
桉树焦枯病菌CpSit1基因的鉴定与表达
2017-07-18 11:12:05刘宏毅叶小真陈慧洁李慧敏杨泽慧冯丽贞
福建农林大学学报(自然科学版) 2017年4期
刘宏毅, 叶小真, 陈慧洁, 李慧敏, 丁 奕, 杨泽慧, 冯丽贞
(1.福建农林大学林学院;2.福建农林大学金山学院,福建 福州 350002)
桉树焦枯病菌CpSit1基因的鉴定与表达
刘宏毅1, 叶小真2, 陈慧洁1, 李慧敏1, 丁 奕1, 杨泽慧1, 冯丽贞1
(1.福建农林大学林学院;2.福建农林大学金山学院,福建 福州 350002)
利用Blast及TCDB数据库对桉树焦枯病菌(Calonectriapseudoreteaudii)的Cpsit1基因进行鉴定;再利用SMART数据库和ProtParam、TMHMM、PHD、ProComp 9.0在线分析工具对Cpsit1基因编码蛋白的理化性质、跨膜螺旋、蛋白质二级结构和亚细胞定位进行预测分析;最后采用qRT-PCR方法对CpSit1基因在焦枯病菌侵染桉树过程中的表达情况进行分析.结果表明:CpSit1基因长度为1 780 bp,其编码的蛋白序列共有592个氨基酸残基组成,并将其鉴定为铁载体-铁:H+同向转运蛋白.其保守结构域为MFS_1;共含有14个跨膜螺旋;在二级结构中α螺旋占45.10%,延伸链占26.01%,无规则卷曲占28.89%.通过qRT-PCR相对定量的方法分析焦枯病菌侵染桉树24、48和72 h后CpSit1基因的表达情况,结果显示,CpSit1基因在这3个时段均发生上调表达,但24 h的表达量明显大于48和72 h.说明桉树焦枯病菌CpSit1基因在病原菌侵染寄主的过程中通过调控铁载体-铁化合物的转运来完成铁元素的摄入,协助其在寄主中的定植.
桉树; 焦枯病; 铁载体转运蛋白; 生物信息学分析; 表达分析
在大多数原核生物和所有真核生物中,铁元素是各种生命活动的必需元素.地壳中,铁元素的储量丰富,但是其难溶性大大限制了真菌对铁的直接吸收.在生境中铁元素含量较低时,大多数微生物和部分植物通过分泌铁载体与Fe3+结合为铁载体-铁化合物来获取铁元素[1].大多数真菌存在4种异羟肟酸型铁载体,即镰孢氨酸、粪生素、铁色素和罗丹明酸,主要由非核糖体肽合成酶(NRPSs, non-ribosomal peptide synthetases)合成[2].铁载体-铁转运蛋白属于MFS超家族,在缺铁的生境下通过转运铁载体-铁化合物为微生物的生长发育提供铁元素.SIT转运蛋白具有典型的MFS结构域,家族成员大多数由400~600个氨基酸残基组成,N端和C端均位于胞内[3].目前已知的SIT转运蛋白均含有14个跨膜α螺旋,多出的2个跨膜螺旋是由胞内中间的环状区插入膜中产生的[4].大部分微生物都通过独特的代谢途径来获取生境中的铁元素.大多数病原真菌主要通过还原铁吸收系统或通过分泌铁载体来获取铁元素.目前研究证明SIT转运蛋白作为铁载体-铁化合物在运输载体中发挥着重要作用,如禾谷镰刀菌(Fusariumgraminearum)FgSit1基因编码的SIT转运蛋白负责调控铁载体-铁化合物FC-Fe3+的吸收[5];构巢曲霉(Emericellanidulans)的SIT转运蛋白MIRB与铁载体-铁化合物三乙酰镰孢氨酸的转运有关[6];酿酒酵母(Saccharomycescerevisiae) Arn1p转运蛋白通过转运生境中的铁载体-铁化合物来获取铁元素[7].
