于5%~50%为常异花授粉[16]。张晓军等[22]研究表明,平贝母(F.ussuriensis)是以自花传粉为主,虫媒异花传粉为辅的兼性自交植物。事实上,在自然界中很难见到绝对的异交或者绝对自交的植物类群,大部分植物是二者兼有的混交模式[18],因为过度的自交和异交都会导致植物对环境的应变能力减弱,适应性下降。
本研究在漳县3100 m海拔2013年采用大鳞茎播种的暗紫贝母2015年花蕾期田间株型多样化特征明显,5叶及以上植株在5月中下旬现蕾开花,花蕾去雄套纸袋处理不结实,验证本研究选蕾恰当,此期花蕾雄蕊未熟,去雄有效,也说明不存在无融合生殖现象,这与胡春等[23]对青藏高原优势毒杂草钝裂银莲花(Anemoneobtusiloba)研究的结论相反,反映出毒草自繁能力更强。自然授粉结果率为75.0%,去雄套纸袋异花人工授粉结实率达82.7%,说明属异花授粉类型,但存在一定的花粉限制,这与任明迅[24]对半边莲的研究结果相似,不去雄套袋自然结实率20.0%~26.7%,说明暗紫贝母具有一定自交亲和性(低于30%),花成熟前后雌雄蕊高低错位和花的下垂弯曲结构可能也是花粉自传几率较低的根源所在。
自花传粉植物西卡柱花草(Stylosanthesscabra)在清晨花朵开放前完成授粉,花粉粒萌发伸长[25],暗紫贝母自交也可能是极大限度繁衍后代的机能体现。去雄不套袋异花授粉结实率(70.0%)较去雄套网袋异花授粉结实率(56.7%)提高13.3%,说明主要传粉媒介为虫媒,但也不排除风媒,进一步验证了陈垣等[7]对其繁育系统研究推测的结论。可能去雄削弱了花器对柱头的保温保湿作用,纸袋从某种程度上可减弱去雄的伤害效应,这也从不去雄套网袋(20.0%)较不去雄套纸袋结实率(26.7%)降低这一结果加以解释,因为前者包含自花和风媒传粉两层含义,而后者仅包含自花传粉效率。不去雄套网袋较去雄不套袋结实率降低也说明传粉媒介除风媒外,主要为虫媒,田间也发现有少量访花蜂类、蝇类、蜘蛛和蚂蚁等,花中有蜜腺且花粉量大而散,去雄不套袋结实率高于去雄套网袋更支持虫媒传粉的证据,因为套网袋隔离了小昆虫进入花的几率,但并没有排除风传花粉的可能。
本研究中,去雄操作仅去除花药,并不影响昆虫访花的决定,因为光谱显示蜜蜂并不能辨别花瓣和花粉的颜色,将花瓣鲜黄色的百合科植物去除雄蕊后访花昆虫大大减少,但去除花药保留花丝种果率最高[12],显花植物特有地利用花行使信号传导功能,吸引传粉昆虫而排斥食花害虫访花的决定,黄色能吸收紫外光,并能扮演花粉和花粉囊色[13]。暗紫贝母花被内镶嵌黄色格纹,雄蕊和雌蕊柱头均呈黄绿色,子房呈紫绿相间圆筒形,具有植株小、花大、群体花期长和花粉多而散的特点,这也印证了其虫媒和风媒交叉传粉的可能性。
4 结论
人为扰动和气候变暖趋势加剧了野生暗紫贝母栖息地的退缩变迁,野生资源趋于枯竭。野生抚育和驯化栽培是促进资源可持续化发展的重要途径,利用种子繁殖可提高繁殖系数,有效保护野生资源。暗紫贝母田间株形多样,单株叶片数和开花数比率的年分配趋势明显,自然异交率约70%,自交率低于30%,属典型异花授粉植物,不存在无融合生殖现象,群体开花期的延长可为稀有访花者和风力提供传粉几率,高寒区夜间风力更大,气温骤降,低温更有利于花粉维持足够时间的活力,提高传粉效率,这些特有的物候特征、田间株型多样化和传粉结实特性是其适应高寒恶劣多变环境的结果,高的异交率和自交花粉限制是其克服繁育退化的一种应激能力。传粉媒介以虫媒为主,风媒为辅,访花有效传粉昆虫有蜜蜂、蝇类、蜘蛛、蝶类和蚂蚁等,对有效传粉昆虫有待进一步鉴定研究。由于开花株茎秆脆弱易折断,开花结果期尽量减少田间人为扰动。种子繁育需采取隔离措施并提倡养蜂,以提高种子产量,确保种子纯度。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
疯狂英语·新悦读(2022年12期)2022-03-08
艺术品鉴(2021年12期)2021-05-25
浙江林业科技(2020年6期)2020-12-25
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
基层中医药(2018年9期)2018-11-09
中外医疗(2018年31期)2018-01-17
中成药(2017年5期)2017-06-13
中国野生植物资源(2014年4期)2014-03-29
种子科技(2011年10期)2011-09-08
