
刈割对暖性草丛草地碳交换的影响
——以渝北岐山草地为例
2017-07-19 10:17王丽华
草原与草坪 2017年3期
王丽华,刘 尉
(1.阿坝师范学院,四川 汶川 623002; 2.成都市农林科学院,四川 成都 611130)
刈割对暖性草丛草地碳交换的影响
——以渝北岐山草地为例
王丽华1,刘 尉2
(1.阿坝师范学院,四川 汶川 623002; 2.成都市农林科学院,四川 成都 611130)
试验以南方改良后的暖性草丛草地为研究对象,探讨刈割对南方草地生态系统碳交换的影响。结果表明:(1)在生长季(4~9月),对照和刈割草地均处于CO2吸收阶段,但刈割草地对CO2的吸收量低于对照草地(P<0.01);在非生长季(10月~次年3月),对照和刈割草地均处于碳排放阶段,但刈割草地CO2排放量显著低于对照组(P<0.01),在全年尺度上,刈割草地净生态系统碳交换量(NEE)与对照草地无显著差异,且均为碳汇草地;(2)刈割草地生态系统呼吸(ER)和土壤呼吸(RS)在个别月份与对照相比有所降低,但在全年尺度上无影响;(3)生长季时,刈割草地生态系统总初级生产力(GEP)高于对照草地(P<0.05),但在非生长季和全年水平下无显著差异(P>0.05)。在生长季和非生长季草地生态系统碳交换量在刈割下有所变化,但在全年水平上效果并不显著。说明草地对外界干扰有一定的自我调适功能,适度刈割并不会影响草地生态系统碳循环。
刈割;草地生态系统碳交换;生长季节;非生长季节
如何利用陆地生态系统进行固碳减排的活动成为举世关注的问题之一,然而人类活动的不断干扰,陆地生态系统一旦遭到破坏,其固碳能力也随之下降,所以保护陆地生态系统的固碳潜力应优先考虑管理措施。在陆地生态系统中,草地生态系统在全球碳循环和气候调节中起着重要作用[1]。我国草地生态系统碳储量约占世界草地生态系统碳储量的8%[2],我国草地单位面积碳密度要高于世界平均水平,因此,在世界生态系统碳储量中具有重要作用[3]。
草地生态系统中CO2的交换和碳的吸收是植物对碳的光合固定、异养和自养呼吸以及土壤碳储存的结果。而刈割作为人为干扰因素改变了草地植被群落、冠层结构以及水分和地热传递等,此时草地的净辐射平衡和能量分配也随之产生变化。因此,刈割通过对第一性生产力、营养物质循环、有机质分解状况以及植物种间的竞争关系等的影响进而影响了大气与草地之间的碳通量[4]。
草地净生态系统碳交换量(Net ecosystem exchange,NEE)是草地生态系统中光合作用与呼吸作用之间的平衡,它的大小体现了生态系统吸收大气中CO2能力的高低。NEE为负值代表草地生态系统作为碳汇,NEE为正值,则为碳源[5]。生态系统总初级生产力(GEP,Gross ecosystem exchange)是指单位时间内植物通过光合作用途径固定的光合产物量或有机碳总量。草地植物通过光合作用产生GEP,将能量和CO2转化为有机碳和能量,作为草地碳循环过程的起始水平,同时也是草地碳循环的基础。生态系统呼吸(ER,Ecosystem repiration)包括了植物冠层呼吸和土壤呼吸,刈割在改变冠层群落的同时也改变群落的呼吸作用,地上生物量的降低伴随着植物呼吸器官的减少,因此,植物冠层呼吸作用与植物地上生物量具有正相关关系。