桉树焦枯病(Calonectriapseudoreteaudii)是热带和亚热带地区桉树种植区最为严重的病害之一,严重影响桉树产业的发展[8].桉树焦枯病菌是由丽赤壳属(Calonectria)真菌引起的病害,其无性态为帚梗柱孢属(Cylindrocladium)真菌[9].据统计,Calonectria现有集群13个,共71种.研究[10-12]表明C.pseudoreteaudii是福建省内最早发现的、分布最广、致病力最强的病原菌株,并对C.pseudoreteaudii基因组测序及桉树组织诱导48 h的焦枯病菌转录组进行高通量测序,发现Cp_Cap02212基因发生上调倍数高达746倍.为了探讨该基因在焦枯病菌侵染桉树过程中所起的作用,本文通过多种生物信息学软件对其编码基因、蛋白结构、亚细胞定位等进行分析,并对其功能进行预测,最后通过实时荧光定量PCR对其在焦枯病菌侵染桉树24、48、72 h 3个阶段的表达情况进行分析,为揭示桉树焦枯病菌的致病机理提供参考.
1 材料与方法
1.1 供试材料
1.1.1 菌株 为C.pseudoreteaudiiYA51,由福建农林大学森林保护研究所于福建永安桉树焦枯病危害区采集分离获得.基因组NCBI 检索编号为MOCD01000000.
1.1.2 主要培养基及试剂 本试验选用桉树抗焦枯病品系尾细桉M1(Eucalyptusurophylla×E.tereticornis)的叶片对焦枯病菌(C.pseudoreteaudii)进行培养.含桉树组织PDB培养基:将桉树叶片研磨成粉末状,以0.5%质量分数加入200 mL的PDB培养基中,高温灭菌.总RNA提取、反转录PCR、实时荧光定量PCR的试剂盒均购自天根生化科技(北京)有限公司.
1.2 方法
1.2.1 生物信息学分析 以Cp_Cap02212的蛋白序列为探针序列,与NCBI的nr数据库进行Blastp比对,获得初步鉴定结果;再将Cp_Cap02212的蛋白序列与TCDB数据库进行比对,对其所属蛋白家族进行鉴定,并命名.
CpSit1转运蛋白的理化性质利用ProtParam进行分析;应用SMART数据库进行在线比对,分析其结构域;采用在线分析工具TMHMM分析跨膜螺旋;采用在线分析工具PBIL和PHD分析蛋白质二级结构,该分析基于ASTP和MaxHom.亚细胞结构定位预测采用ProComp 9.0在线工具.
1.2.2 系统进化树构建 从TCDB数据库中下载MFS家族典型亚家族的蛋白序列,通过MEGA7.0软件[13]进行多重比对,并采用临近法构建系统发育树,Bootstrap值设置为1 000次,其余参数均为默认值.
1.2.3 桉树焦枯病菌菌株培养 将桉树焦枯病菌接种于PDA培养基(琼脂质量分数1.5%)中,培养10 d后用直径为6 mm的打孔器取获菌饼;分别接种于含桉树组织的PDB培养基(含20 μL氨苄青霉素)中,作为处理组;不含桉树组织的作为对照组.每个处理组及对照组各有3个培养基,每个培养基中加入3个菌饼;然后置于摇床中摇晃培养,分别于24、48和72 h取出相应的对照组和处理组,用微孔滤膜收集菌丝,滤纸滤干后用锡纸包裹,于-80 ℃冰箱中保存.
1.2.4CpSit1基因表达分析 采用RNAprep Pure Plant Kit离心柱型试剂盒提取总RNA,提取完成后用Denovix公司生产的超微量核算浓度检测仪检测RNA浓度及质量;对于RNA浓度符合RT-PCR要求的处理组,用FastQuant RT Kit试剂盒进行反转录,样品于-20 ℃冰箱中保存.引物设计采用Beacon Design 7.9软件,委托生工生物工程有限公司合成.CpSit1基因的引物序列为F-GAAACCCAGCGATTGATG,R-TAACCTCCTTCCAGAACAG;内参基因(β-tublin)的引物序列为F-CTATCTTCCGTGGTAAGG,R-TAGGTGGAGTTCTTGTTC.
采用Talent qPCR PreMix试剂盒进行qRT-PCR,反应体系为20 μL体系.2×Talent qPCR Mix 10.0 μL;上游引物0.6 μL;下游引物0.6 μL;cDNA模板1.0 μL;50×ROX Reference Dye 0.4 μL;RNase-Free ddH2O 7.4 μL.加样完成后采用两步法进行扩增,95 ℃预变性30 min,1个循环;95 ℃变性5 s,60 ℃退火延伸15 s,40个循环.反应程序完成后采用2-△△CT方法进行分析.
