
增施氮素对苗期渍水胁迫冬油菜生理特性及产量的调控效应①
2017-07-21 05:05陈红琳陈尚洪郑盛华沈学善刘定辉
土壤 2017年3期
陈红琳,陈尚洪,郑盛华,沈学善,刘定辉
增施氮素对苗期渍水胁迫冬油菜生理特性及产量的调控效应①
陈红琳,陈尚洪,郑盛华,沈学善,刘定辉*
(四川省农业科学院土壤肥料研究所,成都 610066)
研究增施氮素对苗期渍水胁迫后冬油菜生理特性及产量的调控效应,可为冬油菜主产区渍水胁迫后植株生长恢复调控措施的选择提供重要参考。本研究采用盆栽和大田试验,在冬油菜5 ~ 6叶期设置渍水胁迫D0(0 d)、D6(6 d)和D9(9 d)三个处理,并在渍害胁迫解除后设置4个氮素增施水平N0(0 kg/hm2)、N2(30 kg/hm2)、N4(60 kg/hm2)、N6(90 kg/hm2),研究了增施氮素对苗期渍水胁迫后冬油菜的生理响应特性及产量的调控效应。结果表明:与单纯渍水不增施氮素相比,增施氮素均能有效改善冬油菜各项生理指标而提高产量,盆栽和大田试验各指标变化特征基本一致。增施氮素20 d后,随渍水胁迫天数的减少与氮素施用水平的增加,盆栽及大田植株叶片叶绿素含量、叶片和根系超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性以及产量增加,叶片和根系丙二醛(MDA)含量降低,除植株根系CAT活性以外,交互作用对其余各项生理指标及产量的影响均达到显著水平(<0.05)。与单纯渍水不增施氮素相比,盆栽及大田N4D6处理冬油菜叶绿素总含量、叶片和根系抗氧化酶活性以及产量均显著增加13.28% ~ 26.98%(<0.05),叶片和根系丙二醛含量显著降低20.24% ~ 23.67%(<0.05),除POD活性外,其余各指标均已恢复至自然对照水平;盆栽及大田N6D9处理叶绿素总含量、叶片和根系抗氧化酶活性以及产量均显著增加(<0.05),叶片和根系MDA含量显著降低(<0.05),但所有指标仍均未恢复至自然对照水平。因此,在冬油菜主产区苗期渍水胁迫6 d后增施纯氮60 kg/hm2以及渍水胁迫9 d后增施纯氮90 kg/hm2均能显著增强油菜渍水胁迫后的生理代谢活动,而提高作物产量。
氮素;苗期;渍水胁迫;油菜;抗氧化酶
成都平原位于四川盆地西部,是四川冬油菜主产区之一。由于免耕直播省工省力和不易倒伏[1],因而冬油菜在该区种植面积逐年扩大。然而,冬油菜苗期常因遭遇连绵秋雨天气,从而导致渍害影响后期植株抗性、角果发育和籽粒充实[2]。已有研究表明,冬油菜苗期渍水导致植株生理活动受到严重影响,叶绿素含量、光合速率、抗氧化酶(超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT))活性和根系活力显著降低,从而导致籽粒产量下降,渍水9 d以上冬油菜产量相对受害率达20% 以上[3-5]。因此,探索苗期渍水胁迫后冬油菜的生长恢复措施对免耕直播冬油菜实现高产栽培具有重要意义。
目前作物生产中减轻渍害的技术措施主要包括:选育选用耐湿性强的作物品种、开沟排水降低地下水位以及在渍害发生之后采取水肥调控或化学调控等补救措施[6-8]。其中,耐湿品种的筛选鉴定以及开沟排湿耗时费力,而化学调控生产资料成本较高且可能影响作物品质[9]。与之相反,氮素调控因能在一定程度上缓解涝渍对植株生长的伤害作用且施用方便,常被作为遭受渍害作物恢复生长的重要措施[10]。研究表明,水分逆境下增施氮肥可提高小麦籽粒可溶性淀粉合成酶、结合态淀粉合成酶等酶活性以及蛋白质含量[11];施氮可提高水分胁迫下玉米和棉花光合性能,使玉米和棉花增产10% 以上[12-13]。氮素作为对油菜产量和氮肥吸收利用效率影响最大的营养元素[14],在冬油菜返青期复水后,适量增施氮素对植株地上部干物质量、叶绿素含量、光合速率、产量及产量构成有一定程度的补偿效果[15],但氮素对苗期渍水胁迫后冬油菜产量的补偿调控效应以及生理特性影响机制研究还尚少。