草地土壤呼吸是草地生态系统中释放CO2的主要途径,同时也是土壤碳素同化和异化平衡的结果[6],研究土壤呼吸对于预测全球碳素平衡以及了解草地生态系统碳循环具有十分重要的意义。对土壤CO2排放的研究中,张宪洲等[7]报道,土壤温度是决定土壤CO2排放大小的主导因子,而刈割通过减少地上现存生物量和光合组织,使其光合速率下降,增加地表裸露面积致使土壤温度增加,提高土壤和根系的呼吸,对草地碳循环产生重要影响。然而草地生态系统的复杂性以及人为干扰等因素,使得草地生态系统碳通量存在不确定性。丁娓等[8]报道,放牧强度是草地从碳汇转变为碳源的主要原因。张梓瑜等[9]则认为利用方式是影响碳循环的主要因素(放牧下群落净CO2气体交换量日变化表现为生态系统对碳的吸收,而围封和割草则以碳释放为主)。众多研究中,均为生长季节时期人为干扰下对草地生态系统的研究,而对于无干扰下非生长季节下草地生态系统碳循环的研究较少。此外,国内外大量关于刈割的研究已经肯定了适度的刈割有利于草地整体生产力的提高[10],即超补偿生长现象的存在。我国南方天然草地占南方13省总土地面积的26%,且具有良好的水热条件以及单产量高等特点[11-12],具有较高的补偿性生长和固碳潜力。然而,有关我国南方草地碳循环的研究较少,而刈割作为南方草地主要的利用方式之一,在外界干扰较为频繁的情况下,对我国南方草地碳循环的研究显得尤为重要。
1 材料和方法
1.1 研究地概况
试验地设在重庆东北边的云阳清水乡的岐山草地(N 30°35′04.33″,E 108°54′37.23″)进行,该区域海拔1 600 m,年均温度10℃,年降水量1 200 mm,雨期主要集中在4~9月,无霜期240 d,冬春冷凉干旱,为典型的中国南方喀斯特岩溶山区草地,草地整体坡度20°,土壤类型为黄壤土,地表凋落物及立枯物较少,无砾石、无风蚀、无水蚀及盐碱斑,因此,试验区域从气候到地形等均具有南方草地的一些典型特征。此外,试验草地类型为南方暖性草丛的改良草地,在2010年进行鸭茅,白三叶补播,群落组成物种包括禾草类的优势种五节芒(Miscanthusfloridulus)、鸭茅(Dactylisglomerata)、多年生黑麦草(Loliumperenne)等;豆科的优势种为白三叶(Trifoliumrepens)等,草地利用方式为打草场,利用强度为轻度利用(每年夏季用刈割1~2次)。
1.2 生态系统碳换测定方法
2010年10月,在草地内选择2个100 m×100 m的样地(能代表该草地类型)围栏后进行刈割和对照处理,其中刈割样地每年5月进行1次齐地刈割处理(刈割强度为100%,留茬高度为0 cm),在刈割后将刈割掉的生物量清除,对照样地不进行处理。2012年12月底,选择植被类型均匀的地段,沿100 m样带每间隔20 m放置一个金属框及土壤环,对照区和刈割区分别放置金属框(0.5 m×0.5 m)和土壤环各5个(金属框与土壤环相距不超过50 cm),安置好金属框及土壤环后在2013年1~12月的每月中旬,在天气晴朗时用Li-840测量净生态系统交换量(NEE)、生态系统呼吸速率(ER),土壤呼吸速率(Rs),每次测定时间为上午的9∶00~11∶00。