2 结果与分析
2.1CpSit1基因鉴定
将Cp_Cap02212蛋白序列与nr数据库进行Blastp比对,发现与Phaeoacremoniumminimum的SIT转运蛋白序列的一致性高达64%.通过与TCDB数据库比对发现,Cp_Cap02212的蛋白序列与构巢曲霉SIT转运蛋白MIRB的蛋白序列的一致性为37%,相似性达到57.8%,推测其可能为铁载体-铁:H+同向转运蛋白,并命名为CpSit1.CpSit1基因的碱基序列长度为1 780 bp,位于基因组中Scaffold 21正链538882-541073的位置,该基因含有1个外显子,不含有内含子,其蛋白序列由592个氨基酸残基组成.
2.2 CpSit1蛋白理化性质及亚细胞定位

图1 CpSit1蛋白亲水性与疏水性分析Fig.1 Hydrophobicity and hydrophilicity analysis of CpSit1 in C.pseudoreteaudii
利用ProtParam在线分析CpSit1蛋白的理化性质,分子质量为66 294.60 u,理论等电点为6.61;该蛋白在280 nm处的光密度值为2.423~2.431,不稳定系数为28.28(<40),属于稳定蛋白;其平均疏水性为0.486,表明CpSit1蛋白为疏水性蛋白(图1).通过ProComp 9.0在线分析发现,CpSit1蛋白位于细胞膜上的可能性较高.
2.3 CpSit1蛋白结构
通过与SMART数据库比较发现,其在靠近N端78~501个氨基酸(E-value:3.4e-14)位置有一个MFS_1保守结构域,在靠近C端557~576个氨基酸的位置存在一个未知的跨膜区(图2).通过PHD对CpSit1蛋白二级结构(图3)的分析发现,α-螺旋占45.10%,延伸链占26.01%,无规则卷曲占28.89%.其N端和C端主要由无规则卷曲和延伸链组成,而α螺旋与无规则卷曲、延伸相间分布在中间位置.通过TMHMM分析发现,CpSit1蛋白的N端位于胞内的可能性高达99.427%,共有14个α-跨膜螺旋,具有典型的MFS折叠.CpSit1蛋白共有接近299个氨基酸残基位于跨膜螺旋上,这与二级结构预测中α螺旋所占比例相接近.前60个氨基酸残基位于跨膜螺旋的可能性几乎为零,说明其存在信号肽的可能性极低.以上结果表明桉树焦枯病菌的CpSit1蛋白具有明显的SIT转运蛋白的结构特征,进而可能具有SIT蛋白的活性和功能.

图2 CpSit1转运蛋白保守结构域
2.4 系统进化分析
通过TCDB数据库对MFS超家族进行检索,从中选取出7个亚家族,分别为4个铁载体(铁)转运蛋白、3个12跨膜螺旋药物(H+)反向转运蛋白、2个14跨膜螺旋药物(H+)反向转运蛋白、2个糖转运蛋白、2个硝酸盐亚硝酸盐转运蛋白、2个磷酸(H+)同向转运蛋白、1个阳离子(阴离子)同向转运蛋白,共16个MFS超家族蛋白(表1).如图4所示,各亚家族根据其分类落在不同的分支上,其中CpSit1转运蛋白与4个SIT转运蛋白位于同一分支上,分别为构巢曲霉(E.nidulans)的MIRB、MIRC蛋白和酿酒酵母(S.cerevisiae)的ARN1、ARN2蛋白.其中,CpSit1转运蛋白与MIRB位于同一分支上,说明这2个转运蛋白的亲缘关系较为接近,且与TCDB数据库鉴定结果较符合.SIT家族与DHA2家族位于同一分支,其余5个亚家族位于同一分支上.这可能与SIT、DHA2家族均含有14个跨膜螺旋有关,而其余5个亚家族的跨膜螺旋数量均为12个.

图3 CpSit1转运蛋白跨膜螺旋Fig.3 Transmembrane helix of CpSit1 transporter
2.5CpSit1基因表达
通过qRT-PCR相对定量的方法分析焦枯病菌侵
染桉树24、48和72 h后CpSit1基因的表达情况(图5),结果显示,CpSit1基因在24、48和72 h均发生上调表达,其中24 h上调倍数最高,达到23.99倍,明显高于48和72 h.48和72 h的上调倍数低于2倍,差异表达不显著.说明CpSit1基因主要在侵染初期和定植过程中通过调控铁载体-铁化合物的转运为焦枯病菌提供铁元素.