长江上游尤其是四川主产区冬油菜渍害的研究亦主要在种子耐湿性评价及幼苗对渍害的形态、生理指标响应方面[16-18],有关成都平原区冬油菜苗期渍害调控方面的研究还尚未见报道。本研究以四川省冬油菜生产中主推甘蓝型品种“川油36”为供试材料,结合盆栽与大田试验,在冬油菜苗期采用水淹处理模拟渍水胁迫,渍水胁迫解除后增施氮素,旨在探索氮素对苗期渍水胁迫后冬油菜生理特性和产量的影响,并阐明其机理,为渍水胁迫下冬油菜抗灾减灾提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为四川省冬油菜生产主推品种“川油36”,由四川省农业科学院作物研究所提供;肥料品种分别为尿素(含N≥460 g/kg),过磷酸钙(含P2O5≥120 g/kg),氯化钾(含K2O≥600 g/kg),速滋硼(含B≥200 g/kg)。
1.2 试验设计
本研究采用盆栽试验和大田试验相结合的方法,待冬油菜5 ~ 6叶期设置D0(渍水0 d,土壤含水量保持在田间持水量的70% ~ 80%)、D6(6 d)、D9(9 d)3个渍水胁迫时间,渍水处理时始终保持水面高出土壤表面1 cm;在渍水胁迫解除恢复正常水分管理3 d后,设置增施纯氮N0(0 kg/hm2)、N2(30 kg/hm2)、N4(60 kg/hm2)、N6(90 kg/hm2)4个水平。盆栽试验和大田试验分别共12个处理,以N0D0处理为正常生长的自然对照(CK)。
盆栽试验:试验于2015年9月—2016年5月在四川省农业科学院土壤肥料研究所网室进行。供试土壤为水稻土,采自四川省广汉市西高镇李堰村大田试验田块耕层(0 ~ 20 cm),pH为6.50,有机质31.80 g/kg,全氮17.16 g/kg,速效氮199.50 mg/kg,有效磷22.50 mg/kg,速效钾66.50 mg/kg。土壤风干后,过2 mm筛,装盆(每盆7.00 kg,在盆身沿盆周打孔3圈,分别距离盆底距离5、15、25 cm,孔径约1 cm,并贴上过滤网),每处理3次重复。9月下旬播种,播种前施入纯N 0.57 g/盆,P2O50.28 g/盆,K2O 0.36 g/盆,B 0.01 g/盆。4叶期定苗,每盆均匀留苗2株。
大田试验:同期在广汉市西高镇李堰村大田进行。前茬作物为水稻,土壤肥力均匀。每处理为1小区,小区面积4 m × 5 m,3次重复,随机区组排列。为避免水分侧渗,小区间间隔1.5 m,各小区筑埂30 cm厚、30 cm高,并用塑料薄膜对0 ~ 100 cm土层进行水分隔离。9月下旬播种,播种前施用纯N 180.00 kg/hm2,P2O590.00 kg/hm2,K2O 117.00 kg/hm2,B 3.00 kg/hm2。植株行距、株距均为25 cm,4叶期定苗,种植密度为15.00万株/hm2。
1.3 测定指标与方法
1.3.1 生理特性指标 叶片叶绿素含量、丙二醛(MDA)含量和抗氧化酶活性测定选用新叶下第3个叶片,根系MDA含量和抗氧化酶活性测定主要选用靠近根尖的须根部分[19]。各项指标分别于增施氮素当天(第0 天)、第5 天、第10 天、第15天、第20天进行取样测定。盆栽试验每处理每重复取样2盆,大田试验每处理每重复取样4株。
叶绿素a、叶绿素b含量采用分光光度法测定,MDA含量采用硫代巴比妥酸(TBA)反应法测定[20]。称取新鲜叶片0.500 0 g,加pH 7.8的磷酸缓冲液6 ml于预冷的研钵中研磨提取,在4℃下,4 000 r/min离心20 min,即得提取液,用超低温冰箱保存用于SOD、POD、CAT活性测定。SOD活性采用氮蓝四唑( NBT) 还原法,POD活性采用愈创木酚法,CAT活性采用过氧化氢法测定[20]。
1.3.2 产量 成熟期盆栽试验调查每处理植株总株数及总产量,计算单株产量;大田试验各小区适时收获,采用水分测定仪测定油菜籽粒水分含量,调查小区实收产量。
1.4 数据分析
采用SPSS 10.0统计软件进行统计分析及单因素方差(ANOVA)检验,采用新复极差法(Duncan)作处理间平均值的多重比较。
2 结果与分析
2.1 增施氮素对苗期渍水胁迫冬油菜叶绿素总含量的影响