NEE测定方法是将同化箱放在底座(金属框)上密封扣好,在记录好箱体内的温度后启动Li-840自动记录程序(每1秒记录1次数据),记录120 s内箱体内CO2和H2O的浓度变化,测量结束后再次记录箱体内的温度。
ER的测定是在完成NEE测定后进行,将同化箱密封扣打开并将同化箱冲着风向,使箱体内的CO2和H2O尽快恢复至大气水平。恢复后,将同化箱放置在底座上,盖上遮阳布,记录箱体内的CO2和H2O的浓度变化,时间同样为120 s,并记录始、末箱内温度。
土壤呼吸测定采用Li-8100测定系统,测定频次、时间与箱式法测定相同,在测定NEE和ER后进行。为保证数据准确,每次测定前将土壤环内的动植物体进行清除。
1.3 数据分析与处理
试验数据分析采用Microsoft Office Excel 2007和Origin 8.5软件进行数据整理和作图。对照和刈割草地碳储量部分采用成对T检验方法来分析两者之间是否具有显著性差异(P<0.05)。统计分析均由SPSS 17.0统计分析软件完成。生态系统呼吸测定中,采用公式计算NEE和ER[13]。
Fc=[VPav(1000-Wav)/R×S(Tav+273)]×dc/dt
(1)
GEP=ER-NEE
(2)
式中:Fc为CO2交换速率,V为箱体的体积:[(同化箱长宽(箱体高度+底座露出地上部分的高度)];Pav为测量期间箱体内的平均大气压强(kPa);Wav是测量期间箱体内的水分气压(mmol/mol);R代表大气常数8.314 J/(mol·K);S是同化箱的面积(m2);Tav是测量期间箱体内的平均温度。CO2浓度变化速率(dc/dt)采用线性拟合的方法计算,即斜率。生长季、非生长季即年度NEE、ER、Rs和GEP分别为4~9月、10~翌年3月和1~12月的所测得数据的平均值。
2 结果与分析
2.1 刈割对草地生态系统NEE的影响
通过对2013年不同月份草地生态系统NEE测定后发现(图1),NEE在生长季(4~9月)均值为负,而非生长季(10月至次年4月)均为正值,但年度NEE均值为负。在整个生长季,对照组草地生态系统NEE变化趋势基本上为“U”型,在2013年7月达到最大值,为-7.49 μmol/(s·m2),之后开始逐渐降低。NEE在10月达到最大值,为3.40 μmol/(s·m2),之后开始逐渐降低。刈割处理样地的NEE生长季变化过程与对照草地走势基本一致,在7月达到最低值,为-6.34 μmol/(s·m2),但是6月和8月时与5月和9月相比有小幅度的增加。与未刈割草地相比,刈割草地NEE在生长季增加了25.20%,而在非生长季却降低了42.68%,但就整个年度NEE均值分析,两者差异并不明显(P<0.05)。
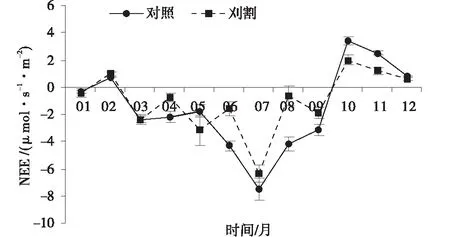
图1 未刈割和刈割处理下的净生态系统交换Fig.1 Variations of NEE in unclipped and clipped grasslands
2.2 刈割对草地呼吸ER的影响
草地呼吸ER具有明显的季节变化,对照组和刈割草地ER之间的季节变化具有较好的一致性。两种处理下,草地在生长季和非生长季ER变化趋势总体为单峰曲线。在生长季时,对照组6月达到峰值,为11.67 μmol/(s·m2),而刈割草地在5月达到峰值,为8.07 μmol/(s·m2),之后便开始逐渐降低。在非生长季,对照组草地的ER在1月下降到最小值,为-0.91 μmol/(s·m2),而刈割草地则在12月下降到最小值,为0.61 μmol/(s·m2),之后开始逐渐增加(图2)。