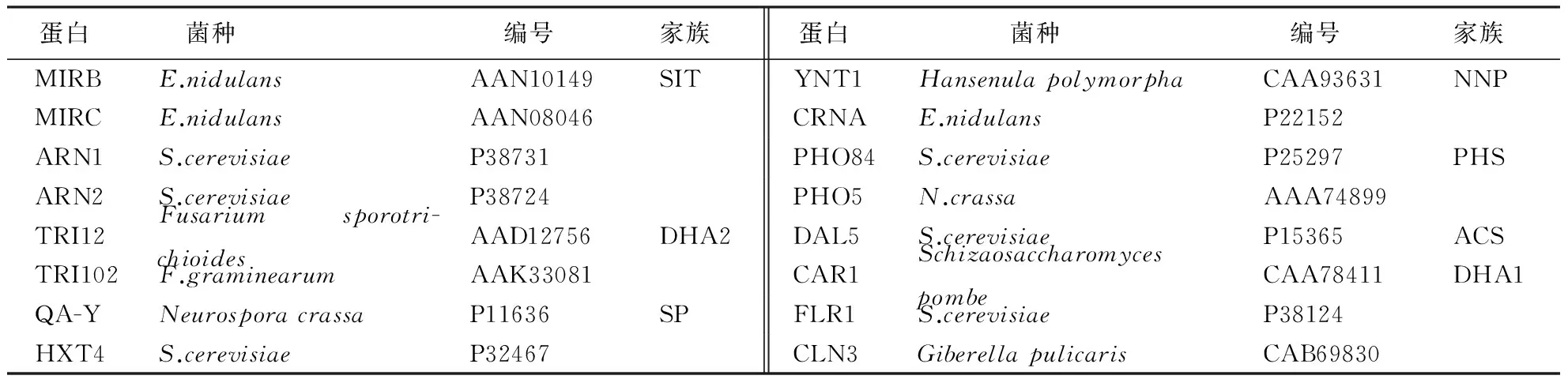
表1 MFS家族典型亚家族
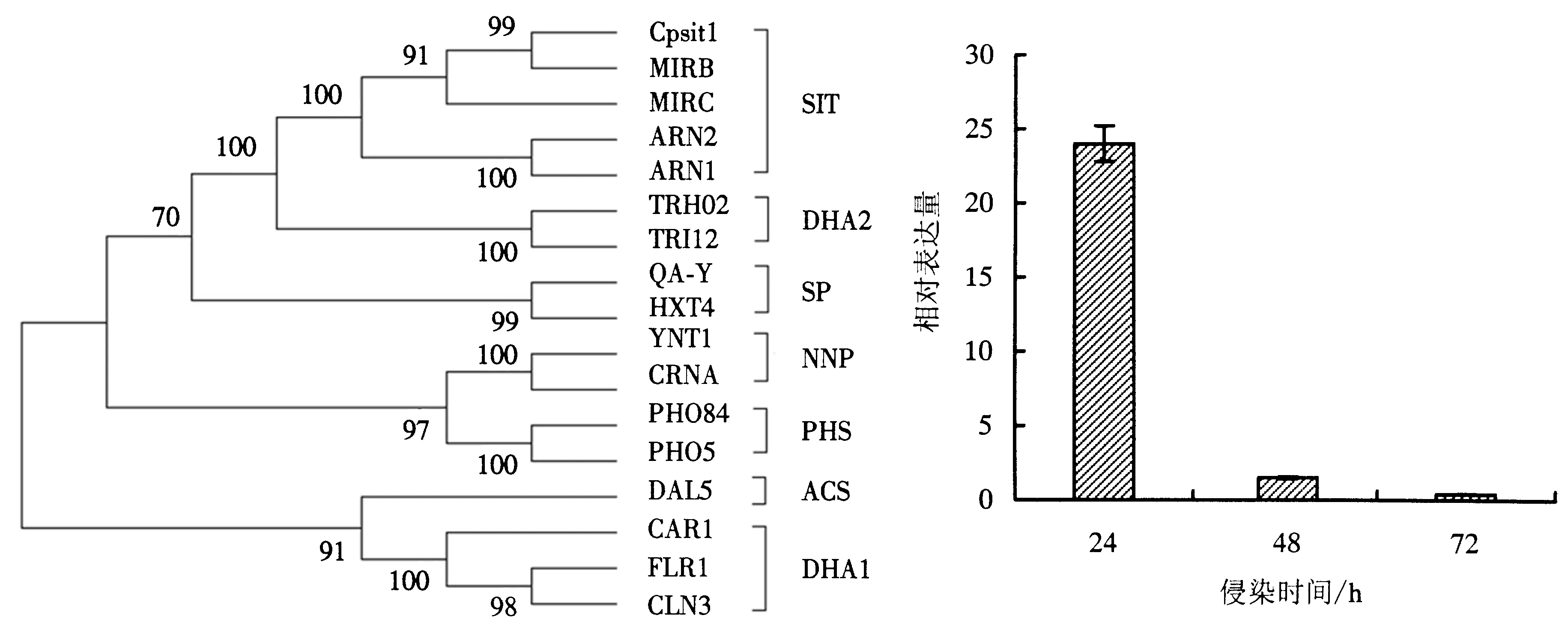
图4 系统发育分析
3 小结与讨论
本研究通过生物信息学方法鉴定出桉树焦枯病菌(C.pseudoreteaudii)的第1个SIT转运蛋白CpSit1,其蛋白序列共由592个氨基酸残基组成.TCDB数据库比对结果显示,其可能为铁载体-铁:H+同向转运蛋白.CpSit1蛋白保守结构域为MFS_1结构域,且蛋白质二级结构以α螺旋、延伸链和无规则卷曲为主.根据TMHMM在线分析,发现CpSit1蛋白共有14个跨膜螺旋,为典型的MFS折叠.根据摇杆开关和门控运输理论[14-15],推测CpSit1蛋白与底物结合后,其N端或C端的7个跨膜螺旋围绕着底物结合位点一侧关闭,另一侧开放,以进行转运.
本研究通过qRT-PCR进行验证,结果显示,CpSit1基因在焦枯病菌侵染桉树24 h后即发生表达,达到23.99倍,明显高于48和72 h;48和72 h的上调倍数低于2倍,差异表达不显著.表明桉树焦枯病菌的CpSit1基因在病原菌侵染寄主的过程中通过调控转运铁载体-铁化合物的转运来完成铁元素的摄入,协助其在寄主中的定植;SIT转运蛋白作为铁载体-铁化合物的运输途径,与病原真菌的生长发育和致病性存在密切联系,这与前人的研究结果[16-20]一致.
[1] HAAS H. Molecular genetics of fungal siderophore biosynthesis and uptake: the role of siderophores in iron uptake and storage[J]. Appl Microbiol Biotechnol, 2003,62(4):316-330.
[2] HELM D V D, WINKLMANN G. Metal Ions in Fungi[M]. Tübingen:Marcel Dekker Inc, 1994:39-98.
[3] SAIER M H. Genome archeology leading to the characterization and classification of transport proteins[J]. Curr Opin Microbiol, 1999,2(5):555-561.
[4]SAIER M H. Tracing pathways of transport protein evolution.[J]. Molecular Microbiology, 2003,48(5):1 145-1 156.
[5] PARK Y S, KIM T H, CHANG H I, et al. Cellular iron utilization is regulated by putative siderophore transporter FgSit1 not by free iron transporter inFusariumgraminearum[J]. Biochem Biophys Res Commun, 2006,345(4):1 634-1 642.