在盆栽及大田条件下,与渍水胁迫D0处理相比,渍水胁迫D6、D9处理的冬油菜叶绿素a+b总含量显著降低21.19% 以上(表1)。与单纯渍水不增施氮素处理(N0D6、N0D9)比较,增施氮素均不同程度提高了盆栽及大田不同渍水处理的叶绿素总含量。增施氮素第20天时,盆栽及大田渍水胁迫D6、D9处理植株叶绿素总含量分别以N4D6、N6D9处理最高,较对应未增施氮素处理显著增加17.75% 以上,且N4D6处理均已恢复至自然对照(CK)水平,与CK差异不显著,而N6D9处理仍较CK显著降低23.73% 以上。方差分析结果表明,盆栽及大田渍水胁迫天数、氮素增施水平以及二者交互作用对叶绿素总含量均有显著影响(<0.05),随渍水胁迫天数的减少与氮素施用水平的增加,植株叶绿素总含量增加。
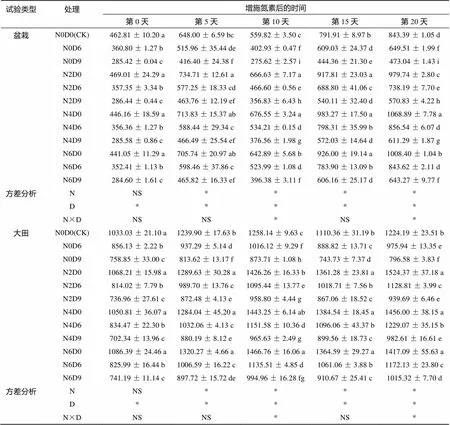
表1 增施氮素对渍水胁迫冬油菜叶绿素总含量的影响(mg/kg)
注:N0D0、N2D0、N4D0、N6D0分别表示渍水胁迫0 d增施氮素0 kg/hm2(自然对照,CK)、30、60、90 kg/hm2;N0D6、N2D6、N4D6、N6D6分别表示渍水胁迫6 d增施氮素0、30、60、90 kg/hm2;N0D9、N2D9、N4D9、N6D9分别表示渍水胁迫9 d增施氮素0、30、60、90 kg/hm2。数据表示为均值±标准误差,同列小写字母不同表示处理间差异达到<0.05显著水平。* 表示在<0.05水平差异显著,NS为不显著。下同。
2.2 增施氮素对苗期渍水胁迫冬油菜抗氧化酶活性的影响
在盆栽及大田条件下,与渍水胁迫D0处理相比,渍水胁迫D6、D9处理均降低了冬油菜幼苗叶片和根系SOD活性,显著降低了叶片POD活性以及叶片、根系CAT活性,提高了幼苗根系POD活性(图1、图2、图3)。
增施氮素对冬油菜抗氧化酶活性的影响特征在盆栽与大田条件下表现基本一致。与单纯渍水胁迫不增施氮素相比,增施氮素均不同程度提高了不同渍水胁迫幼苗叶片和根系SOD、POD和CAT活性(图1、图2、图3)。增施氮素第20天时,植株叶片和根系SOD、POD及CAT活性在渍水胁迫D6处理中均以N4D6处理最高,较N0D6处理显著增加13.28% ~ 26.98%,且叶片和根系SOD、CAT活性均已恢复至CK水平,但与CK差异不显著,叶片和根系POD活性较CK略有降低;在渍水胁迫D9处理中均以N6D9处理最高,但仍显著低于CK水平。方差分析结果表明,渍水胁迫天数、氮素增施水平对植株叶片和根系的各抗氧化酶活性均有显著影响(<0.05),二者交互作用对植株根系CAT活性无显著影响。
2.3 增施氮素对苗期渍水胁迫冬油菜丙二醛含量的影响
在盆栽及大田条件下,与渍水胁迫D0处理相比,渍水胁迫D6、D9处理均显著增加了冬油菜幼苗叶片和根系MDA含量(图4)。而与单纯渍水胁迫不增施氮素相比,增施氮素均不同程度降低了不同渍水胁迫幼苗叶片和根系MDA含量。增施氮素第20天时,N4D6处理叶片和根系MDA含量且均已恢复至CK水平,但与CK差异不显著,较N0D6处理显著降低20.24% ~ 23.67%;N6D9处理叶片和根系MDA含量仍较CK显著增加22.09% ~ 27.50%,但较N0D9处理显著降低20.92% ~ 25.69%。方差分析结果表明,盆栽及大田渍水胁迫时间、氮素增施水平及二者交互作用对植株叶片和根系的MDA含量均有显著影响(<0.05),随渍水胁迫天数的减少和氮素施用水平的增加,植株叶片和根系MDA含量降低。
2.4 增施氮素对苗期渍水胁迫冬油菜产量的影响
与渍水胁迫D0处理相比,成熟期盆栽及大田渍水胁迫D6处理冬油菜产量显著降低12.90% 以上,渍水胁迫D9处理冬油菜产量显著降低22.66% 以上;与单纯渍水不增施氮素相比,增施氮素均不同程度提高了冬油菜产量(表2)。成熟期盆栽及大田N4D6处理产量较N0D6处理显著增产14.44% 以上,且均恢复至CK水平;N6D9处理较N0D9处理显著增产21.26% 以上,但仍较CK显著减产8.57% 和6.21%。方差分析结果表明,盆栽及大田渍水胁迫天数、氮素增施水平以及二者交互作用对产量均有显著影响(<0.05),随渍水胁迫天数的减少和氮素施用水平的增加,冬油菜产量增加。
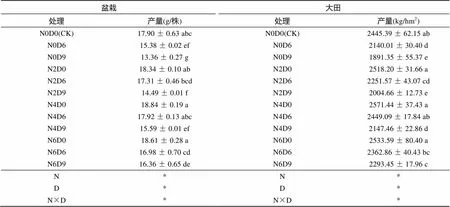
表2 增施氮素对苗期渍水胁迫冬油菜产量的影响
3 讨论
3.1 叶绿素含量