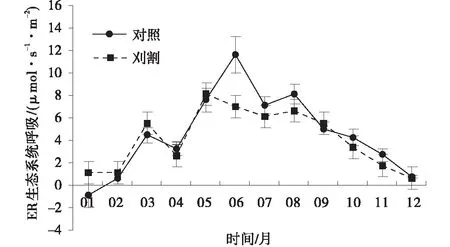
图2 未刈割和刈割处理下的群落生态系统呼吸速率Fig.2 Variations of ER in unclipped and clipped grasslands
2.3 刈割对土壤呼吸速率RS的影响
草地土壤呼吸RS具有明显的季节变化,其总体变化趋势呈现为单峰型,且对照和刈割草地的Rs变化趋势一致。2个处理下,草地RS从3月开始增加,到8月时达到最大值,为18.23 μmol/(s·m2)和15.58 μmol/(s·m2),在达到最大值后开始下降,直到2月时下降到最低值(图3)。刈割草地在个别月RS平均值低于对照组草地,但2个处理下RS在生长季和非生季平均值上无显著差异(P>0.05)。
2.4 刈割对草地总初级生产力GEP的影响
草地总初级生产力GEP有明显的季节变化,对照组草地和刈割草地之间的季节变化趋势一致,草地GEP从3月开始增加,在7月达到最大值,分别为12.45和14.61 μmol/(s·m2),8月之后开始下降并在2月下降到最低值。在生长季的7、8和9月时,刈割草地GEP均高于对照组草地(图4)。在非生长季节(P>0.05),刈割草地GEP在10月和11月略高于未刈割草地GEP,其余月之间没有显著差异。从草地GEP年平均值分析,刈割草地GEP对照组草地无显著差异(图5,P>0.05)。
3 讨论
刈割草地NEE的季节变化规律与对照草地基本相同,说明刈割并未改变NEE的季节变化形式,这一研究结果与徐丽君等[14]研究结论一致。从对照和刈割草地的全年NEE平均值分析,草地生态系统总体表现为碳吸收,且刈割并未对草地NEE造成影响。然而
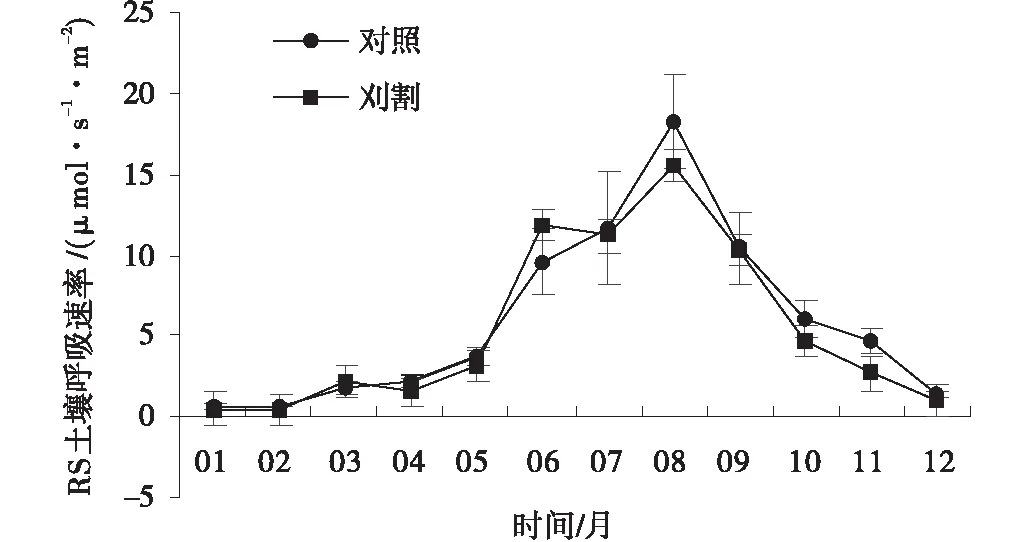
图3 对照和刈割处理下的土壤呼吸速率Fig.3 Variations of Rs in unclipped and clipped grasslands
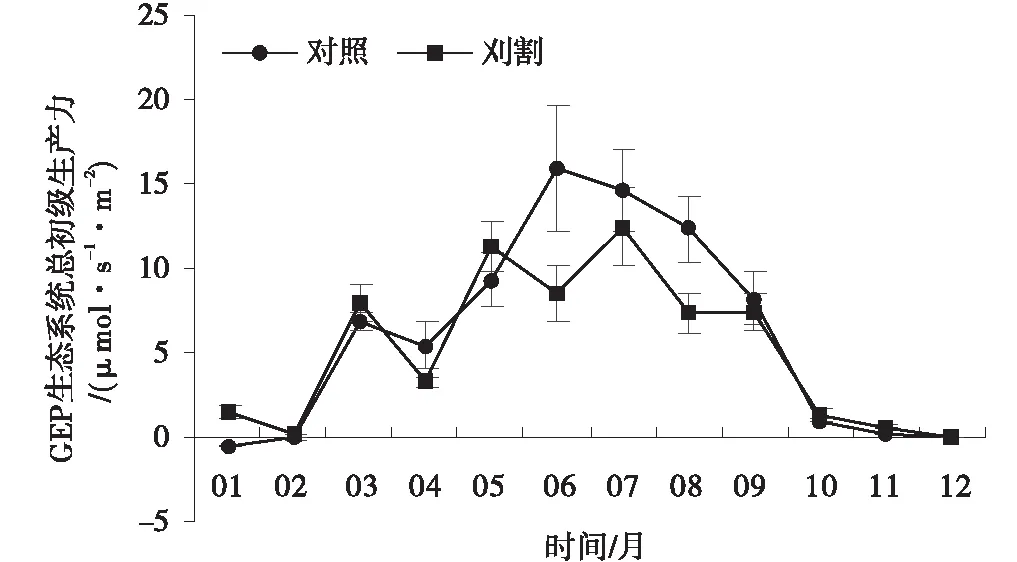
图4 对照和刈割处理下的草地总初级生产力Fig.4 Variations of GEP in unclipped and clipped grasslands
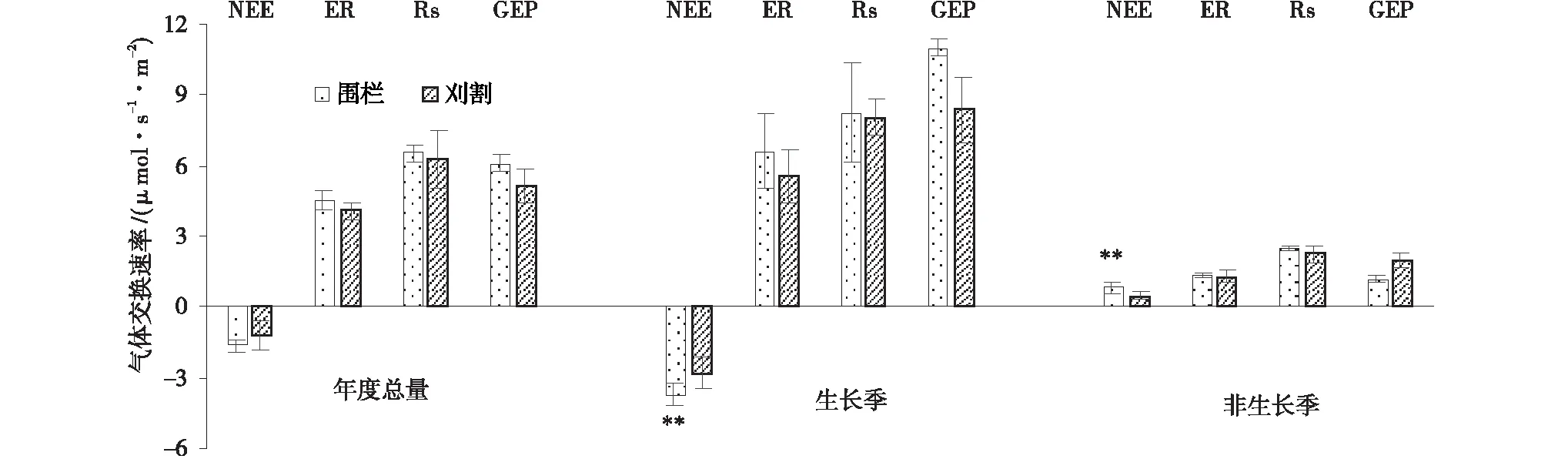
图5 未刈割和刈割处理下的群落生长季与非生长季及年度总气体交换速率Fig.5 Average value of NEE,ER,Rs,GEP in unclipped and clipped grasslands 注:刈割与对照处理间*表示差异显著(P<0.05),**表示差异极显著(P<0.01)
生长季和非生长季草地NEE平均值却存在差异。在生长季(4~9月),对照和刈割草地NEE均表现为负值,其变化趋势呈“U”型,且均在7月达到NEE最大值,从生长季各月NEE变化分析,随着温度和降水量的增加(土壤含水量增加),草地植物营养物质的合成和积累增多,极大的促进了植物光合作用,并且提高了植物对CO2的吸收量[15]。因此,草地生态系统在此时期处于碳吸收阶段。试验中,刈割草地NEE在6~9月时普遍低于对照草地,说明经过刈割处理后,一方面可能是因为草地生物量锐减,植物群落整体光合能力下降,因此对CO2的吸收量降低[16]。