[6] RAYMOND-BOUCHARD I, CARROLL C S, NESBITT J R, et al. Structural requirements for the activity of the MirB ferrisiderophore transporter ofAspergillusfumigatus[J]. Eukaryotic Cell, 2012,11(11):1 333-1 344.
[7] MOORE R E, KIM Y, PHILPOTT C C. The mechanism of ferrichrome transport through Arn1p and its metabolism inSaccharomycescerevisiae[J]. Proc Natl Acad Sci USA, 2003,100(10):5 664-5 669.
[8] BOOTH T H, JOBANOVIC T, OLD K M, et al. Climatic mapping to identify high-risk areas forCylindrocladiumquinqueseptatumleaf blight onEucalyptusin mainland South East Asia and around the world[J]. Environ Pollut, 2000,108(3):365-372.
[9] 陈全助,陈慧洁,郭文硕,等.桉树焦枯病菌(Calonectriapseudoreteaudii)生物学特性测定[J].福建林学院学报,2014,34(4):328-332.
[10] CHEN S F, LOMBARD L, ROUX J, et al. Novel species ofCalonectriaassociated withEucalyptusleaf blight in Southeast China[J]. Persoonia, 2011,26:1-12.
[11] LOMBARD L, CROUS P W, WINGFIELD B D, et al. Phylogeny and systematics of the genusCalonectria[J]. Stud Mycol, 2010,66:31-69.
[12] CHEN Q, GUO W, FENG L, et al. Transcriptome and proteome analysis ofEucalyptusinfected withCalonectriapseudoreteaudii[J]. J Proteomics, 2015,115:117-131.