叶绿素是光合作用的主要色素,常作为衡量叶片衰老程度的指标,叶绿素含量的高低可反映出光合能力的强弱[15]。渍水造成植物根系缺氧,生理上引起活性氧大量积累,导致叶绿体结构遭到破坏,从而使油菜叶片中叶绿素a、叶绿素b等光合色素含量显著降低[4,21]。氮素是叶绿体的主要成分,氮素对作物叶绿素含量的影响直接制约着光合作用的强弱[22-24],施氮能促进植物叶片叶绿素的合成。本试验条件下,盆栽及大田渍水胁迫6 d和9 d显著降低了冬油菜叶绿素a+b总含量。与单纯渍水不增施氮素相比,渍水胁迫后增施氮素均提高了冬油菜叶绿素总含量,增施氮素60 kg/hm2能使渍水胁迫6 d幼苗叶绿素总含量恢复至自然对照水平,但增施氮素90 kg/hm2仍未能使渍水胁迫9 d幼苗叶绿素总含量恢复至自然对照水平。综上所述,在冬油菜主产区,油菜遭渍水胁迫后增施氮素能在一定程度缓解光合色素因渍水胁迫引起的降解,从而提高植株光合作用能力,但长时间渍水胁迫可能导致幼苗叶绿体结构遭到严重破坏,其光合色素的合成不易恢复至未渍水胁迫状态。
3.2 抗氧化系统
植物在逆境胁迫下,植株体内活性氧过度积累,从而导致膜脂过氧化程度加剧,造成膜系统伤害,而抗氧化酶系统及抗氧化物质是植物体内清除活性氧的主要途径[25]。油菜遭渍水胁迫后MDA含量以及各抗氧化酶活性的响应特征因渍水天数的差异而不同[3-4]。本研究表明,油菜遭连续渍水胁迫6 d以上,冬油菜叶片和根系SOD、CAT活性以及叶片POD活性均降低,根系POD活性、叶片和根系MDA含量升高。氮素是植物体内蛋白质、核酸、激素等重要物质的组成成分,对植物的生命活动具有重要的意义[26]。本试验条件下,与单纯渍水不增施氮素相比,增施氮素均能有效改善冬油菜各项生理指标,增施氮素第20天时,渍水胁迫天数、氮素增施水平以及二者交互作用对除根系CAT活性以外的其余各项生理指标均有显著影响(<0.05)。说明渍水胁迫后增施氮素可能诱导了相关抗氧化酶基因的表达,改善了植株的各项生理指标,能显著提高SOD、POD、CAT活性,从而能及时清除因渍水胁迫产生的活性氧,减弱细胞膜脂过氧化程度,促进了植株生理代谢活动的恢复。增施氮素60 kg/hm2可使渍水胁迫6 d的植株除POD活性外的其余各项指标均恢复至最佳并略高于自然对照水平,增施氮素90 kg/hm2可使渍水胁迫9 d的植株各项指标性状均较未增施氮素植株显著改善,但仍未能恢复至自然对照水平。可能由于渍水胁迫9 d以上对植株的氧化胁迫危害已严重超出自身代谢平衡系统,增施氮素后仍不能及时完全清除因渍水胁迫9 d产生的活性氧或清除速率缓慢,细胞膜脂过氧化程度减弱缓慢,或与长时间渍水胁迫后氮素增施量不足有关。因此,关于长时间渍水胁迫(渍水胁迫天数≥9 d)的氮素最佳增施水平以及增施氮素后冬油菜的蛋白或基因表达差异还有待进一步研究,这对冬油菜渍水胁迫后植株生长恢复以及耐湿性品种的筛选有重要作用。
3.3 产量
氮肥是影响油菜产量的重要因素[27]。谷晓博等[15]研究表明,在油菜返青期水分亏缺时,增施氮量90 kg/hm2时补偿效果最佳;但邹小云等[28]就花期渍水逆境下氮素对油菜产量及氮肥利用效率的影响结果表明,花期渍水逆境下增施氮肥对油菜籽粒产量的形成贡献不大。而本研究试验结果表明,渍水胁迫后,盆栽及大田增施氮素均能不同程度提高冬油菜产量,渍水胁迫天数、氮素增施水平以及二者交互作用对产量均有显著影响(<0.05)。说明渍水胁迫后增施氮素改善了冬油菜苗期叶片的光合和植株的生理指标,促进了植株生理代谢活动的恢复,从而苗期维持了植株一定的生长量和干物质积累,为冬油菜渍水胁迫后产量的形成及产量补偿奠定了基础。渍水胁迫6 d时,增施氮素60 kg/hm2处理产量最高,已恢复至自然对照水平,较不增施氮素显著增产14.44% 以上;渍水胁迫9 d时,增施氮素90 kg/hm2处理产量最高,较未增施氮素显著增产21.26% 以上。综上所述,冬油菜遭渍水胁迫后可通过增施氮素来恢复植株的生理代谢功能和挽回产量损失,为使植株恢复至未受渍水胁迫产量水平时还应注重氮素的合理增施量。
4 结论
在冬油菜主产区,冬油菜苗期渍水胁迫6 d、9 d分别导致产量较未渍水胁迫显著降低12.90% 以上和22.66% 以上,但渍水胁迫6 d后增施纯氮60 kg/hm2可挽回产量损失14.44%,渍水胁迫9 d后增施纯氮90 kg/hm2可挽回产量损失21.26%。