草地植物在进入非生长季节后,植物体逐渐衰老枯萎,光合作用日渐降低,草地生态系统对碳的吸收活动减弱,因此,在非生长季时草地通常处于碳排放过程[14]。试验中,非生长季对照和刈割草地NEE平均值表现为正值,表现为CO2源。但对照草地NEE显著大于刈割草地,说明此时生态系统在对照和刈割下虽然同为碳排放阶段,但刈割草地在10~11月时CO2排放量小于对照草地。这是因为草地植物在刈割后,会将根部贮藏的碳水化合物向地上转移以恢复地上光合组织的缺失,然而这种内部资源分配方式的改变只是植物受损后的一种短期响应[17],植物为了从刈割伤害中恢复正常的生长,恢复其获取碳的能力而延迟叶片的衰老[18],增加了植物对CO2的吸收量,从而导致刈割草地NEE变小。类似的研究结果出现在Hartemink[19]、候扶江[20]和Simith等[21]的相关研究中。
在草地生态系统中,草地土壤呼吸速率(Rs)受土壤温度和土壤含水量的共同影响,然而在生长季时受土壤含水量影响偏大,而非生长季时则受土壤温度影响较大[22]。试验中,刈割和对照草地Rs具有明显的季节变化。草地植物返青开始后,温度和降水量逐渐增加致使草地土壤温度和含水量逐渐升高,因此草地Rs开始增加,到8月温度和降水量达到顶峰时,Rs也达到了最大值[23]。然而,进入10月后,草地降水量减少,此时草地土壤Rs主要受土壤温度的影响,因气温下降导致了根系呼吸和土壤微生物代谢活动减弱,土壤呼吸活动逐步降低,在2月降到最低值。
有关放牧或刈割对土壤呼吸的研究已有很多,但结论并不一致,有降低[24],有升高[25]以及无影响[26]。试验中,对照草地Rs在个别月要高于刈割草地,是因草地土壤环境受地上植被的影响,刈割后草地植被覆盖率减少,地上生物量累积和输送到土壤中的养分也随之减少,因此植物根系和土壤微生物呼吸作用减弱。但从全年、生长季和非生长季平均值草地Rs分析,刈割草地土壤呼吸在个别月虽然有所降低,但并不显著,可能是由于刈割年限较短,因此并未对草地造成明显的影响。与郭明英等[27]对刈割后呼伦贝尔草甸草原土壤呼吸特性的研究结果一致。
草地生态系统呼吸(ER)包括群落冠层呼吸和土壤呼吸,群落冠层呼吸主要受地上生物量的影响[28],因此具有较大的季节性动态变化。植物光合速率通常会随外界温度的升高而增强[29],温度既影响植物的呼吸作用也影响其光合能力。对2013年全年草地ER进行测定后发现,生长季和非生长季草地ER变化呈单峰曲线,对照草地在生长季时草地ER随温度增加而增加,在7月达到最大值后下降,并在1月(非生长季)下降到最低值,这是由于在生长季时随着温度的增加,植物快速生长、累积生物量增多,使植物冠层呼吸作用增强,导致了草地生态系统呼吸增加;在非生长季时气温较低,光合能力减弱,植物衰老枯萎地上生物量逐渐降低,草地生态系统呼吸也随之降低。在刈割处理下,草地生态系统呼吸的季节变化规律与对照草地一致,但在生长季节,刈割草地ER在5月达到最大值,之后便逐渐降低,这是因为刈割将部分地上生物量移除后,草地地上现存量处于较低水平,植被冠层叶面积减少,群落冠层呼吸也随之降低。在对照和刈割处理下,刈割虽然在个别月对草地生态系统呼吸有一定的影响,但在草地生长季、非生长季以及全年水平上并无显著差异,这进一步体现了我国南方草地对刈割有较强的抗干扰能力。