[13] KUMAR S, STECHER G, TAMURA K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Mol Biol Evol, 2016,33(7):1 870-1 874.
[14] DEFELICE L J. Transporter structure and mechanism[J]. Trends in Neurosciences, 2004,27(6):352-359.
[15] SHIMAMURA T, WEYAND S, BECKSTEIN O, et al. Molecular basis of alternating access membrane transport by the sodium-hydantoin transporter Mhp1[J]. Science, 2010,328(5 977):470-473.
[16] 路媛媛.铁离子对玉米弯孢叶斑病菌生长发育及致病性的影响[C]//中国植物病理学会.中国植物病理学会2016年学术年会论文集.南京:中国植物病理学会,2016:1.
[17] OIDE S, MOEDER W, KRASNOFF S, et al. NPS6, encoding a nonribosomal peptide synthetase involved in siderophore-mediated iron metabolism, is a conserved virulence determinant of plant pathogenic ascomycetes[J]. Plant Cell, 2006,18(10):2 836-2 853.
[18] HOF C, EISFELD K, WELZEL K, et al. Ferricrocin synthesis inMagnaporthegriseaand its role in pathogenicity in rice[J]. Mol Plant Pathol, 2007,8(2):163-172.
[19] 王迪.玉米大斑病菌T-DNA插入突变体库的构建及StNPS6基因的功能研究[D].长春:吉林大学,2014.
[20] YIN Z, LIU H, LI Z, et al. Genome sequence ofValsacanker pathogens uncovers a potential adaptation of colonization of woody bark[J]. New Phytol, 2015,208(4):1 202-1 216.
(责任编辑:叶济蓉)
Identification and expression analysis ofCpSit1 gene inCalonectriapseudoreteaudii
LIU Hongyi1, YE Xiaozhen2, CHEN Huijie1, LI Huimin1, DING Yi1, YANG Zehui1, FENG Lizhen1
(1.College of Forestry; 2.College of Jinshan, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China)
Cpsit1 was identified by Blast and TCDB database, and then the physicochemical property, transmembrane helix, secondary structure and subcellular localization ofCpSit1 were analyzed and predicted by SMART database, ProtParam, TMHMM, PHD and ProComp 9.0 online service. Finally expression ofCpSit1 gene was detected by qRT-PCR within 24, 48 and 72 hCalonectriapseudoreteaudiiinfection of Eucalyptus. Results showed thatCpsit1 was identified as proton-coupled symporter for siderophore, with CpSit1 gene being 1 780 bp and encoding 592 amino acids. And the conserved domain ofCpSit1 is MFS_1. α-helix accounted for 45.10% of the secondary structure, with extension chain and random coil reaching 26.01% and 28.89%, respectively. QRT-PCR results revealed that expressions of CpSit1 gene were all upregulated within 24, 48 and 72 hC.pseudoreteaudiiinfection, and the expression level in 24 h was significantly higher than the other 2 stages. It proved thatCpSit1 gene could mediate the iron metabolism pathway ofC.pseudoreteaudiiby regulating siderophore and assistC.pseudoreteaudiiin the process of colonization.
Eucalyptus;Calonectriapseudoreteaudii; siderophore-iron transporter; bioinformatics; gene expression
2017-01-18
2017-05-17
福建省财政厅资助项目(K81150002、K81139238、K8112004A).
刘宏毅(1990-),男,硕士研究生.研究方向:森林病理学.Email:fjliuhongyi@126.com.通讯作者叶小真(1983-),女,讲师,博士.研究方向:森林病理学.Email:lisayxz@163.com.
S763.15
A
1671-5470(2017)04-0418-05
10.13323/j.cnki.j.fafu(nat.sci.).2017.04.010
猜你喜欢
新疆农业科学(2022年6期)2022-07-13 08:02:52
南方农业(2021年18期)2021-12-25 14:01:24
环境保护与循环经济(2021年7期)2021-11-02 08:10:42
林业科技情报(2020年3期)2020-03-02 13:21:36
生物工程学报(2019年11期)2019-12-09 13:09:12
西南农业学报(2019年6期)2019-07-18 05:32:16
心肺血管病杂志(2019年1期)2019-04-22 01:12:00
教学考试(高考生物)(2017年4期)2017-12-13 09:02:41
林业科学研究(2017年4期)2017-08-07 10:18:08
华东理工大学学报(自然科学版)(2014年2期)2014-02-27 13:48:43