[1] 苏伟, 鲁剑巍, 周广生, 等. 免耕及直播密度对油菜生长、养分吸收和产量的影响[J]. 中国农业科学, 2011, 44(7): 1519–1526
[2] 王寅, 汪洋, 鲁剑巍, 等.直播和移栽冬油菜生长和产量形成对氮磷钾肥的响应差异[J]. 植物营养与肥料学报, 2016, 22(1): 132–142
[3] 王琼, 张春雷, 李光明, 等. 渍水胁迫对油菜根系形态与生理活性的影响[J]. 中国油料作物学报, 2012, 34(2): 157–162
[4] 李玲, 张春雷, 张树杰, 等. 渍水对冬油菜苗期生长及生理的影响[J]. 中国油料作物学报, 2011, 33(3): 247–252
[5] 宋丰萍, 胡立勇, 周广生, 等.渍水时间对油菜生长及产量的影响[J]. 作物学报, 2010, 36(1): 170–176
[6] 陶霞, 李慧琳, 万林, 等.叶面喷施吲哚乙酸对油菜蕾薹期渍水的缓解效应[J]. 中国油料作物学报, 2015, 37(1): 55–61
[7] 李玲, 李俊, 张春雷, 等.外源ABA和BR在提高油菜幼苗耐渍性中的作用[J]. 中国油料作物学报, 2012, 34(5): 489–495
[8] Leul M, Zhou W J. Alleviation of waterlogging damage in winter rape by application of uniconazole: Effects on morphological characteristics, hormones and photosynthesis[J]. Field Crops Research. 1998, 59(2): 121–127
[9] 秦东玲, 李钊, 尉菊萍, 等.作物抗冷性及其化学控制机理研究进展[J]. 作物杂志, 2016(4): 26–35
[10] 薛晓萍, 周治国, 张丽娟, 等.棉花花后临界氮浓度稀释模型的建立及在施氮量调控中的应用[J]. 生态学报, 2006, 26(6): 1781–1791
[11] 范雪梅, 姜东, 戴廷波, 等.花后干旱和渍水下氮素供应对小麦籽粒蛋白质和淀粉积聚关键调控酶活性的影响[J]. 中国农业科学, 2005, 38(6): 1132–1141
[12] 陈家宙, 林丽蓉, 吕国安, 等.红壤施氮对玉米水分胁迫指数的影响[J]. 植物营养与肥料学报, 2010, 16(5): 1114–1119
[13] 郭文琦, 赵新华, 陈兵林, 等. 氮素对花铃期短期渍水棉花根系生长的影响[J]. 作物学报, 2009, 35(6): 1078– 1085
[14] 邹小云, 刘宝林, 宋来强, 等. 花期渍水逆境下氮素对油菜产量及氮肥利用效率的影响[J]. 应用生态学报, 2016, 27(4): 1169–1176
[15] 谷晓博, 李援农, 杜娅丹, 等.不同施氮水平对返青期水分胁迫下冬油菜补偿效应的影响[J]. 中国生态农业学报, 2016, 24 (5): 572–581
[16] 张雪花, 李浩杰, 张锦芳, 等. 甘蓝型油菜耐湿性初步评价和主成分分析[J]. 中国油料作物报, 2011, 33(2): 98– 103
[17] 李浩杰, 张雪花, 蒲晓斌, 等.甘蓝型油菜对田间模拟湿害胁迫的生理响应[J]. 西南农业学报, 2013, 26(1): 84– 88
[18] 李浩杰, 柴靓, 蒲晓斌, 等.室内水淹和田间模拟湿害对甘蓝型油菜耐湿性鉴定[J]. 西南农业学报, 2016, 29(6): 1250–1256
[19] 张树杰, 廖星, 胡小加, 等. 渍水对油菜苗期生长及生理特性的影响[J]. 生态学报, 2013, 33(23): 7382–7389
[20] 王学奎, 黄见良. 植物生理生化实验原理与技术[M]. 北京: 高等教育出版社, 2015
[21] 张晓红, 冯梁杰, 杨特武, 等.冬季低温胁迫对油菜抗寒生理特性的影响[J]. 植物生理学报, 2015, 51(5):737–746
[22] 郭天财, 冯伟, 赵会杰, 等.两种穗型冬小麦品种旗叶光合特性及氮素调控效应[J]. 作物学报, 2004, 30(2): 115–121