对照和刈割草地生态系统总初级生产力(GEP)在生长季和非生长季下变化趋势总体呈单峰曲线,且两处理间变化趋势基本一致。草地GEP与植被状况密切相关,因此天气回暖后,气温逐渐上升,植物进入营养生长阶段,地上生物量开始慢慢累积,在7月份时日光合有效辐射达到最大值,气温达到峰值且当地降水丰富促使植物光合能力达到最大值[30],此时地上生物量处于年度最高值,致使草地GEP同样达到了峰值。之后温度和降水量下降,草地植被生物量减少,因此草地GEP逐渐降低,到2月份时下降到最低值。
前人对放牧的研究报道[31-32],放牧后的草地可以消除一些限制植物生长的因素(如老叶片、立枯物等),提高植物的光合补偿能力,并有利于植被的更新及再生长,维持甚至提高草地最大的、持续的生产力。此外,研究者发现[33]植物的补偿性生长与剩余叶片的叶面积指数(LAI)有关,刈割作用改变了草地植被的冠层结构,从而改变了植被对光的截获和吸收,草地GEP也随之发生变化。当草地群落覆盖率高达100%时,植物叶片对光的截获量较大,适度的放牧或刈割降低了LAI(在一定阈值范围内),植物可通过再生萌蘖得以平衡和补偿,此时草地现存量较高;当LAI下降到阈值范围之外时,植物的再生能力较弱,草地地上现存量降低[34]。Zhao等[35]对黑麦草进行刈割梯度试验后发现,20%刈割强度下黑麦草剩余出现了明显的光和补偿作用,其地上相对生长速率也有所提高并出现了超补偿生长,而在60%刈割强度下,并没有出现明显的光合补偿效果,且相对生长速率也较低。试验中草地生长季GEP平均值高于对照草地,而在非生长季无显著差异,说明适度的刈割可能刺激了植物的超补偿生长,从而提高了草地生产力水平,因此植物的超补偿性生长有利于草地生态系统初级生产力的提高。这一结果与Chimner[36]、董全民等[37]的研究结果一致。
4 结论
刈割并没有改变草地在生长季和非生长季的响应模式,刈割与对照草地均在生长季节表现为碳汇,而在非生长季节为碳源,在全年平均值表现为碳汇。此外,刈割同样不会改变草地NEE、ER、RS及GEP固有的季节变化趋势。在生长季时期,刈割草地为碳净吸收阶段,但其对CO2的吸收量低于对照草地,然而在非生长季时期为碳排放阶段,其对CO2的排放量同样低于对照草地,因此在全年水平上并无显著差异,依旧表现为碳汇草地。刈割草地的ER和Rs虽然在个别月有所降低(P<0.01),但在不同非生长季节及全年尺度上看同样无显著差异。试验中,刈割草地GEP在生长季节时的平均值高于对照草地,而在非生长季和全年水平上无显著差异,说明刈割刺激了植物的生长从而提高了草地净初级生长力水平。
[1] Reeder J D,Schuman G E.Influence of livestock grazing on C sequestration in semiarid mixed grass and short grass rangelands[J].Environmental Pollution,2002,116:457-463.