[23] 许东亚, 焦哲恒, 孙榅淑, 等. 烤烟成熟期氮素灌淋调亏对烟叶生长发育及质量的影响[J]. 土壤, 2015, 47(4): 658–663
[24] 宝俐, 董金龙, 李汛, 等. CO2浓度升高和氮素供应对黄瓜叶片光合色素的影响[J]. 土壤, 2016, 48(4): 653–660
[25] 刘广明, 李金彪, 王秀萍, 等. 外源水杨酸对黑麦草幼苗盐胁迫的缓解效应研究[J]. 土壤学报, 2016, 53(4): 995–1002
[26] 杨晓清, 周再知, 梁坤南, 等. 氮素对模拟胁迫下土沉香幼苗抗旱生理的影响[J]. 热带作物学报, 2013, 34(6): 1121–1127
[27] 王寅, 鲁剑巍, 李小坤, 等. 江浙油菜主产区冬油菜的区域适宜施氮量研究[J]. 土壤学报, 2013, 50(6): 1117– 1128
[28] 邹小云, 刘宝林, 宋来强, 等.花期渍水逆境下氮素对油菜产量及氮肥利用效率的影响[J]. 应用生态学报, 2016, 27(4): 1169–1176
Regulation Effects of Adding Nitrogen on Physiological Properties and Yield of Rapeseed After Waterlogging During Seedling
CHEN Honglin, CHEN Shanghong, ZHENG Shenghua, SHEN Xueshan, LIU Dinghui*
(Soil and Fertilizer Institute, Sichuan Academy of Agricultural Sciences, Chengdu 610066, China)
The regulation of adding nitrogen on crop physiological properties and yield after waterlogging stress can provide valuable information for restoring crop growth. In this study, the method of combining field and pot experiments were adopted, including four adding nitrogen rates of 0, 30, 60 and 90 kg/hm2and three waterlogging time treatments of 0, 6 and 9 days. The results showed that compared with the treatment of only waterlogging stress but no adding nitrogen, adding nitrogen improved the physiological indexes and enhanced the yield, all the indexes had similar changing characters in both pot and field experiments. After 20 days from adding nitrogen, the chlorophyll contents in leaves, the activities of SOD, CAT and POD in leaves and roots, as well as the yield increased and the MDA contents in leaves and roots decreased while the waterlogging time decreased and the nitrogen rates increased, waterlogging time and nitrogen rates had significant interaction effects on the other physiological indexes and yield except the