[2] Ni J.Carbon storage in terrestrial ecosystems of China:Estimates at different spatial resolutions and their responses to climate change[J].Climatic Change,2001,49:339-358.
[3] 樊江文,钟华平,梁飚,等.草地生态系统碳储量及其影响因素[J].中国草地,2003,25(6):51-58.
[4] Flerchinger G N,Sauerb T J,Aikenc R A.Effects of crop residue cover and architecture on heat and water transfer at the soil surface[J].Geoderma,2003,116:217-223.
[5] 金相灿,屠清瑛.湖泊富营养化调查规范[M].北京:中国环境科学出版社,1990:286-302.
[6] 崔晓勇,陈佐忠,陈四清.草地土壤呼吸研究进展[J].生态学报,2001,21(2):215-325.
[7] 张宪洲,石培礼,刘允芬,等.青藏高原高寒草原生态系统土壤CO2排放及其碳平衡[J].地球科学,2004,34(增刊Ⅱ):193-199.
[8] 丁娓,孙霞,贾阳阳,等.不同放牧强度对天山北坡草原草地生态系统碳交换速率的影响[J].新疆农业大学学报,2014,37(3):197-202.
[9] 张梓瑜,龚吉蕊,刘敏,等.温带草原不同土地利用方式下优势种植物和生态系统的气体交换[J].植物生态学报,2013,37(8):718-727.
[10] Painter E,Belsky A J.Application of herbivore optimization theory to rangelands of the western United States[J].Ecological Applications,1993,4:2-9.
[11] 吕世海.我国南方草地资源现状及其发展前景[J].四川草原,2005(6):37-41.
[12] 滕永青,李斌斌,王友国,等.合理开发利用南方草地资源促进草地畜牧业发展[J].中国畜牧兽医,2008,35(4):136-140.
[13] 周广胜.全球碳循环(第一版)[M].北京:气象出版社,2003:162-189.
[14] 徐丽君,唐华俊,杨桂霞,等.贝加尔针茅草原生态系统生长季碳通量及其影响因素分析[J].草业学报,2011,20(6):287-292.
[16] 张文丽,陈世苹,苗海霞,等.开垦对克氏针茅草地生态系统碳通量的影响[J].植物生态学报,2008,32(6):1301-1311.
[17] Reichman O J,Smith S C.Responses to simulated leaf and root herbivory by a biennial,Tragopogondubius[J].Ecology,1991,72:116-124.
[18] Welter S C.Arthropod impact on plant gas exchange[M].1989:135-150.
[19] Hartemink N,Jongejans E,Kroon H D.Flexible life history responses to flower,rosette bud removal in three perennial herbs[J].Oikos,2004,105:159-167.
[20] 侯扶江.放牧对牧草光合作用、呼吸作用和氮、碳吸收与转运的影响[J].应用生态学报,2001,12(6):938-942.
[21] Smith S E.Variation in response to defoliation between populations ofBoutelouacurtipendulavar.Caespitosa(Poaceae) with different livestock grazing histories[J].American Journal of Botany,1998,85(9):1266-1272.
[22] 郭明英,徐丽君,杨桂霞,等.不同刈割间隔对羊草草甸草原割草地土壤呼吸的影响[J].草原与草坪,2010,30(6):10-14.
[23] Gao Y Z,Marcus G,Lin S,etal.Belowground net primary productivity and biomass allocation of grassland in Inner Mongolia is affected by grazing intensity[J].Plant Soil,2008,307:41-50.