CAT activities in roots (<0.05). Compared with the treatment of only waterlogging stress but no adding nitrogen, the chlorophyll contents in leaves, the activities of SOD, POD and CAT in leaves and roots, as well as the yield significantly increased from 13.28% to 26.98% (<0.05), the MDA contents in leaves and roots significantly decreased 20.24% to 23.67% under the treatment with waterlogging lasted 6 days and applying N 60 kg/hm2in both pot and field experiments (<0.05), other indexes already restored to the level of the treatment of no waterlogging stress and adding nitrogen (CK) except the POD activities; The chlorophyll contents in leaves, the activities of SOD, POD and CAT in leaves and roots, as well as the yield significantly increased (<0.05), the MDA contents in leaves and roots significantly decreased under the treatment with waterlogging lasted 9 days and applying N90 kg/hm2in both pot and field experiments (<0.05), but all the indexes could not restore to the level of CK. In the main planting region of winter rape, adding 60 kg/hm2after 6 d waterlogging stress or 90 kg/hm2nitrogen after 9 d waterlogging stress can restore and promote the plant growth and reduce the yield loss.
Nitrogen; Seedling; Waterlogging stress; Rapeseed; Antioxidative enzymes
10.13758/j.cnki.tr.2017.03.014
S565.4;S14
A
四川省财政创新能力提升工程项目(2014QNJJ-013)、国家科技支撑计划项目(2014BAD11B00)、国家现代农业产业技术体系四川油菜创新团队项目(川农业函[2014]91号)和四川省财政基因工程项目(2016ZYPZ-013)资助。
(dinghuiliu@163.com)
陈红琳(1984—),女,四川大英人,硕士研究生,助理研究员,主要从事油菜高产栽培生理生化研究。E-mail: chenhl0107@163.com
猜你喜欢
今日农业(2022年1期)2022-11-16
电脑迷(2022年10期)2022-11-08
安徽农业科学(2022年16期)2022-09-02
农业工程学报(2022年5期)2022-06-22
中国麻业科学(2021年5期)2021-12-02
农技服务(2021年6期)2021-09-22
麦类作物学报(2021年4期)2021-05-25
海峡姐妹(2020年6期)2020-07-25
海峡姐妹(2020年1期)2020-03-03
领导决策信息(2017年13期)2017-06-21