[24] Wang Z W,Jiao S Y,Han G D,etal.Soil Respiration Response to Different Stocking Rates onStipabrevifloraGriseb.Desert Steppe[J].Journal of Inner Mongolia University,2009,40(2):186-193.
[25] Stark S,Strommer R,Tuomi J.Reindeer grazing and soil microbial processes in two suboceanic and two subcontinental tundra heaths[J].Oikos,2002(97):69-78.
[26] Frank A B.Carbon dioxide fluxes over a grazed prairie and seeded pasture in the Northern Great Plains[J].Environmental Pollution,2002(116):397-403.
[27] 郭明英,卫智军,徐丽君,等.不同刈割年限天然草地土壤呼吸特性研究[J].草地学报,2011,19(1):51-57.
[28] 宗宁,石培礼,蔣婧,等.短期氮素添加和模拟放牧对青藏高原高寒草甸生态系统呼吸的影响[J].生态学报,2013,33(19):6191-6201.
[29] 林栋,吕世海,冯朝阳,等.华北山地阳坡中生灌草植被对CO2浓度和温度变化的光合响应[J].草业科学,2008,25(4):135-140.
[30] 戚秋慧,盛修武,姜恕.内蒙羊草草原群落光合速率日变化及其与环境条件之间相互关系的初步研究[J].生态学报,1983,3(4):333-340
[31] 汪诗平,王艳芬.不同放牧率下糙隐子草种群补偿性生长的研究[J].植物学报,2001,43(4):413-418.
[32] 朱志红,王刚,赵松岭.不同放牧强度下矮嵩草无性系分株种群的动态与调节[J].生态学报,1994,14(1):40-45.
[33] Hilbert D W.Optimization of plant root/shoot ratios and internal nitrogen concentration[J].Annals of Botany,1990,66:91-99.
[34] 董世魁,丁路明,徐敏云,等.放牧强度对高寒地区多年生混播禾草叶片特征及草地初级生产力的影响[J].中国农业科学,2004,37(1):136-142.
[35] Zhao W,Chen S P,Lin G H.Compensatory growth responses to clipping defoliation inLeymuschinensis(Poaceae) under nutrient addition and water deficiency conditions[J].Plant Ecol,2008,196:85-99
[36] Chimner R,Welker J.Influence of grazing and precipitation on ecosystem carbon cycling in a mixed-grass prairie[J].Pastoralism,2011,1(1):1-15.
[37] 董全民,赵新全,马玉寿.放牧强度对高寒混播人工草地群落特征及地上现存量的影响[J].草地学报,2012,20(1):10-15.
Effect of clipping on CO2exchange in warm-temperature tussock——A case study in the north part of Chongqing,China
WANG Li-hua1,LIU Wei2
(1.AbaTeachersUniversity,Wenchuan623002,China; 2.ChengduAcademyofAgricultureandForestrySciences,Chengdu611130,China)
The aim of this study is to provide a scientific basis for the accurate assessment of the potential of carbon fixation by exploring the relation between CO2exchange of improved warm-temperature tussock and clipping.The result showed that 1) in growing season,unclipped and clipped grassland were both in carbon sequestration period,but CO2absorption amount of clipped grassland was lower than the unclipped.In non-growing season,unclipped and clipped grassland were both in carbon emission period,but the CO2emission amount of clipped grassland was lower than the unclipped.Therefore,NEE of clipped grassland was not affected by clipping,and clipped and unclipped grassland both were carbon sink;2) As compared with unclipped grassland,ER and RS of clipped grassland decreased in several months,but the whole effect was not significant;3) In growing season,GEP of clipped grassland was higher than unclipped,but no significant difference was found in non-growing season and full year.
clipping;CO2flux in grassland ecosystem;growing season;non-growing season
2017-04-10;
2017-04-28
四川省教育厅重点项目(17ZA0001);阿坝师范学院校级重点项目(ASA16-05)资助
王丽华(1986-),女,河南郑州人,博士,讲师,从事草地资源与生态学研究。 E-mail:464608579@qq.com 刘尉为通讯作者。
S 812
A
1009-5500(2017)03-0022-07
猜你喜欢
今日农业(2020年19期)2020-12-14
幼儿100(2020年31期)2020-11-18
快乐作文(1.2年级)(2019年3期)2019-09-10
疯狂英语·初中版(2019年4期)2019-09-10
英语文摘(2019年5期)2019-07-13
幼儿画刊(2018年10期)2018-10-27
小太阳画报(2018年6期)2018-05-14
小雪花·成长指南(2016年2期)2016-03-16
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
